
Abstract
Top-down effects of predators can have important consequences for ecosystems. Insectivorous birds frequently have strong predation effects on herbivores and other arthropods, as well as indirect effects on herbivores’ host plants. Diet studies have shown that birds in temperate ecosystems consume arthropods in winter as well as in summer, but experimental studies of bird predation effects have not attempted to quantitatively separate winter predation impacts from those in summer. To understand if winter foraging by insectivorous birds has consequences for arthropods or plants, we performed a meta-analysis of published bird exclusion studies in temperate forest and shrubland habitats. We categorized 85 studies from 41 publications by whether birds were excluded year-round or only in summer, and analyzed arthropod and plant response variables. We also performed a manipulative field experiment in which we used a factorial design to exclude birds from Quercus velutina Lam. saplings in winter and summer, and censused arthropods and herbivore damage in the following growing season. In the meta-analysis, birds had stronger negative effects on herbivores in studies that included winter exclusion, and this effect was not due to study duration. However, this greater predation effect did not translate to a greater impact on plant damage or growth. In the field experiment, winter exclusion did not influence herbivore abundance or their impacts on plants. We have shown that winter feeding by temperate insectivorous birds can have important consequences for insect herbivore populations, but the strength of these effects may vary considerably among ecosystems. A full understanding of the ecological roles of insectivorous birds will require explicit consideration of their foraging in the non-growing season, and we make recommendations for how future studies can address this.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Predators frequently have important influences on ecosystems through direct effects on prey and indirect effects on nonadjacent lower trophic levels through trophic cascades (Paine 1980; Schmitz et al. 2000; Shurin et al. 2002). Understanding the role of predators has become particularly important given that organisms at higher trophic levels are frequently extirpated more rapidly from degraded habitats than those at lower trophic levels (Duffy 2003). A large number of studies have focused on insectivorous birds as tractable subjects for experimental manipulation to quantify such top-down effects. Insectivorous birds have the potential to strongly influence terrestrial communities through feeding on insects and other arthropods, resulting in direct effects on populations of arthropods and indirect effects on plants (Van Bael et al. 2008; Mooney et al. 2010; Mäntylä et al. 2011). In temperate regions, research on these impacts of birds has generally focused on summer growing seasons when birds are breeding and insect herbivores and deciduous foliage are most prevalent. However, bird foraging on arthropods in winter may have important consequences for herbivore and arthropod predator populations in the following spring and summer.
Winter bird communities are often composed of a resident subset of the breeding bird community in addition to non-breeding species that migrate north in spring to nest. The effects of bird predation in a system may differ with seasonal changes in bird community composition: in tropical agroforests, predation on arthropods increased in communities with higher migratory bird richness (Van Bael et al. 2008). The density of wintering birds varies considerably among temperate sites and habitats. In the mixed hardwood forests of Hubbard Brook (NH, USA), winter bird density was estimated at 1.5 birds/ha, less than 10 % of the summer abundance and summer bird biomass, although these winter estimates excluded species that did not breed at the site (Holmes and Sturges 1975). However, densities in Pennsylvania forests were several times higher than this (Graber and Graber 1979). In pine forests in Colorado, over 80 % of summer bird foraging activity was by year-round resident species (Mooney 2006, 2007). Wintering insectivorous birds can consume large numbers of terrestrial arthropods throughout the season. Betts (1955) found that tits (Paridae) in spruce forests commonly consumed Lepidoptera, Hemiptera, Coleoptera, and spiders, and the decline in arboreal arthropods throughout winter was attributed to this predation effect (Gibb 1960). Other observational studies in forests also suggest spiders, eggs, pupa, and insects including larvae and adults in diapause are important winter resources for insectivorous songbirds (Williams and Batzli 1979; Jansson and Bromssen 1981; Laurent 1986).
More recently, the effects of bird predation have been quantified using bird exclusion experiments. Although the majority of such experiments have been performed in temperate systems (e.g., >66 % of studies in Mooney et al. 2010 and Mäntylä et al. 2011) that exhibit strong seasonality, most studies focus exclusively on bird predation effects during the growing season. Many studies are limited to a single growing season (Holmes et al. 1979; Sipura 1999; Murakami and Nakano 2000; Mooney 2006; Barber and Marquis 2009, 2011a) or exclude birds year-round but do not distinguish between the effects of bird predation in different seasons (Marquis and Whelan 1994; Forkner and Hunter 2000; Strong et al. 2000; Lichtenberg and Lichtenberg 2002; Barber and Marquis 2011b). The few exclusion studies that examine winter bird predation have generally focused on predation of spiders (Askenmo et al. 1977), although Gunnarsson (1996) also documented bird effects on other arthropods in spring following winter exclusion, and Solomon et al. (1976) used cages to demonstrate that birds were a significant source of mortality for overwintering pupae of codling moth (Cydia pomonella). These studies all found significant effects of winter bird exclusion on prey.
Whether or not winter bird foraging suppresses arthropods may have consequences for the summer growing season. Feeding on herbivore pupa, eggs, or diapausing larva may reduce herbivore abundance and thus herbivory in the following summer through a cross-season trophic cascade. Overwintering insects may represent a population bottleneck, because these individuals can be the source of many more individuals in the following spring and summer. Because of this, an individual winter predation event could have a large impact on summer abundance. To understand the direct and indirect impacts of winter bird foraging, and to compare these effects to those of summer birds, two approaches were used. First, a meta-analysis was conducted using a database of insectivore exclusion studies (Mooney et al. 2010) to determine if results of studies that exclude birds in the non-growing season differ from those limited to the growing season. Second, an experiment was carried out in which bird access to Quercus velutina Lam. (black oak) was factorially manipulated in the non-growing and growing seasons to determine the abundance of arthropods and the resulting herbivore damage to plants.
Materials and methods
Meta-analysis
Studies were selected from a database of vertebrate exclusion studies that was previously analyzed by Van Bael et al. (2008) and Mooney et al. (2010). This database includes published and unpublished experiments that manipulated vertebrate insectivores (birds, bats, and/or lizards) using exclosures and measured responses of arthropods, plants, or both. For details of study selection or exclusion, see Mooney et al. (2010). We also included bird exclusion studies that have been published since the Mooney et al. (2010) database was completed. From these, a subset of studies were selected that (1) were conducted at latitudes greater than 30° (N or S) with seasonal growing and non-growing seasons, (2) focused on bird exclusion, and (3) gave details about the dates that the exclosures were in place.
Given that recent meta-analyses have shown clear direct effects of bird predation on arthropods and indirect effects on plants (Van Bael et al. 2008; Mooney et al. 2010; Mäntylä et al. 2011), our goal was not to repeat these analyses, but rather to determine the contribution of wintering birds in temperate systems to these direct and indirect effects. Each study was categorized as excluding birds in both the growing (“summer”) and non-growing (“winter”) seasons or in summer only. All studies measured arthropod and plant responses in the growing season (from spring to fall). Unweighted log response ratios (Hedges et al. 1999) were calculated for four types of responses: herbivores, arthropod predators, plant damage, and plant growth. This is the natural log of the ratio of the response when birds are present to the response when birds are excluded. A negative LRR indicates a reduction in the response variable by bird predation; for example, a negative herbivore LRR means fewer herbivores in control than exclusion treatments. Herbivore and arthropod predator responses included both abundance and biomass measurements. When studies reported longitudinal data, the last reported datapoint was used; this increases the likelihood that reported effects reflect systems at equilibrium, and reduces the chance of reporting phases of transient dynamics (Elser et al. 2007). Additionally, using final datapoints ensures that reported values from studies that excluded birds in both winter and summer will reflect potential effects of both winter and summer predation. General linear models were used to test the effect of the exclusion period on each response type. Because the length of the growing season generally decreases at higher latitudes, it was expected that any effects of winter versus summer exclusion might vary with latitude, so latitude was included as a covariate in all models. Studies limited to summer may be significantly shorter than year-round exclusion, so study duration (number of months from first setting up exclosures until final data collection, regardless of whether exclusion was continuous or not) was also included. Studies were also categorized by habitat (forest, shrubland, or grassland), but because no studies in grasslands have excluded birds in winter, results from grassland habitat would not be informative, so we excluded these studies and limited the analysis to experiments performed in forest or shrubland habitats. The difference in deviance between the full model and the model with each variable removed independently was compared using the drop1() command in R 2.13.0 (R Development Core Team 2010); this difference approximates a chi-square distribution (Zuur et al. 2009).
Field experiment
This experiment was performed at Tyson Research Center (St. Louis County, MO, USA; 38° 31′ 1″N, 90° 32′ 60″W) along a forest edge on a southeast-facing hillside approximately 0.6 km in length. The surrounding habitat is predominantly mixed oak-hickory forest where Q. velutina is a fairly common canopy tree and an uncommon understory tree. Seedlings and saplings of Q. velutina are most frequently found in gaps or along edges such as the study site. Common wintering birds at this site include Carolina chickadee (Poecile carolinensis), tufted titmouse (Baeolophus bicolor), eastern bluebird (Sialia sialis), northern cardinal (Cardinalis cardinalis), and dark-eyed junco (Junco hyemalis). Other than juncos, these species are also common breeding birds in addition to migrant passerines.
In fall 2007, 52 Q. velutina saplings were identified and separated into nine spatial blocks based on proximity. Saplings ranged from 0.8 to 2 m in height. In October 2007, PVC frames were constructed around each tree. Trees separated by less than 1 m were contained within the same frame so that there were four frames per block (36 total) (Fig. 1). Frames were randomized by bird exclusion treatment so that each block contained one frame per treatment combination: birds excluded in winter, summer, both seasons, or no exclusion (control). Birds were excluded by stretching monofilament netting over frames during the appropriate seasons. This netting (2.5 cm openings) has been used in numerous other bird exclusion experiments and allows arthropods, including large lepidopterans, to pass through while excluding insectivorous birds (Marquis and Whelan 1994; Barber and Marquis 2011a, b). Netting was in place by early November 2007 for winter exclusion, and was switched for summer exclusion in April 2008 just prior to budbreak. Leaf litter that fell on exclosures was cleared during fall and winter, although litter that fell from trees within exclosures remained below the trees, so leaf litter within and outside exclosures was not visibly different.

One block of bird exclosure frames in winter
Arthropods were censused on all trees in mid-May, early July, and early September 2008. These times represent peaks in insect arthropod abundance on oaks in Missouri (Marquis and Whelan 1994), and are characterized by distinct herbivore species assemblages (Marquis and Le Corff 1997; Forkner et al. 2004). Censuses followed Forkner et al. (2004): the upper and lower surface of every leaf was inspected, and all arthropods were recorded. The number of leaves examined was also counted during each census to standardize arthropod abundance for each frame. Trees averaged 202.1 leaves (1SE = 18.1, range 41–414 leaves), and there were no significant differences in leaf number among treatments whether analyzed without (treatment, F 3,31 = 0.676, P = 0.573) or with (treatment, F 3,23 = 0.912, P = 0.451; block, F 8,23 = 2.351, P = 0.052) block included.
In the second week of October, samples of ten leaves were collected from each tree to estimate percent area consumed by herbivores. Leaves were collected systematically, so that if a tree had 100 leaves, every tenth leaf was collected by counting from the base of the tree toward branch tips. This ensures that sampled leaves are distributed throughout the plant. Herbivory was estimated visually by comparing to reference leaves of a known damage level.
We analyzed abundances of insect herbivores and arthropod predators [mostly ants, spiders, and harvestman (Opiliones)] separately using generalized linear mixed models (GLMMs) with a Poisson distribution and log link function. We fitted abundance models with the function glmmPQL() in the MASS package (Venables and Ripley 2002) of R, which calls lme() in the nlme package (Pinheiro et al. 2012) and accounts for overdispersion, treating block as a random variable and including number of leaves as a covariate. Each census was analyzed independently for two reasons. Although repeated censuses are usually treated as repeated measures of the same replicates, small variations in the number of leaves examined on each tree across the three censuses make it difficult to use a single leaf number as a covariate for all three censuses. Analyzing each census independently allows us to include a more precise covariate of sampling. Further, repeated censuses in a similar, adjacent Q. alba (white oak) system have been treated this way because few, if any, individual insects occur in more than one of the three peaks in insect abundance (Barber and Marquis 2011b). Mean leaf damage was analyzed using a GLMM with a Gaussian distribution and identity link, treating block as a random factor. For frames with multiple trees, damage was averaged among trees. In all models, we examined residuals to verify normality and homoscedasticity. We removed an influential outlier in the May and September herbivore analyses (Cook’s distance calculated from a model in which block was treated as fixed, May outlier = 17.9, September outlier = 7.9).
Results
Meta-analysis
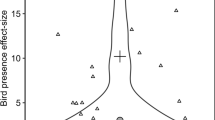
A total of 85 studies from 41 publications fit the criteria for inclusion in this analysis (see the Electronic supplementary material, ESM). Of these, 36 excluded birds in both winter and summer, and 49 were limited to summer. Studies with winter exclusion were longer than summer-only studies (mean months ± 1SE, winter 15.5 ± 1.8, summer 6.1 ± 1.3, t = 4.33, P < 0.001). There was a trend for studies with winter exclusion to be performed at lower latitudes, but the difference was minor (mean latitude ± 1SE, winter 42.4 ± 1.6, summer 45.9 ± 1.1, t = 1.83, P = 0.072). Birds had a significantly stronger effect on herbivores in studies that excluded birds during the winter (Table 1). Herbivore LRRs were on average 0.64 less in studies with winter exclusion, corresponding to a 28 % greater herbivore reduction than in studies that only excluded birds in the summer (Fig. 2). There was also a trend toward greater plant growth increases when birds were excluded in winter, but the effect was marginally significant. Bird effects on arthropod predators and plant damage did not differ between studies with and without winter exclusion. Herbivore, predator, and damage responses were significantly correlated with latitude such that bird effects were stronger (i.e., decreased arthropods and damage more) at higher latitudes, and effects on plant damage declined with study duration (Table 1). Bird predation effects on herbivores and arthropod predators did not differ between forest and shrubland habitats, although sample size in shrublands was limited (n = 9 studies).

Log response ratios (LRRs, ± 95% confidence intervals) of herbivores, arthropod predators, plant damage, and plant growth for studies excluding birds in both winter and summer (white circles) or only in summer (black circles). Numbers above confidence intervals are sample sizes
Field experiment
We recorded 453 individual herbivores representing 51 species or morphospecies (see the ESM) and 179 predators from five orders (Opiliones, Araneae, Coleoptera, Hemiptera, and Hymenoptera). These densities are comparable to those reported previously on understory Q. velutina (Le Corff and Marquis 1999; Forkner et al. 2004), although May herbivore densities in this study were lower [converted to herbivores/m2 leaf area based on mean leaf area of 96.7 cm2 (Le Corff and Marquis 1999), mean ± SE, May 1.26 ± 0.21, July 2.04 ± 0.37, September 3.54 ± 0.69]. However, this conversion likely underestimates the densities in this study. Although trees in this study were saplings, most were exposed to higher levels of sunlight than is typical in a forest understory, causing smaller leaves. One tree died prior to spring budbreak and could not be replaced.
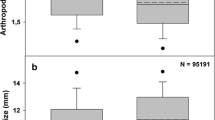
Treatments had relatively small effects on herbivore abundance. Winter exclusion alone never significantly increased herbivore abundance. Summer exclusion increased herbivores only in September, and the addition of winter exclusion eliminated this effect. Although herbivores were most abundant on control trees in July, this result was of marginal statistical significance. Bird exclusion did not significantly affect arthropod predator abundance, although in July control trees tended to have fewer predators than any of the other treatments (Table 2; Fig. 2). Leaf damage did not differ between the exclusion treatments (Table 3; Fig. 3).

Effects of bird exclusion on leaf damage to Quercus velutina. Values are estimated means based on a generalized linear mixed model, ±1SE. Con control, Sum summer bird exclusion, Win winter bird exclusion. There are no significant differences among treatment groups
Discussion
Despite strong evidence that birds can have important impacts on terrestrial plant communities through predation on arthropods (Van Bael et al. 2008; Mooney et al. 2010; Mäntylä et al. 2011), little is known about the consequences of such effects in winter. Using a meta-analysis of published studies, we have shown that winter-foraging birds do affect terrestrial communities, and that these effects can be detectable as strong reductions in herbivore densities the following growing season. However, these effects were not seen in the field experiment, where birds were factorially excluded in winter and summer. Below we discuss the results of both the meta-analysis and the field experiment, and make recommendations for how future studies might more accurately quantify the ecological effects of bird predation on their arthropod prey and on plants.
Meta-analysis
Studies that excluded birds in winter recorded a significantly stronger effect on herbivores than studies that only excluded in summer. While insectivorous birds have been shown to reduce herbivore abundance or biomass in many systems, and meta-analyses have shown this to be a widespread effect, this result indicates that these predation effects are influenced by the feeding activities of birds outside of the growing season. This result might be expected if studies that exclude birds in winter tend to be carried out over longer periods of time, which could result in increased effect sizes. Although studies that excluded in winter did tend to be longer, study duration was uncorrelated with bird effects on arthropods, in agreement with the results of Mooney et al. (2010).
The increased bird effects are probably due to direct winter feeding on herbivore eggs, pupae (Solomon et al. 1976), or diapausing larvae (Williams and Batzli 1979; Laurent 1986) or adults (Williams and Batzli 1979), which reduces the number of herbivores present in the following spring and summer. Another potential mechanism by which birds may affect herbivore abundances is by reducing densities of arthropod predators, thus releasing herbivores from additional predation [although Mooney et al. (2010) actually found stronger effects of vertebrate predators on herbivores when arthropod predators were more numerous]. Although birds reduced arthropod predators in this analysis (Fig. 2), the magnitude of this reduction did not differ between year-round and summer-only exclusion studies. Thus, it is unlikely that differential predation on arthropod predators explains the observed difference in bird effects on herbivores, and it is more likely that the difference is due to direct feeding on the herbivores themselves.
Increased predation of herbivores in winter, however, did not result in stronger effects on plant damage or growth. On average, birds increased plant growth more in studies with winter exclusion than in those without, but the effect was not significant, and plant damage effects did not follow this trend. Although the magnitude of indirect bird effects on plants is comparable to those in Mooney et al. (2010), the number of studies that quantified effects on plants is fairly small. Indirect predator effects on plants are often attenuated compared to direct effects on herbivores (Schmitz et al. 2000; Mooney et al. 2010), and with a limited number of studies measuring plant growth responses, it may be difficult to detect indirect effects of seasonal predation at a fine resolution.
This study was limited to forest and shrubland habitats. Although bird exclusion studies have been performed in several grassland ecosystems (e.g., Joern 1986, 1992; Belovsky and Slade 1993; Branson 2005), these experiments were always limited to summer exclusion. There may be practical challenges to maintaining exclosures in winter; in some multi-year grassland studies, exclosures were disassembled each fall and reassembled in the spring (e.g., Bock et al. 1992; Branson 2005). In grassland ecosystems that receive heavy snowfall, arthropods may not be accessible to birds in winter, but at sites with a milder non-growing season, potential impacts of bird predation may have been missed.
Field experiment
To better understand the independent effects of winter predation (rather than simply in addition to summer predation), we factorially excluded birds from saplings in winter and summer. Winter bird predation had little influence on insect herbivores or their effects on Q. velutina saplings in this study. Removing bird predation in the summer increased herbivore abundance only during the late-season census. Indeed, the only impact of winter bird exclusion was to remove this effect of summer exclusion: trees on which birds were excluded throughout the study did not differ in herbivore abundance from control or winter-exclusion-only trees (Fig. 4a). The absence of a bird predation effect in May and July is consistent with other recent exclusion experiments conducted on congeneric Q. alba at nearby locations, which found no direct effect of bird exclusion on summer herbivore abundance (Barber and Marquis 2009, 2011a, b). Although compensatory predation by arthropods is a possible explanation for an absence of vertebrate predation treatment effects, arthropod predator abundance was mostly unaffected by exclusion treatments, except in July, when they tended to be low only on control trees.

Effects of bird exclusion from Quercus velutina on a insect herbivore abundance and b arthropod predator abundance. Winter exclusion did not affect the abundance of either group, but summer exclusion alone increased herbivore abundance in September. Con control, Sum summer bird exclusion, Win winter bird exclusion
A lack of winter bird predation effects may be related to the life-history patterns of the herbivore community. Many of the Lepidoptera present as larvae in spring in this system oviposit late in the season or spend the winter as larvae in diapause. These represent likely winter food sources for insectivorous birds. However, spring herbivore abundances were low in all treatments in this experiment when compared to other studies of Q. velutina herbivores in the same region (Le Corff and Marquis 1999; Forkner et al. 2004). This may indicate low food availability for winter birds, which would likely forage in richer areas. Low abundance of spring herbivores may also in part explain leaf damage results. Spring herbivores are responsible for a significant proportion, and possibly the majority, of the total seasonal leaf herbivory on oaks (Barber and Marquis 2009). Despite the positive effects of summer bird exclusion on late-season herbivore abundance, there was no impact of birds on Q. velutina leaf damage. In other words, the increase in herbivore abundance did not translate into increased herbivory. Because late-season herbivores likely contribute less to total leaf damage, the lack of an indirect effect of birds on these oaks is not surprising.
Independent winter foraging effects were not detected, which contradicts expectations based on our meta-analysis results. There are characteristics of this system and site that may differentiate it from others in the meta-analysis. The plants in the experiment were saplings growing along a sun-exposed edge, so this is likely a high-productivity system, which trophic theory predicts should have strong top-down effects (Oksanen et al. 1981). In their comprehensive analysis, Mooney et al. (2010) showed that the ratio of arthropod predators to herbivores in experiments increased with the normalized difference vegetation index (NDVI), a measure of productivity. Compared to those experiments, our Q. velutina system had an average to lower-than-average predator:herbivore ratio [0.28, −1.07, and −1.32 in May, July, and September, respectively, calculated as ln(predators/herbivores)], despite being a high-productivity habitat. This may be due to high levels of defensive tannins in this sunny habitat; the condensed tannin concentration in Q. velutina is significantly higher in the sun-exposed canopy compared to the shaded understory (Forkner et al. 2004). Well-defended foliage may reduce the energy available to herbivores, reduce abundances of higher trophic levels, and thus dampen top-down effects during the growing season (Sipura 1999).
Synthesis and recommendations
We have demonstrated that winter foraging by insectivorous birds can affect insect populations, but the magnitude of this effect varies across ecosystems. To more accurately quantify and better understand bird predation effects, we suggest components that should be incorporated into future studies when feasible:
1. Year-round or factorial seasonal exclusion—results of exclusion experiments that are limited to the growing season may not accurately reflect the impacts of the local bird community. To estimate the independent effects of winter and summer predation, factorial designs similar to our field experiment will be necessary.
2. Longitudinal measurements—tracking herbivore and particularly arthropod predator populations prior to and during the growing season will allow ecologists to determine the role of temporal dynamics of taxonomic or trophic groups in these multitrophic systems.
3. Combining bird and arthropod predator exclusion—manipulating access by or the abundance of arthropod predators will clarify how bird and arthropod predation interactively influence herbivores and their impacts on plants (Mooney et al. 2010).
4. Considering life-history characteristics of arthropod prey—diverse arthropod communities often include species with diverse life histories, such as varied phenologies and voltinities. Different species also overwinter in different lifestages (Miliczky et al. 2008) and in different locations, which may influence susceptibility to predation. In particular, the importance of bird foraging in leaf litter versus on plants is unknown, and ecologists should consider how bird exclosures that exclude birds from the leaf litter below focal plants (as in this study) may influence results.
Despite these outstanding questions, we have clearly demonstrated that insectivorous birds in temperate systems can have important impacts on arthropod communities, even outside of the growing season. In our meta-analysis, the greater reduction of herbivores in studies with year-round exclusion indicates that winter predation by birds has effects in the subsequent spring and summer. However, these effects may be context dependent, as illustrated by our field study in which winter bird predation did not affect later herbivore abundance.
References
Askenmo C, von Brömssen A, Ekman J, Jansson C (1977) Impact of some wintering birds on spider abundance in spruce. Oikos 28:90–94
Barber NA, Marquis RJ (2009) Spatial variation in top-down direct and indirect effects on white oak (Quercus alba L.). Am Midl Nat 162:169–179
Barber NA, Marquis RJ (2011a) Light environment and the impacts of foliage quality on herbivorous insect attack and bird predation. Oecologia 166:401–409
Barber NA, Marquis RJ (2011b) Leaf quality, predators, and stochastic processes in the assembly of a diverse herbivore community. Ecology 92:699–708
Belovsky G, Slade JB (1993) The role of vertebrate and invertebrate predators in a grasshopper community. Oikos 68:193–201
Betts MM (1955) The food of titmice in oak woodland. J Anim Ecol 24:282–323
Bock CE, Bock JH, Grant MC (1992) Effects of bird predation on grasshopper densities in an Arizona grassland. Ecology 3:1706–1717
Branson DH (2005) Direct and indirect effects of avian predation on grasshopper communities in northern mixed-grass prairie. Environ Entomol 34:1114–1121
Duffy JE (2003) Biodiversity loss, trophic skew and ecosystem functioning. Ecol Lett 6:680–687
Elser JJ, Bracken MES, Cleland EE, Gruner DS, Harpole WS, Hillebrand H, Ngai JT, Seabloom EW, Shurin JB, Smith JE (2007) Global analysis of nitrogen and phosphorus limitation of primary production in freshwater, marine, and terrestrial ecosystems. Ecol Lett 10:1135–1142
Forkner RE, Hunter MD (2000) What goes up must come down? Nutrient addition and predation pressure on oak herbivores. Ecology 81:1588–1600
Forkner RE, Marquis RJ, Lill JT (2004) Feeny revisited: condensed tannins as anti-herbivore defences in leaf-chewing herbivore communities of Quercus. Ecol Entomol 29:174–187
Gibb JA (1960) Populations of tits and goldcrests and their food supply in pine plantations. Ibis 102:163–208
Graber JW, Graber RR (1979) Severe winter weather and bird populations in southern Illinois. Wilson Bull 91:88–103
Gunnarsson B (1996) Bird predation and vegetation structure affecting spruce-living arthropods in a temperate forest. J Anim Ecol 65:389–397
Hedges LV, Gurevitch J, Curtis PS (1999) The meta-analysis of response ratios in experimental ecology. Ecology 80:1150–1156
Holmes RT, Sturges FW (1975) Bird community dynamics and energetics in a northern hardwoods ecosystem. J Anim Ecol 44:175–200
Holmes RT, Schultz JC, Nothnagle P (1979) Bird predation on forest insects: an exclosure experiment. Science 206:462–463
Jansson C, Bromssen A (1981) Winter decline of spiders and insects in spruce Picea abies and its relation to predation by birds. Ecography 4:82–93
Joern A (1986) Experimental study of avian predation on coexisting grasshopper populations (Orthoptera: Acrididae) in a sandhills grassland. Oikos 46:243–249
Joern A (1992) Variable impact of avian predation on grasshopper assemblies in sandhills grassland. Oikos 64:458–463
Laurent JL (1986) Winter foraging behaviour and resource availability for a guild of insectivorous gleaning birds in a southern alpine larch forest. Ornis Scand 17:347–355
Le Corff J, Marquis RJ (1999) Difference between understory and canopy in herbivore community composition and leaf quality for two oak species in Missouri. Ecol Entomol 24:46–58
Lichtenberg JS, Lichtenberg DA (2002) Weak trophic interactions among birds, insects and white oak saplings (Quercus alba). Am Midl Nat 148:338–349
Mäntylä E, Klemola T, Laaksonen T (2011) Birds help plants: a meta-analysis of top-down trophic cascades caused by avian predators. Oecologia 165:143–151
Marquis RJ, Le Corff J (1997) Estimating pre-treatment variation in the oak leaf-chewing insect fauna of the Missouri Ozark Forest Ecosystem Project (MOFEP). In: Brookshire BL, Shifley S (eds) Proceedings of the Missouri Ozark Forest Ecosystem Project Symposium: an experimental approach to landscape research (Gen Tech Rep NC-193). USDA Forest Service, Washington, DC, pp 342–346
Marquis RJ, Whelan CJ (1994) Insectivorous birds increase growth of white oak through consumption of leaf-chewing insects. Ecology 75:2007–2014
Miliczky ER, Horton DR, Calkins CO (2008) Observations on phenology and overwintering of spiders associated with apple and pear orchards in south-central Washington. J Arachnol 36:565–573
Mooney KA (2006) The disruption of an ant–aphid mutualism increases the effects of birds on pine herbivores. Ecology 87:1805–1815
Mooney KA (2007) Tritrophic effects of birds and ants on a canopy food web, tree growth, and phytochemistry. Ecology 88:2005–2014
Mooney KA, Gruner DS, Barber NA, Van Bael SA, Philpott SM, Greenberg R (2010) Interactions among predators and the cascading effects of vertebrate insectivores on arthropod communities and plants. Proc Natl Acad Sci USA 107:7335–7340
Murakami M, Nakano S (2000) Species-specific bird functions in a forest-canopy food web. Philos Trans Roy Soc B 267:1597–1601
Oksanen L, Fretwell SD, Arruda J, Niemela P (1981) Exploitation ecosystems in gradients of primary productivity. Am Nat 118:240–261
Paine RT (1980) Food webs: linkage, interaction strength and community infrastructure. J Anim Ecol 49:667–685
Pinheiro J, Bates D, DebRoy S, Sarkar D, R Development Core Team (2012) nlme: linear and nonlinear mixed effects models (in R package version 3.1-103). R Foundation for Statistical Computing, Vienna
R Development Core Team (2010) R: a language and environment for statistical computing. R Foundation for Statistical Computing, Vienna
Schmitz OJ, Hamback PA, Beckerman AP (2000) Trophic cascades in terrestrial systems: a review of the effects of carnivore removals on plants. Am Nat 155:141–153
Shurin JB, Borer ET, Seabloom EW, Anderson K, Blanchette CA, Broitman B, Cooper SD, Halpern BS (2002) A cross-ecosystem comparison of the strength of trophic cascades. Ecol Lett 5:785–791
Sipura M (1999) Tritrophic interactions: willows, herbivorous insects and insectivorous birds. Oecologia 121:537–545
Solomon ME, Glen DM, Kendall DA, Milsom NF (1976) Predation of overwintering larvae of codling moth (Cydia pomonella (L.) by birds. J Appl Ecol 13:341–352
Strong AM, Sherry TW, Holmes RT (2000) Bird predation on herbivorous insects: indirect effects on sugar maple saplings. Oecologia 125:370–379
Van Bael SA, Philpott SM, Greenberg R, Bichier P, Barber NA, Mooney KA, Gruner DS (2008) Birds as predators in tropical agroforestry systems. Ecology 89:928–934
Venables WN, Ripley BD (2002) Modern applied statistics with S, 4th edn. Springer, New York
Williams JB, Batzli GO (1979) Winter diet of a bark-foraging guild of birds. Wilson Bull 91:126–131
Zuur AF, Ieno EN, Walker NJ, Saveliev AA, Smith GM (2009) Mixed effects models and extensions in ecology with R. Springer, Berlin
Acknowledgments
We thank A. Loayza, E. Justus, and the staff at Tyson Research Center for invaluable help. K. Mooney and three anonymous reviewers provided helpful comments that greatly improved the manuscript. This work was supported by University of Missouri-TWA scholarships, the St. Louis Audubon Society, USEPA-STAR Fellowship #FP91648501, and Northern Illinois University.
Author information
Authors and Affiliations
Corresponding author
Additional information
Communicated by Sven Bacher.
Electronic supplementary material
Below is the link to the electronic supplementary material.
Rights and permissions
About this article
Cite this article
Barber, N.A., Wouk, J. Winter predation by insectivorous birds and consequences for arthropods and plants in summer. Oecologia 170, 999–1007 (2012). https://doi.org/10.1007/s00442-012-2367-z
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00442-012-2367-z