
Abstract
Carbon and nitrogen stable isotope ratios are used to assess diet composition by determining bounds for the relative contributions of different prey to a predator’s diet. This approach is predicated on the assumption that the isotope ratios of predator tissues are similar to those of dominant food sources after accounting for trophic discrimination (ΔxX), and is formulated as linear mixing models based on mass balance equations. However, ΔxX is species- and tissue-specific and may be affected by factors such as diet quality and quantity. From the different methods proposed to solve mass balance equations, some assume ΔxX to be exact values whilst others (based on Bayesian statistics) incorporate variability and inherent uncertainty. Using field data from omnivorous reef fishes, our study illustrates how uncertainty may be taken into account in non-Bayesian models. We also illustrate how dietary interpretation is a function of both absolute ΔxX and its associated uncertainty in both Bayesian and non-Bayesian isotope mixing models. Finally, collated literature illustrate that uncertainty surrounding ΔxX is often too restricted. Together, these data suggest the high sensitivity of mixing models to variation in trophic discrimination is a consequence of inappropriately constrained uncertainty against highly variable ΔxX. This study thus provides guidance on the interpretation of existing published mixing model results and in robust analysis of new resource mixing scenarios.
Similar content being viewed by others
Avoid common mistakes on your manuscript.
Introduction
Stable isotope analysis (SIA) is a powerful tool, widely used in food web and trophodynamics research (Vander Zanden et al. 1999; Pinnegar and Polunin 2000). SIA applications are based on constrained and element-specific trophic-induced isotopic shifts between consumers and their resources (Δ15N or Δ13C generalized as ΔxX). Δ13C is usually small, such that carbon data reflect basal resources, and will inform on primary production sources sustaining species (Melville and Connolly 2003; Benstead et al. 2006) or principal prey items and their relative mixes (Phillips 2001; Moore and Semmens 2008). In contrast, Δ15N is large and relates to an individual’s trophic position (Post 2002). Variance in isotope values may also be used to infer information on niche breadth (Bearhop et al. 2004; but see Flaherty and Ben-David 2010).
The use of SIA in assessment of diet composition has grown exponentially, providing time integrative and intra-population resolution trophic data, which has proved particularly powerful in combination with the fine temporal and taxonomic resolution data derived from direct dietary observations (Schindler and Lubetkin 2004). Such resource mixing approaches are predicated on the assumption that isotope ratios of predator tissues are similar to that of dominant food sources after accounting for ΔxX (DeNiro and Epstein 1976), and that dominant food sources can be separated isotopically. A predator’s tissues are thus an isotopic mixture of its dietary sources; SIA can then indicate the proportional contributions of these sources, but only when the isotopic signatures of each source differ.
Initial mixing models were based on balancing equations (Fry 2006) and solved through a series of linear equations. When n isotope systems are used, the unique proportional contribution of n + 1 isotopically distinct dietary sources can be resolved for a given ΔxX in each isotope system (Phillips 2001; Martinez del Río and Wolf 2005) (Eqs. 1, 2).
and
For example, the above linear mixing model has an exact solution with two isotopes and three sources. However, the number of potential sources often greatly exceeds the number of isotopes systems, preventing identification of unique solutions of source proportions.
The need to incorporate multi-source mixing has led to development of new modeling approaches based not on unique solutions but on identification of potential ranges. The first software developed to resolve these Multisource Mass-balance Mixing Models (4M) (IsoSource; Phillips and Gregg 2003) was based on iterative techniques and provided constrained boundaries of source utilization and ranges of feasible mixes in the nutrition of predators. IsoSource calculates all possible combinations of source proportions that sum to 100% in small increments (e.g., 1%). Then, the predicted isotope values of each mixture are computed using linear mixing models that preserve mass balance (Eqs. 1, 2). Isotope values of computed mixtures are then compared with the observed isotope values; the range of combinations that matches within a user-specified tolerance value (e.g., 0.1‰) is then described. Thus, IsoSource outputs are ranges of feasible contributions for each source, given as minima and maxima or 1–99% percentiles, and their usefulness and interpretation depend on how constrained these ranges are. Large ranges are usually uninformative, unless the minimum is relatively high, which identifies that the source is important. Small ranges represent relatively well-constrained estimates of the source contribution and give a useful solution.
Since Phillips and Gregg (2003), a number of other methods have been proposed (see Box 1 in “Literature review”, below). Such resource mixing models have many applications: for example, they have been used to describe plant water uptake dynamics (Asbjornsen et al. 2007) and rainfall infiltration processes in soils (Bao et al. 2009), trace the contribution of aged and contemporary organic matter to rivers (Caraco et al. 2010), estimate contributions of different sediment sources to water channels or estuaries (Collins et al. 2010; Gibbs 2008), and dietary proportional contributions (Moore and Semmens 2008; Parnell et al. 2010).
As with IsoSource, 4M models generally are most useful in further applications when they identify constrained potential mixes. However, the likelihood that this occurs is dependent on the geometry of the convex hull (also referred to as the prey polygon; the smallest convex perimeter that includes all points) formed by sources in isotope space, the position of the predator within that polygon after back-correcting for ΔxX (Phillips and Gregg 2003; Moore and Semmens 2008) (Fig. 1) and the number of sources. When the consumer is in the middle of the resource convex hull, the range of feasible solutions tends to be diffuse and few potential mixes may be ruled out (Phillips and Gregg 2003). As the number of sources increases, models also become increasingly underdetermined and feasible range estimates generally increase such that outputs becomes less informative (Boecklen et al. 2011).

Model input for stable isotope mixing models for two predatory fish, a Escrófalo and b Turco and various prey. Triangles represent δ13C and δ15N predator values: open down-triangle represents original values, open up-triangle represents values after Δ13C = 1 and Δ15N = 3.4 corrections, solid up-triangle represents values after Δ13C = 1.5 and Δ15N = 3.2 corrections. Shaded box represents δ13C and δ15N possible predator values after Δ13C and Δ15N corrections under high uncertainty (HU) scenario for non-Bayesian implementations. Inset white box represents δ13C and δ15N possible predator values after Δ13C and Δ15N corrections under low uncertainty (LU) scenario for non-Bayesian implementations. Curves represent Δ13C and Δ15N assuming LU (dashed lines) and HU (dotted lines) scenarios. Solid circles represent δ13C and δ15N prey values: SU sea urchin, Cr crab, Ch chiton, FA fishA, FB fishB, Sc scallop, Oc octopus, An anchovy, Pl polychaete
Three factors determine the placement of a consumer within the convex hull, one ecological and the other two methodological. In the former, feeding strategy influences where a consumer is likely to sit. Centrally-placed consumers are more common for generalist opportunist predators because they frequently lack dominant prey items and or utilize a wide range of resources. Methodologically, the placement of a consumer depends on the ΔxX and the precision with which the convex hull is defined. The use of inadequate ΔxX values will result in inaccurate diet proportions (Bond and Diamond 2011) and thus has been one stimulus for repeated calls for better understanding of ΔxX and the factors that affect it (Caut et al. 2008; Wolf et al. 2009; Martinez del Río et al. 2009), such as in elasmobranchs (Hussey et al. 2010; Logan and Lutcavage 2010). Similarly, the convex hull is given with definitive boundaries, but the isotopic signatures of the resources themselves are variable.
Although there is need for more experimental research identifying factors driving variation in ΔxX, the best that may be realistically achieved is a species- and tissue-specific mean ΔxX and constrained variance based on an understanding of, for example, the effects of ration size, diet quality or diet isotopic ratios on ΔxX (Caut et al. 2009, 2010; Auerswald et al. 2010; Perga and Grey 2010). Such variance in estimates of ΔxX will likely persist in analyses despite increased understanding, as they cannot typically be obtained for all wild populations, especially where characterization of the diet is a principal objective of analyses. Moreover, experimental determination of ΔxX is not possible for some species that are difficult to maintain in captivity, and uncertainty will surround extrapolating experimental results to wild populations.
It is therefore informative to explore how the different 4M perform in relation not just to ΔxX but to ΔxX uncertainty. Few 4M models addressed uncertainty formally, SIAR being one example. In others (e.g., IsoSource, Source&Step, and LP-Tracer) uncertainly must be addressed indirectly, outside of model analyses, for example via a sensitivity analysis. The main goal of this paper is to (1) show how ΔxX uncertainty is or could be incorporated in the different 4M packages, (2) assess the consequences of including variance in ΔxX as either a range or a mean with its standard deviation, and (3) evaluate how informative the different 4M outputs are. In doing so, the assumptions and tolerances of 4M techniques in source mixing are assessed to inform future appropriate use of such analyses.
Materials and methods
Data
Analyses were conducted on two published fish predator–prey datasets of Galván et al. (2009) containing both gut content and isotope data. Both species are predatory reef-fish that inhabit northern Patagonian shore reefs. Pinguipes brasilianus (hereafter ‘turco’ by its common name) feeds mainly on reef invertebrates, principally grazers and filter-feeders, but also small reef fish (Galván et al. 2009). Sebastes oculatus (hereafter ‘escrófalo’) is as an ambush predator that feeds opportunistically on mobile organisms that pass near its refuge (Barrientos et al. 2006; Galván et al. 2009); it consumes pelagic and benthic fish and invertebrates. Fishes are a ideal group to evaluate 4M performance being the taxa to which mixing models are most frequently applied (Boecklen et al. 2011).
Samples of predator dorsal muscle (n = 6 for each fish) and whole prey were collected and processed using standard protocols for C and N isotope data (see Galván et al. 2009 for details). Twelve potential common prey species were investigated, but because of small isotopic differences among several prey, these species were pooled, leading to seven general groups for each consumer. For turco, SU = sea urchin (Arbacia dufresnei); Cr = crab (Rochinia gracilipes and Leurocyclus tuberculosus); Ch = chiton (Chaetopleura isabelle); FA = fishA (Triathalassothia argentina and Dules auriga); FB = fishB (Ribeiroclinus eigenmanni and Helchogrammoides cunninghami); Sc = scallop (Aequipecten tehuelchus) and Pl = polychaete (Eunices argentinensis), were selected as possible food sources. Oc = octopus (Octopus tehuelchus), An = anchovy (Engraulis anchoita); FA, FB, Cr, Sc and Pl were selected for escrófalo (Fig. 1). Seven prey sources were used for both predators to avoid possible biases resulting from differing numbers of sources.
Model implementations and scenarios
IsoSource, Source&Step, LP-Tracer (thereafter named together as non-Bayesian implementations) and SIAR were performed under different Δ13C and Δ15N uncertainty scenarios [single values (SV), low uncertainty (LU) and high uncertainty (HU)] based principally on estimates of fish-specific ΔxX values (Sweeting et al. 2007a, b).
For SV, non-Bayesian applications were performed using ΔxX corrections without uncertainty. Two different estimates (a, b) of ΔxX were explored: SVa (Δ13C = 1.0‰ and Δ15N = 3.4‰) representing a global mean of all species tissues and habitats (e.g., Vander Zanden et al. 1999) and SVb (Δ13C = 1.5‰ and Δ15N = 3.2‰) representing fish-specific ΔxX values (Sweeting et al. 2007a, b).
For LU, ΔxX was constrained within the most probable range. In non-Bayesian 4M, Δ15N was varied between 3.0 and 3.4‰ (Sweeting et al. 2007a), whilst Δ13C varied from 1.0 to 2.0‰ (Sweeting et al. 2007b). Each 4M run used 0.2‰ increments of Δ13C and Δ15N values covering all possible combinations (Fig. 1). For the Bayesian approach, ΔxX LU was taken as one SD about the mean estimate from Sweeting et al. (2007a, b) for fish (Δ13C = 1.5 ± 0.4‰ and Δ15N = 3.2 ± 0.4‰).
The LU scenario was also performed for Bayesian models with (LUp) and without (LU) utilizing the capacity of Bayesian models to incorporate further prior information (Parnell et al. 2010), in this case, gut contents analysis (escrófalo: Cr = 0.55 ± 0.1, Oc = 0.05, FA = 0.1, FB = 0.1, Sc = 0.05, Po = 0.05, An = 0.1; turco: SU = 0.25 ± 0.05, FA = 0.05, FB = 0.05, Cr = 0.2, Sc = 0.15, Po = 0.2, and, Ch = 0.1; Barrientos et al. 2006; Galván et al. 2009), otherwise uncertainty in δ13C and δ15N of prey values (i.e., δ13C and δ15N prey mean ± SD) was the same. Dietary proportions naturally follow a Dirichlet distribution; and the distribution parameters (α) are the values that the user can input into the model. α parameters were calculated using prey proportions and the standard deviation of one source (Cr for escrófalo and SU for turco) by means of a specific SIAR function (Parnell and Jackson 2010).
The HU scenario represents a situation with unknown species-specific ΔxX and includes nearly the whole range of Δ13C and Δ15N published for fish tissues (reviews in Sweeting et al. 2007a, b). For non-Bayesian applications, Δ13C varied from 0.0 to 2.6‰ and Δ15N from 1.0 to 4.6‰. As in LU scenarios, models were run with 0.2‰ increments. For SIAR, ΔxX values were means ± SD of muscle from controlled feeding studies in Sweeting et al. (2007a, b) (Δ15N = 3.2 ± 1.3‰ and Δ13C = 1.5 ± 1.2‰). This scenario was also performed adding, as in LU scenario, prior potential contributions based on gut contents (HUp).
Results are reported following the authors’ recommendation for each 4M implementation (i.e., IsoSource: 1–99% percentiles, LP-Tracer and Source&Step: minimum and maximum values, and SIAR: 95% credible interval).
Literature review
To evaluate how ΔxX and associated variation is currently addressed in describing diet mixing a literature review was conducted of 4M literature through the SCOPUS electronic database. The search was restricted to that literature where 4M was applied to dietary description of fishes. Searches covered January 2009 (approximately 1 year after first 4M Bayesian publication; Box 1) up to October 2011.
Results and discussion
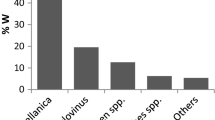
The literature review identified 54 publications that used one of the five 4M presented in Box 1 since 2009 to estimate the diet composition of over 100 different fish species (Online Resource 1). Bayesian models represent 40% of the literature in this period. While the proportion has increased yearly, Bayesian methods represented half of the mixing literature by the end of the review period, despite sound theoretical benefits for their use to incorporate uncertainty.
Among all 4M studies collated, none used a species- and tissue-specific ΔxX. Uncertainty in ΔxX was therefore a ubiquitous problem in the mixing models reviewed here. ΔxX proxies were either from similar species or derived from reviews. Only 10% (six) of studies suggest the utilization of a proxy from similar species. Thus, there was a heavy reliance of literature review generated means of ΔxX. Problematically, single ΔxX values varied substantially among reviews based on publication date and data and inclusion criteria, such that estimates of Δ13C ranged from 0 to 1.4‰ and Δ15N, from 2.2 to 3.6‰ (e.g., Post 2002; McCutchan et al. 2003; Caut et al. 2009).
None of the non-Bayesian studies have incorporated uncertainty in ΔxX to date. Incorporating ΔxX uncertainty more easily is one driver of recommendations for switching to Bayesian models. However, there was little consistency among Bayesian models reviewed here as to the magnitude of uncertainty applied, despite a similar samples type (fish white muscle), objective (dietary description), and restrictions (use of generic ΔxX). The combination of disparate choices of ΔxX and a commonly low uncertainty leads to variable dietary interpretation (Bond and Diamond 2011). In contrast, those studies that use high uncertainty are likely to produce comparable results regardless of specific choices of ΔxX due to high overlap. Eight studies using Bayesian models (40% of Bayesian studies reviewed) used ΔxX uncertainties comparable with our HU scenario.
So how does uncertainty impact trophic interpretation? The combination of both uncertainty in trophic shifts and the numerous potential prey items proved particularly problematic in determining constrained resource mixing in omnivorous fishes. Under the HU scenario, 42 and 50% of the possible predator positions were inside the prey polygon of escrófalo and turco, respectively (Fig. 1), while 100 and 66% were inside the prey polygon assuming the LU scenario and all were in the prey polygon under the SV scenario (Fig. 1). Thus, HU results summarize 112 and 134 outputs whilst LU results summarize 18 and 12 outputs for escrófalo and turco, respectively. Solutions that resulted in predator positions that lay outside the prey polygon after corrections were mostly due to Δ13C values, although which Δ13C and Δ15N combinations resulted in non-feasible values differed among species (Fig. 1). When Δ13C was >1.8‰, turco values were outside the polygon, irrespective of the Δ15N value. For escrófalo Δ13C values, <1.0‰ fell outside the polygon. Assuming that all important prey items were either sampled or at least had isotopic signatures that fell within the prey polygon proposed, the latter result showed that potential Δ13C variation among different fish species could be bigger than the δ13C difference between extreme prey (e.g., anchovy = pelagic prey vs. criptic-fish = benthic prey) for escrófalo.
Non-Bayesian methods had similar outputs (Table 1), but IsoSource’s outputs were the most constrained, providing narrower ranges than other models (Table 1). However, IsoSource’s outputs represent 1–99% quantiles of the feasible results, instead of the whole feasible range, and, when IsoSource’s maximum and minimum feasible contributions were considered, results were nearly identical to Source&Step and LP_Tracer results. The IsoSource feature to inform 1–99 percentiles has the advantage of truncating distributions that have very small numbers of observations in long tails and is therefore more robust to these possible outliers (Benstead et al. 2006).
The results obtained using non-Bayesian methods under a HU scenario were non-informative due to the broad range of possible contributions calculated for each prey (Table 1), ranging from 0 to over 90% for three prey (fishA, fishB, and polychaetes) and two (fishA and fishB) prey groups in escrófalo and turco, respectively. Among the possible Δ13C and Δ15N combinations, results varied from well-constrained outputs (e.g., IsoSource output, escrófalo, Δ13C = 1.2‰ and Δ15N = 4.4‰, fishB 46–56%) to diffuse outputs (e.g., IsoSource output, escrófalo, Δ13C = 1.6‰ and Δ15N = 3.0‰, fishB 4–65%) (Fig. 2). This is mainly because under HU assumptions the predator position moved from one side to the other of the prey polygon and as a result of this feasible contribution varied from ‘main prey’ to ‘minor prey’ (Fig. 3).

Histograms show the spread of feasible dietary contributions under high uncertainty (HU) scenario running non-Bayesian approaches for (a–g) prey of Escrófalo and (h–n) prey of Turco. Solid line and dotted line represent the spread of feasible solutions using Δ13C = 1.0‰ and Δ15N = 3.4‰ (SVa scenario) and Δ13C = 1.5‰ and Δ15N = 3.2‰ (SVb scenario) discrimination factors, respectively

Sensitivity analyses of IsoSource outputs under high uncertainty (HU) scenario. 3D plots show the 99% quantile of feasible dietary contribution for each prey in turco’s diet: a sea urchin, b crab, c chiton, d fishA, e fish B, f scallop, and g polychaete
Non-Bayesian methods under a LU scenario produced smaller overall ranges than HU (Table 1), but they were still poorly constrained for both species. Although some escrófalo prey had narrow ranges, for example, crab, scallop and anchovy with contributions ≤30% (Table 1, IsoSource output), all output mixes included zero as a possible contribution, and at least one other prey group with high variable contribution (e.g., escrófalo’s IsoSource output, fishB = 0–94%). These results occurred despite constraining ΔxX to the most probable range.
Under the SVa scenario, escrófalo had two mandatory prey (fishB and crabs) that were present in all feasible solutions, while other prey could be described as occasional or minor (Table 1). However, under the SVb scenario, only fishB was a mandatory prey, and other prey like polychaete or octopus could be more important than crabs (Table 1). This comparison shows how dramatically output interpretation could change within the most probable range of ΔxX values (see Dubois et al. 2007 for a similar comparison). In contrast to escrófalo, turco outputs were most constrained under SVa ΔxX assumptions (Table 1; Fig. 2). These results suggest that not only the dietary mixes for a given species but also comparative analyses among co-occurring predators face problematic interpretation as a consequence of ΔxX uncertainty. For example, the relative degree of omnivory differs for each fish under different ΔxX assumptions and it is unlikely that the same ΔxX is applicable to both species.
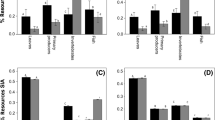
The Bayesian approach provided more constrained solutions under all the scenarios (Table 2; Fig. 4). However, it is not possible to directly compare non-Bayesian and Bayesian results, in that Bayesian models had the advantage of producing source contribution estimates with explicit probability distributions (see Box 1: SIAR and MixSIR). The results showed that LU and HU assumptions had very similar outputs when models with or without prior information were compared (Table 2). However, the comparison of models with the same uncertainty, but incorporated prior information, showed that the input of priors resulted in narrower credible intervals and fewer prey with 0‰ contributions as a feasible result.

Posterior estimates of proportional dietary contributions of prey sources to turco obtained running the software SIAR with both uninformative and informative priors under low uncertainty (LU and LUp, respectively) and uninformative priors under high (HU) scenarios
Under HU scenarios, Bayesian outputs showed some prey had narrower credible intervals compared to LU (e.g., polychaete as turco prey and fishB as escrófalo prey; Table 2; Fig. 4). These unintuitive results for the main prey item are similar to those of Moore and Semmens (2008), who also compared high and low uncertainty scenarios (see Moore and Semmens 2008; Table 1, Eggs and Fish). The unintuitive results are attributable to standard errors of the Δ13C and Δ15N corrections being large compared to the spread of the consumers (Fig. 1). This pattern was also evident for non-Bayesian methods under the HU scenario, which resulted in ~50% of solutions outside the prey polygon. There was then less ‘space’ for the dietary contributions to move around, and thus, uncertainty reduced, when the ΔxX standard error increases (A. Parnell, SIAR author, personal communication). However, the input of prior information mitigated this mathematical artifact and under HUp assumptions the estimated credible intervals were similar to under LUp scenario (Table 2).
Ecological interpretation of non-Bayesian 4M runs dramatically changed even with small shifts in the predator position inside the prey polygon. These shifts induced changes in the relative importance of diet items, the number of food sources eaten (omnivory), and, in the case of among species comparisons, interpretations of resource partitioning. Given the lack of specific ΔxX values, it seems necessary to conduct a sensitivity analyses among most probable ΔxX ranges. However, results here showed that the wide range of feasible ΔxX values usually results in poorly constrained and uninformative results. Even the comparison between two well-supported estimates of ΔxX correction (i.e., SVa vs. SVb) resulted in trophodynamics of each species (see Bond and Diamond 2011 for a similar result on Bayesian models).
Bayesian models incorporate ΔxX uncertainty in a useful way, and under low uncertainty scenarios gave constrained estimates of diet proportions. In addition, their outputs have the advantage of producing estimated probability distributions of source contributions to a mixture instead of only feasible solutions. On the other hand, mean values describing the average of all the source contribution estimates should be treated with utmost caution and never equated with unique solutions to the mixing problem; because the diet of predators is influenced by factors such as prey availability and palatability or competitive interactions, and none of the available implementations incorporate these factors; therefore, there is no reason to expect that its distribution means will accurately index the unique solutions that scientists seek (Benstead et al. 2006).
The unintuitive decrease in the 95% credible interval of prey contributions when ΔxX uncertainty increases in Bayesian models’ output emphasizes the need to use specific ΔxX values for isotopic values of species, tissues, and diets (Caut et al. 2008), and the importance of the call for more experiments (Wolf et al. 2009; Martinez del Río et al. 2009). In this regard, it is important to choose for dietary analyses the tissue with smallest ΔxX uncertainty; which in fish seems to be muscle when is compared with liver or heart (Sweeting et al. 2007a, b).
Pre-existing results of dietary analyses based on 4M non-Bayesian approaches must be used with careful consideration of ΔxX and compared to currently accepted mean values and sources of variation. Given the position of the predator inside the prey polygon, one can explore qualitatively how alterations in ΔxX are likely influence model outputs and feasible prey ranges. This analysis could be done following the discussion here and the rules of Phillips and Gregg (2003) for the graphical examination and interpretation of mixing diagrams.
Conclusions
Literature review illustrates a heavy reliance of mixing models on review-derived estimates of trophic shifts. These estimates have substantial uncertainty. A researcher’s choice of ΔxX can lead to fundamentally different trophic interpretations, particularly if uncertainty is inappropriately constrained, and this problem occurs in all 4M models, Bayesian or otherwise. Where uncertainty is high, differences in absolute ΔxX are less relevant, as widely overlapping ΔxX are assessed. We therefore recommend the use of species- and tissue-specific discrimination values with constrained uncertainty. In the absence of species- and tissue-specific values, the most parsimonious choice is to use generic discriminations values with high uncertainty and to input prior information from literature and field observations. In the absence of such prior information, under parameterization will generally result in poorly constrained model outputs.
References
Asbjornsen H, Mora G, Helmers MJ (2007) Variation in water uptake dynamics among contrasting agricultural and native plant communities in the Midwestern U.S. Agric Ecosyst Environ 121:343–356
Auerswald K, Wittmer MHOM, Zazzo A, Schäufele R, Schnyder H (2010) Biases in the analysis of stable isotope discrimination in food webs. J Appl Ecol 47:936–941
Bao W, Wang T, Hu H, Qu S (2009) Isotopic variations of soil water in rainfall infiltration experiment. Zhongshan Daxue Xuebao/Acta Scientiarum Natralium Universitatis Sunyatseni 48:132–137
Barrientos C, Gonzalez M, Moreno C (2006) Geographical differences in the feeding patterns of red rockfish (Sebastes capensis) along South American coasts. Fish Bull 104:489–497
Bearhop S, Adams C, Waldron S, Fuller R, Macleod H (2004) Determining trophic niche width: a novel approach using stable isotope analysis. J Anim Ecol 73:1007–1012
Benstead JP, March JG, Fry B, Ewel KC, Pringle CM (2006) Testing IsoSource: stable isotope analysis of a tropical fishery with diverse organic matter sources. Ecology 87:326–333
Boecklen W, Yarnes C, Cook B, James A (2011) On the use of stable isotopes in trophic ecology. Annu Rev Ecol Evol Syst 42:411–440
Bond A, Diamond A (2011) Recent Bayesian stable-isotope mixing models are highly sensitive to variation in discrimination factors. Ecol Appl 21:1017–1023
Bugalho MN, Barcia P, Caldeira MC, Cerdeira JO (2008) Stable isotopes as ecological tracers: an efficient method for assessing the contribution of multiple sources to mixtures. Biogeosciences 5:1351–1359
Caraco N, Bauer JE, Cole JJ, Petsch S, Raymond P (2010) Millennial-aged organic carbon subsidies to a modern river food web. Ecology 91:2385–2393
Caut S, Angulo E, Courchamp F (2008) Caution on isotopic model use for analyses of consumer diet. Can J Zool 86:438–445
Caut S, Angulo E, Courchamp F (2009) Variation in discrimination factors (Δ15N and Δ13C): the effect of diet isotopic values and applications for diet reconstruction. J Appl Ecol 46:443–453
Caut S, Angulo E, Courchamp F, Figuerola J (2010) Trophic experiments to estimate isotope discrimination factors. J Appl Ecol 47:948–954
Collins AL, Walling DE, Webb L, King P (2010) Apportioning catchment scale sediment sources using a modified composite fingerprinting technique incorporating property weightings and prior information. Geoderma 155:249–261
DeNiro MJ, Epstein S (1976) You are what you eat (plus a few ‰): the carbon isotope cycle in food chains. Geol Soc Am Abstr Program 8:834–835
Dubois S, Jean-Louis B, Bertrand B, Lefebvre S (2007) Isotope trophic-step fractionation of suspension-feeding species: implications for food partitioning in coastal ecosystems. J Exp Mar Biol Ecol 351:121–128
Flaherty E, Ben-David M (2010) Overlap and partitioning of the ecological and isotopic niches. Oikos 119:1409–1416
Fry B (2006) Stable isotope ecology, 1st edn. Springer, New York
Galván D, Botto F, Parma AM, Bandieri L, Mohamed N, Iribarne O (2009) Food partitioning and spatial subsidy in shelter-limited fish species inhabiting patchy reefs of Patagonia. J Fish Biol 75:2585–2605
Gibbs MM (2008) Identifying source soils in contemporary estuarine sediments: a new compound-specific isotope method. Estuar Coasts 31:344–359
Hussey NE, MacNeil MA, Fisk AT (2010) The requirement for accurate diet-tissue discrimination factors for interpreting stable isotopes in sharks. Hydrobiologia 654:1–5
Jackson AL, Inger R, Bearhop S, Parnell A (2009) Erroneous behaviour of MixSIR, a recently published Bayesian isotope mixing model: a discussion of Moore & Semmens (2008). Ecol Lett 12:E1–E5
Logan JM, Lutcavage ME (2010) Reply to Hussey et al.: the requirement for accurate diet-tissue discrimination factors for interpreting stable isotopes in sharks. Hydrobiologia 654:7–12
Lubetkin SC, Simenstad CA (2004) Multi-source mixing models to quantify food web sources and pathways. J Appl Ecol 41:996–1008
Martinez del Río C, Wolf B (2005) Mass-balance models for animal isotopic ecology. In: Starck JM, Wang T (eds) Physiological and ecological adaptations to feeding in vertebrates. Science Publishers, Enfield, pp 142–174
Martinez del Río C, Wolf N, Carleton SA, Gannes LZ (2009) Isotopic ecology ten years after a call for more laboratory experiments. Biol Rev 84:91–111
McCutchan JH, Lewis WM, Kendall C, McGrath CC (2003) Variation in trophic shift for stable isotope ratios of carbon, nitrogen, and sulfur. Oikos 102:378–390
Melville AJ, Connolly RM (2003) Spatial analysis of stable isotope data to determine primary sources of nutrition for fish. Oecologia 136:499–507
Moore JW, Semmens BX (2008) Incorporating uncertainty and prior information into stable isotope mixing models. Ecol Lett 11:470–480
Parnell A, Jackson A (2010) SIAR: stable isotope analysis in R. R package version 4.0.2. http://CRAN.R-project.org/package=siar
Parnell A, Inger R, Bearhop S, Jackson AL (2010) Source partitioning using stable isotopes: coping with too much variation. PLoS ONE 5:e9672
Perga M-E, Grey J (2010) Laboratory measures of isotope discrimination factors: comments on Caut, Angulo & Courchamp (2008, 2009). J Appl Ecol 47:942–947
Phillips DL (2001) Mixing models in analyses of diet using multiple stable isotopes: a critique. Oecologia 127:166–170
Phillips DL, Gregg WJ (2003) Source partitioning using stable isotopes: coping with too many sources. Oecologia 136:261–269
Pinnegar J, Polunin NVC (2000) Contributions of stable-isotope data to elucidating food webs of Mediterranean rocky littoral fishes. Oecologia 122:399–409
Post D (2002) Using stable isotopes to estimate trophic position: models, methods, and assumptions. Ecology 83:703–718
R Development Core Team (2010) R: a language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria. http://www.R-project.org. ISBN: 3-900051-07-0
Rubin D (1988) Using the SIR algorithm to simulate posterior distributions. In: Bernardo J, Degrood M, Lindley D, Smith A (eds) Bayesian Statistics 3: proceedings of the third Valencia international meeting, June 1–5, 1987. Clarendon Press, Oxford, pp 385–402
Schindler DE, Lubetkin SC (2004) Using stable isotopes to quantify material transport in food webs. In: Polis GA, Power ME, Huxel GR (eds) Food webs at the landscape level. University of Chicago Press, Chicago, pp 25–42
Sweeting CJ, Barry JP, Barnes C, Polunin NVC, Jennings S (2007a) Effects of body size and environment on diet-tissue δ15N fractionation in fishes. J Exp Mar Biol Ecol 340:1–10
Sweeting CJ, Barry JP, Polunin NVC, Jennings S (2007b) Effects of body size and environment on diet-tissue δ13C fractionation in fishes. J Exp Mar Biol Ecol 352:165–176
Vander Zanden M, Shuter B, Lester N, Rasmussen J (1999) Patterns of food chain length in lakes: a stable isotope study. Am Nat 154:406–416
Wolf N, Carleton S, Martinez del Río C (2009) Ten years of experimental animal isotopic ecology. Funct Ecol 23:17–26
Acknowledgments
Funding was provided by a postdoctoral fellowship from Concejo Nacional de Investigación Científica y Técnica (CONICET) and CONICET/R09/639 (D.E.G.) and Natural Environment Research Council grant NE/D013437/1 (C.J.S.) and Agencia Nacional de Promoción Científica y Tecnológica PICT 2008-1433 (D.E.G.). Comments of A. Parnell (School of Mathematical Sciences, University College Dublin) on SIAR performance greatly improved the manuscript.
Author information
Authors and Affiliations
Corresponding author
Additional information
Communicated by Robert Hall.
Electronic supplementary material
Below is the link to the electronic supplementary material.
Rights and permissions
About this article
Cite this article
Galván, D.E., Sweeting, C.J. & Polunin, N.V.C. Methodological uncertainty in resource mixing models for generalist fishes. Oecologia 169, 1083–1093 (2012). https://doi.org/10.1007/s00442-012-2273-4
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00442-012-2273-4