
Abstract
The high diversity of phytophagous insects has been explained by the tendency of the group towards specialization; however, generalism may be advantageous in some environments. The cerambycid Apagomerella versicolor exhibits intraspecific geographical variation in host use. In northern Argentina it is highly specialized on the herb Pluchea sagittalis (Asteraceae), while in central and southern areas it uses seven Asteraceae species. To study host species geographical variation from ecological and evolutionary perspectives, we investigated field host availability and use across a wide latitudinal range, and performed laboratory studies on insect oviposition preference and larval performance and mitochondrial DNA (mtDNA) variation in a phylogeographical framework. Geographic variation in host use was unrelated to host availability but was highly associated with laboratory oviposition preference, larval performance, and mtDNA variation. Genetic studies revealed three geographic races of A. versicolor with gene flow restriction and recent geographic expansion. Trophic generalism and oligophagy within A. versicolor seem to have evolved as adaptations to seasonal and spatial unavailability of the preferred host P. sagittalis in cooler areas of the species’ geographic range. No single genotype is successful in all environments; specialization may be advantageous in environments with uniform temporal and spatial host availability, while being a trophic generalist may provide an adaptive advantage in host-constrained environments.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Plant-feeding insects comprise a highly diverse group that is estimated to encompass about 30 million species (Erwin 1982), which is more diverse than its sister nonphytophagous clades (Mitter et al. 1988). This difference has been attributed to the tendency of the group towards specialization and to undergo host plant shifts resulting in host-race formation followed by speciation (Craig et al. 1993; Bush 1994). In addition, phytophagous insects exhibit a kind of evolutionary fidelity known as taxonomic conservation: shifts to host plants that are phylogenetically related to plants used by the insect lineage in the past are more common than shifts to unrelated plants (Futuyma et al. 1993; Janz et al. 2001). Geographical host species changes in phytophagous insects are often related to changes in feeding or oviposition preference (Futuyma 1983). The study of host range dynamics in herbivorous insects can help to elucidate the underlying mechanisms involved in the generation and maintenance of biodiversity, which is a central goal of evolutionary biology (Futuyma and Moreno 1988; Jaenike 1990).
Specialization in phytophagous insects has been explained as being the result of an optimization process related to the use of a host plant, which constrains insect performance in other hosts (Futuyma and Moreno 1988; Mopper et al. 1995). Specialization may also provide adaptive advantages (Smith 1988; Craig et al. 1993; Groman and Pellmyr 2000), such as escape from natural enemies (predation/parasitism) (Bernays and Graham 1988; Crespi and Sandoval 2000), the possibility of using a better habitat (Futuyma and Keese 1992), and/or adaptation to the chemical defenses of a plant species or group of plants (Dethier 1954; Fraenkel 1959; Ehrlich and Raven 1964; Futuyma and Keese 1992).
Most studies dealing with the evolution of host specificity reconstruct host use in groups of related insects by means of phylogenetic analysis (Kelley and Farrell 1998; Thompson 1998; Futuyma 2001; Janz et al. 2001), or study shifts to introduced hosts, e.g., Rhagoletis pomonella (Walsh) (Prokopy et al. 1988) and butterflies of the genera Euphydryas (Thompson 1993) and Papilio (Singer et al. 1993). However, some insect species exhibit geographical variation in host use, which may represent a recent or ongoing evolutionary process (Singer et al. 1992; Bigger and Fox 1997; Thompson 1998). The study of an insect’s intraspecific host species use variation may provide hints about the evolution of host use and insect diversification (Funk and Bernays 2001). The aphid Uroleucon ambrosiae, for instance, is monophagous on Ambrosia trifida (Asteraceae) in the east, which is its ancestral condition, and polyphagous on several species of Asteraceae in the west of its distribution in North America. Trophic generalism within U. ambrosiae may have evolved as an adaptation to the scattered and unpredictable distribution of the preferred host A. trifida in the most arid area of the insect distribution (Funk and Bernays 2001). Nevertheless, as in most studies of intraspecific variation in host use, they could not find a correlation between genetic variation and host use (Jaenike 1990; Funk and Bernays 2001), suggesting that the variation encountered could be an ecological response to host availability.
The stem borer Apagomerella versicolor (Coleoptera: Cerambycidae, subtribe Aerini) exhibits intraspecific variation in host use within its distribution range: it is highly specialized on Pluchea sagittalis Cabr in northern Argentina, while in the central and southern regions it is oligophagous on seven species of Asteraceae, including P. sagittalis (Logarzo and Gandolfo 2005). The geographic distribution of the cerambycid in Argentina covers a wide latitudinal range, from warm subtropical areas in the north (23° S) to Patagonia in the south (40° S), which encompasses wide variations in climate and vegetation. Life cycle variations also reflect this disparity in environmental conditions, since beetles from latitudes higher than 31° S are obligate diapausers (Logarzo et al. 2002; Logarzo and Gandolfo 2005), while northern populations (25–27° S) are at least bivoltine and do not diapause (Logarzo and Gandolfo 2005).
Apagomerella versicolor was formerly considered a candidate agent for the biological control of cocklebur (Xanthium cavanillesii) in the United States (Logarzo et al. 2002), but it was rejected because of its broad host range. The genus Apagomerella includes only two species, A. versicolor and A. dissimilis. Females lay only one egg per oviposition site, and there is always only one larva per stem. The larvae bore through the stem towards the root, where they pupate (Logarzo et al. 2002).
Our main questions regarding host range variation in A. versicolor were (1) is geographical variation in host species use an ecological response to host plant species availability, or is it an evolutionarily derived condition, and (2) if it is evolutionarily derived, which are the ecological factors involved?
To address these questions we studied intraspecific geographical variation in host use in A. versicolor, combining field studies of host plant use and availability with controlled experiments on oviposition preference, larval performance and genetic variation in a phylogeographical framework.
Materials and methods
Field studies
Host plants studied
We studied the seven host plant species reported for A. versicolor (Logarzo and Gandolfo 2005): Ambrosia tenuifolia Spreng, A. scabra, Pluchea sagittalis Cabrera, P. absinthioides DC, Xanthium cavanillesii Schouw, X. spinosum L, and X. strumarium L, all of which belong to the family Asteraceae and have erect single stems. Detailed information on the characteristics of these plants is given in Table 1. For A. tenuifolia, we only considered plant specimens with a stem diameter of >5 mm for the studies, since larvae of A. versicolor cannot develop in narrower stems (Logarzo et al. 2002).
Study area
Field studies of host plant availability and utilization were conducted between 1990 and 1994 and between 2003 and 2005. These studies covered all of the distribution area of the cerambycid in Argentina, between 23° S and 40° S (Logarzo et al. 2002). This area was divided into three zones based on previous studies on host plant occurrence and use (Logarzo and Gandolfo 2005). Zone 1 (approx. 23°–27° S): all hosts were present and P. sagittalis was the only plant attacked. Zone 2 (approx. 27°–38° S, Central and Eastern Argentina): all hosts were present and attacked. Zone 3 (approx. 31°–40° S, bordering west and south of zone 2): all hosts were present except P. sagittalis; P. absinthioides was the only plant species that was confirmed as being attacked (Fig. 1). Zones 1, 2 and 3 covered approx. 22, 62 and 16%, respectively, of the total studied area.

Zonification of the distribution range of A. versicolor in Argentina according to information on host plant species occurrence and their use by A. versicolor, as gathered from previous studies and observations. In zone 1, all host plant species except A. scabra were present but only Pluchea sagittalis was used by A. versicolor; in zone 2, all seven host plant species were present and they were all used; in zone 3, P. absinthioides, X. cavanillesii and X. spinosum were present, but only the former was used
The geographical distribution of A. versicolor comprises three ecoregions: Pampa grassland, Chaco forest and Monte shrubland (Logarzo et al. 2002). Zone 1 occupies the warmest and most humid areas of northern Chaco, zone 2 is located in southern Chaco and Pampa, while zone 3 is in the Monte. The Chaco is a savanna that has thorn forests dominated by Schinopsis spp. (Anacardiaceae), Aspidosperma (Apocynaceae) and Prosopis spp. (Mimosoideade) interspersed in grasslands, with different degrees of dominance of both. Mean annual temperatures decrease from 23°C in the north to 21°C in the south, and mean annual precipitations range from 1,300 mm in the east to 500 mm in the west, with the precipitation occurring mostly in the summer (Cabrera 1994). The Pampas (sensu Morrone 2006) originally consisted of temperate grasslands, savannas, shrublands or xeric woodlands, all of which are now mostly crop ecosystems. The dominant grasses of the Pampas belong to the genera Stipa, Piptochaetium, Aristida, Melica, Briza, Bromus, Eragrostis and Poa. The climate is mild (19.7–13.4°C), with precipitation of 600–1,200 mm, distributed more or less evenly throughout the year. The Monte ecoregion is dominated by scrublands of resinous evergreen bushes, mostly in the Zygophyllaceae family, interspersed in a matrix of grasses, herbs and bare soil. The climate is temperate–arid (17.5–13.4°C) with very little rainfall (between 80 and 250 mm per year), which occurs in summer in the north or in the winter in the south. There is some evidence that the Monte and the Pampa represent an impoverished Chaco (Cabrera 1994; Morrone 2006).
Host plant availability
Host plant species availability was estimated along 85 transects (100 km each) randomly located in the three zones. All transects were sampled between September and April (spring and fall in the southern hemisphere); most of them between the years 1990 and 1994, and a few between 2003 and 2005. The number of transects surveyed in each zone was proportional to its area: 17 (20.0%) in zone 1; 54 in zone 2 (63.5%); and 14 in zone 3 (16.5%), covering the distributional range of the cerambycid (between latitudes of 23° S and 40° S). For each of the seven host plant species of A. versicolor, we recorded the number of patches and the number of plant specimens within each patch (all host plant species studied grow clumped in monospecific patches) at five sampling points placed every 20 km along each transect. All host plant species patches located within a 300 m distance of the sampling point were counted and recorded. Sampling points located at river sides, road sides, etc., were included in the study, since those are suitable habitats for the studied host plant species (see Table 1). Local weather during the field studies was not recorded.
To evaluate if the host plant species differed in their availabilities within each zone, we used a generalized linear model analysis with a Poisson error distribution and a log link function, using host plant species as the explanatory variable and plant species abundance as the response variable. Analyses were conducted using the nlme function in the lme4 package of the R statistical software package, version 2.4.0 (http://www.r-project.org). We used Akaike’s information criterion (AIC) to compare the fits of different models. For each zone, we started with a model that compared the availabilities of all host plant species present in the respective zone, and then we simplified the model by excluding the host plant species that did not contribute to the model. This simplification was considered to be justified only if the associated AIC value was at least two units lower than the AIC associated with the previous model (Burnham and Anderson 2002).
We also checked model deviance for overdispersion using the “estimated scale” parameter, Φ, returned by nlme (i.e., the model residual deviance over the residual degrees of freedom), which indicates departure from the assumed Poisson error distribution (Crawley 2005). A model is overdispersed if Φ > 1, and underdispersed if Φ < 1. When we found that the value of Φ departed substantially from 1 for a given model, we re-ran the analysis using the quasi-Poisson distribution (by setting family = quasipoisson in the lmer function call).
The relationship between host plant species (P. sagittalis, P. absinthioides, X. cavanillesii, X. spinosum and A. tenuifolia) availability and latitude was analyzed by linear regression analyses (SPSS 11.5; SPSS, Inc., Chicago, IL, USA), using abundance as the response variable and latitude as the fixed independent variable. To perform the regression, the number of patches observed in a degree latitudinal band was grouped and standarized to 100 km (in order to overcome the unbalanced number of transects per latitudinal band).
Host plant utilization
The numbers of attacked patches and attacked plants per patch of each host plant species were recorded between April and November (to ensure larval or pupal presence) at the same sampling points used for the estimation of host plant availability. At each patch, 30–100 plants (depending on patch size) were dissected and inspected for the presence of A. versicolor larvae. Pupae found in the stumps were also recorded. All cerambycid larvae and pupae collected were reared to adulthood on a meridic diet (Logarzo et al. 2002) to confirm insect identification. When larvae were found dead inside the host plant, they were collected and identified in the laboratory. The damage produced by the larvae was also used for identification purposes.
The host plant utilization of A. versicolor was analyzed using the Neu method (Neu et al. 1974). This method compares host plant species availability with the actual proportion of plant individuals of that species used by A. versicolor, using a chi-square goodness of fit test. Critical assumptions of the method are that all of the insect observations are independent, and that the availability of each host plant is the same for all A. versicolor specimens. In this study, availability was measured as the total number of plants of each species counted. Two null hypotheses were tested: (1) A. versicolor utilizes plant species in proportion to their availability, considering all of the host plant species simultaneously; (2) A. versicolor utilizes plant species in proportion to their availability, considering all of the host plant species separately.
When a difference in plant utilization was detected (null hypothesis 1), simultaneous confidence intervals (CI) were calculated using a Bonferroni t-statistic,
where \( \hat{p}_{j} \) is the proportion of plants utilized by A. versicolor and α/2k is used to ensure that k (= number of host plant species analyzed) simultaneous CIs have an overall α = 0.05.
Three categories of interaction were established: (1) “preferred,” where A. versicolor utilized a host plant species more than its availability, i.e., the CI lower limit was above the plant species’ availability; (2) “used in proportion,” where the insect utilized a host plant species in proportion to its availability, i.e., the plant species availability was within the CI for the proportion used by the cerambycid, and (3) “used less,” where the insect utilized a host plant species less than its availability, i.e., the upper CI limit was below the plant species’ availability.
Data from zones 1 and 3 were not analyzed because the cerambycid used only one host plant species in those zones.
Laboratory studies
We explored oviposition preference, larval performance and genetic variation associated with A. versicolor geographical variation in host utilization. We used A. versicolor adults that emerged in the laboratory from larvae and pupae collected in the field. Last instars (sixth or seventh) were obtained by dissecting host plant stems and stumps. Pupae were found only in roots or stumps. Last instars and pupae were reared individually until adult emergence in 34-ml plastic cups with paper lids that were filled with moist tissue paper (3/4 of total volume) to provide a humid substrate. All insects were held at 25–30°C, 70–80% RH, with a 14–10 L:D photoperiod. Some host plant species yielded very few larvae and could not be used for some (A. tenuifolia) or any (X. strumarium, X spinosum, A. scabra) laboratory studies.
Oviposition preferences and larval performance test
We performed multiple choice tests to investigate if oviposition preferences and/or larval performance were affected by the geographical and host plant origins of females. Oviposition preference could not be measured independently from larval performance, because it was not possible to determine whether or not A. versicolor laid an egg in a plant specimen, given that the eggs are endophytic and females frequently introduce their ovipositor into the stem (leaving a hole: an oviposition mark) but do not lay any eggs (Gandolfo et al. 1997). The response variable used in this test was therefore the presence of an emerged adult of A. versicolor (only one larva can survive per plant) at each offered host plant specimen. Thus, this study integrates female preference for ovipositing on a given host species and larval aptitude for reaching the adult stage on that host.
A female was offered four host plant species simultaneously: P. sagittalis, X. cavanillesii, A. tenuifolia, and P. absinthioides from which it could choose freely one or more plant species in which to oviposit. In these experiments, we used mated females derived from the following zones and the following host plant species attacked in each zone: P. sagittalis from zones 1 and 2; X. cavanillesii and A. tenuifolia from zone 2; and P. absinthioides from zones 2 and 3. Five to 15 replicates were run per type of female, resulting in a total of 61 cages. Each female was placed in a 1 m side cage with voile screen walls containing one specimen of each of the four host plants. After 24 h, the females were removed and the plants were labeled and isolated to avoid infestation with wild adults. At the end of the summer, when the larvae reached their last instars, the plants were dissected and the presence of A. versicolor was recorded. The females used in these tests had been previously mated with insects from the same zone and host plant. All host plant species specimens used in the tests were collected, either as plants (A. tenuifolia, P. saggitalis, P. absinthioides) or as seeds (X. cavanillesii) in zone 2, and were grown inside mesh cages to avoid attack by free-living A. versicolor females.
Genetic differentiation among zones
The degree of genetic differentiation was investigated using samples of adults collected in the three zones. Specimens from different collecting sites in Corrientes and Formosa Provinces in zone 1 (n = 17) were pooled due to the scarcity of individuals per site. In zone 2 (n = 50), five different localities were sampled due to its broad latitudinal range: Gualeguaychú and Colón (Entre Ríos Province); Paso de los Libres (Corrientes Province); and Mar del Plata and Reta (Buenos Aires Province). Specimens from zone 3 (n = 11) were collected from the locality of Río Colorado (Río Negro Province).
Total DNA was extracted from adult specimens using the Puregene® DNA Purification Kit (Gentra System) according to the manufacturers protocol. A 629 bp fragment of the mitochondrial cytochrome oxidase subunit I (COI) gene was PCR amplified and sequenced as described in Piccinali et al. (2009). New COI sequences were deposited in Genbank (accession numbers EU232664–EU232691).
Nucleotide diversity was estimated using Watterson’s estimator (θ W) and the average number of pairwise differences observed per site (π). Haplotype variation was estimated using the number of haplotypes (h) and haplotype diversity (Hd). The tests of Tajima (1989), Fu (1997) and Ramos-Onsins and Rozas (2002) were applied to determine if the populations of A. versicolor were in genetic drift–mutation equilibrium. The significance of each test was calculated after performing 1,000 coalescent simulations based on a Monte Carlo process with no recombination (Hudson 1990). All calculations were carried out using DnaSP 5.0 (Librado and Rozas 2009).
Population structure was analyzed by means of an analysis of molecular variance (AMOVA, Excoffier et al. 1992). Sequences were arranged in two hierarchical levels: (1) site of collection (population), and (2) the zone to which the populations belong. The significances of the variance components and the Ф statistics were calculated by means of 10,000 permutations of haplotypes between populations and zones using the Arlequin 3.1 software (Excoffier et al. 2005). Genetic differentiation between pairs of zones was also tested with K *ST statistics (Hudson et al. 1992), as implemented in DnaSp 5.0 (Librado and Rozas 2009). Phylogenetic relationships between COI haplotypes were analyzed using a maximum parsimony approach. The shortest trees were found with the implicit enumeration search option implemented in TNT (Goloboff et al. 2008). All characters were regarded as unordered and unweighted because all parsimony-informative nucleotide changes were synonymous transitions. Statistical support for clades was assessed by means of a bootstrap method (Felsenstein 1985) based on 1,000 replicates and Bremer support (Bremer 1994). Phylogeographic patterns were analyzed with nested clade analysis (NCA) (Templeton et al. 1992) using the automation approach implemented in ANeCA (Panchal 2007), which includes the TCS 2.1 (Clement et al. 2000) and GeoDis 2.5 (Posada et al. 2000) programs.
Results
Field studies
Host plant availability
We recorded 247 patches of the seven host plant species at 425 sampling points. Four host species accounted for 88% of the observed host patches: X. cavanillesii (107 patches, 43%), P. sagittalis (58 patches, 23%), P. absinthioides (29 patches, 12%), and A. tenuifolia (25 patches, 10%) (Table 2a).
In zone 1, all hosts were present with the exception of A. scabra. Pluchea sagittalis and X. cavanillesii had similar availabilities and were more abundant than the remaining four host plant species (z = 2.419, P = 0.0156, Φ = 0.75), according to the simplest model fit (AIC = 166.58) (Table 2a).
In zone 2, the seven host species were present and, according to the simplest model fit (AIC = 520.29), the most abundant host was X. cavanillesii (z = 4.261, P = 0.000), followed by P. sagittalis (z = 3.618, P = 0.000), Ambrosia tenuifolia, and P. absinthioides, which had similar availabilities (z = 2.629, P = 0.008), and were more abundant than X. spinosum, X. strumarium, and A. sacra, which did not differ in their availabilities (z = 1.535, P = 0.1248) (Table 2a).
In zone 3, four hosts were present: X. cavanillesii, P. absinthioides, X. spinosum, and A. tenuifolia. According to the simplest model fit (AIC = 129.04, vs. 130.57), Xanthium cavanillesii and P. absinthioides had similar availabilities and were more abundant than X. spinosum and A. tenuifolia (z = 2.880, P = 0.004, Table 2a).
The three zones showed different patterns of numbers of plants per patch. In zone 1, all host species had similar numbers of plants per patch (F = 0.495, df = 5, 44, P = 0.778). In zone 2, P. absinthioides had more plants per patch than the other hosts, which had similar numbers per patch (F = 6.522, df = 5, 147, P = 0.000) (Table 2a). In zone 3, P. absinthioides had more plants per patch than X. spinosum, but neither of these differed significantly from the other two hosts, X. cavanillesii and A. tenuifolia (F = 4.336, df = 3, 39, P = 0.01) (Table 2a).
Regression analyses showed that the availabilities of P. sagittalis (R 2 = 0.80, df = 1, 12, F = 54.4, P = 8.6 × 10−6) and P. absinthioides (R 2 = 0.66, df = 1, 12; F = 23.38; P = 0.000) were negatively and positively associated with latitude. In contrast, the availabilities of X. cavanillesii (R 2 = 0.07, df = 1, 12, F = 0.96, P = 0.34) and X. spinosum (R 2 = 0.03, df = 1, 12, F = 0.45, P = 0.51) were unrelated to latitude (Fig. 2).

Relationship between the availabilities of P. sagittalis, P. absinthioides, X. cavanillesii, and X. spinosum and latitude, assessed through linear regressions. Host availability was estimated as the number of stands observed every 100 km in 1° latitudinal bands. The regressions of X. cavanillesii and X. spinosum were not significant
Host plant utilization
A. versicolor host plant species use varied according to geographic area. In zone 1, all 19 patches of P. sagittalis were attacked, with an average of >50% of plants attacked per patch, while the patches of the other five host plant species (n = 31) exhibited no infestation by the cerambycid (Table 2b).
In zone 2, all host plant species were present, and their patches were attacked by the insect in proportion to their availability (X 2 = 5.20, df = 5, P = 0.54) (Table 2b). However, P. sagittalis was the preferred host plant in terms of number of plants used [proportion of plants available (PPA) = 0.208; confidence interval of proportion of insect use (CI) = 0.327–0.388]; while Xanthium cavanillesii (PPA = 0.493, CI = 0.474–0.538) and A. scabra (PPA = 0.007, CI = 0.001–0.012) were used in proportion to plant availability, and the remaining hosts, X. spinosum (PPA = 0.035, CI = 0.001–0.012), X. strumarium (PPA = 0.020, CI = 0.000–0.008), A. tenuifolia (PPA = 0.089, CI = 0.026–0.050) and P. absinthioides (PPA = 0.141, CI = 0.053–0.085), were used less than predicted based on plant availability.
Although four host plant species were present in zone 3 (P. absinthioides, X. cavanillesii, X. spinosum and A. tenuifolia), only P. absinthioides was attacked (86.7% of sampled patches, n = 15) (Table 2b), with an average of 24.7% (SD = 25.9) of the plants attacked per patch.
Laboratory studies
Oviposition preference and larval performance tests
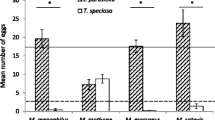
Oviposition preference and/or larval performance varied according to the host plant and the geographic origin of the tested females (Table 3). Females collected in zone 1 oviposited and larvae reached adulthood only on P. sagittalis (100% of the offered plants); females from zone 2 produced adults on P. sagittalis, X. cavanillesii and A. tenuifolia (90.2, 48.8, and 4.9%, of the offered plants, respectively); and females from zone 3 produced adults on P. sagittalis and to a lesser extent on P. absinthioides (Table 3).
Genetic differentiation among zones
Total nucleotide diversity was 0.0113 when measured on the basis of the number of segregating sites (θ W), and 0.005 when expressed in terms of the mean number of pairwise nucleotide differences per site (π) (Table 4). Thirty-five polymorphic sites (five nonsynonymous and thirty synonymous) were detected in the entire sample; 14 of these were parsimony-informative (Table 4). The total number of haplotypes was 28, with a total haplotype diversity (Hd) of 0.891. Most haplotypes were specific to each zone, with only three haplotypes being shared by zones 1 and 2 and one shared by zones 1 and 3 (Table 4).
The level of nucleotide variation differed greatly among zones. Zone 1 was the most diverse, followed by zones 2 and 3. The latter was much less variable than the other two (Tables 4, 5). Although zones 1 and 2 displayed rather similar nucleotide diversities, the former had higher values of π and Hd. Indeed, the mean number of pairwise differences among haplotypes was greater in zone 1 than in zone 2, and their frequencies were more even.
Levels of genetic differentiation among populations of the different zones were highly significant (σ 2a = 1.06, Ф CT = 0.52, df = 2, P = 0.009), accounting for 51% of the total sequence variation, whereas differentiation between populations within zones was low and nonsignificant (σ 2b = 0.04, Ф SC = 0.04, df = 5, P = 0.127). Pairwise comparisons revealed that zones 1 and 2 were slightly but significantly differentiated (K *ST = 0.063, P < 0.0001), but that zones 1 and 2 were not significantly differentiated from zone 3 (zone 1–zone 3: K *ST = 0.349, P < 0.0001; zone 2–zone 3: K *ST = 0.332, P < 0.0001).
Neutrality tests revealed significant departures from neutral expectations in populations of A. versicolor (Table 5). D T was significant and negative for the total sample and zone 2, while Fs and R 2 were significant for the total sample, zone 1 and zone 2.
The parsimony analysis found 20 most parsimonious trees of 138 steps. The strict consensus tree is shown in Fig. 3. Interestingly, haplotype m, recorded in zone 1, was the first that branched off in all trees, suggesting that it represents an old variant that is still segregating in ancestral populations. The remaining variants clustered into two clades, one including variants from zones 1 and 3 and the other comprising haplotypes from zones 1 and 2. However, only the first group had bootstrap support of >50% and high Bremer support (Fig. 3).

Strict consensus of the 20 most parsimonious trees of A. versicolor COI haplotypes. Numbers above and below the branches are the ≥50% bootstrap and Bremer support values, respectively. The phylogenetic tree was drawn with MrEnt 2.1 (Zuccon and Zuccon 2010)
The haplotype network (Fig. 4) showed only one ambiguous connection, indicative of either homoplasy or (less probably) recombination. The most frequent haplotype (a), which occupied a central position in the network, was found in 23 individuals spanning all localities sampled in zone 2, and the surrounding haplotypes differing in one or two mutational steps from a, were detected in either zone 2, zone 1, or both. Remarkably, two groups of divergent haplotypes separated from haplotype a by four or five mutational steps can be observed in the network (Fig. 4). One of these groups contained haplotypes z and x, which were only found in northern localities of zone 2, while the other group involved haplotypes n (shared between zones 1 and 3) and u (exclusive to zone 3) (Fig. 4). In agreement with the maximum parsimony analysis, haplotype m was separated by six or seven mutational steps from the rest. The NCA indicated significant associations between haplotypes and geographical locations (Table 6, Fig. 4). In particular, clades 1–8 and 2–5 as well as the total cladogram showed patterns compatible with long-distance colonization and/or past fragmentation, while clade 3–2 suggest restricted gene flow with isolation by distance.

Nested cladistic analysis of the A. versicolor COI haplotype network. Circles with letters are recorded haplotypes. Patterns represent the geographic areas in which the haplotypes were found. Z1, zone 1; Z2, zone 2; Z3, zone 3. Small open circles are inferred intermediate haplotypes that were not sampled in the study. Each line is a single mutational change. Boxes enclose haplotypes at each nested level. Numbers identify each nested clade. An asterisk means a significant association between the geographic and genetic distances in that clade
Discussion
Is geographical variation in host use an ecological response to host plant availability, or is it an evolutionarily derived condition?
The cerambycid Apagomerella versicolor exhibits wide geographical variation in host species utilization in nature (Logarzo and Gandolfo 2005), ranging from monophagy on P. sagittalis or P. absinthioides in zones 1 and 3, respectively, to oligophagy in zone 2 (Table 2b). Such intraspecific variation in host use was not related to host availability in the field (Table 2a, Fig. 2). If that was the case, the cerambycid should attack Xanthium cavanillessi in zones 1 and 3, where this host plant species is as abundant as the attacked Pluchea hosts, and should use X. cavanillessi more than P. sagittalis at zone 2, given that the former is more abundant in that area. Thus, our study suggests that host range variation in A. versicolor is not due to an ecological response to host availability, as has been observed in other studies (Bernays and Graham 1988).
An alternative explanation for the differences in host plant use vis a vis host availability is that there is some kind of genetic determination of host use (i.e., females of different geographic areas differ in oviposition host preference). The consistency found between field host plant use and insect host adaptation, measured as laboratory host use (Table 7), suggests that geographic differences in host use are not due to a genotype–environment interaction or phenotypic plasticity (Via 1984; Via and Lande 1985), but rather a genetically determined trait. In fact, the net results for oviposition preferences and larval performance in the laboratory choice test closely resembled patterns of host plant use in the field, with the apparent exception of females of zone 3 (Tables 2b, 3). Females from zone 3 preferred P. sagittalis over P. absinthioides in laboratory assays (Table 3), while in the field the cerambycid was only recovered from the latter. Interestingly, females of zone 3 showed a greater preference for P. sagittalis, a host which they never encounter in the field, since this host plant is absent in zone 3 (Table 2a). This preference for P. sagittalis exhibited by zone 3 females may represent the ancestral state in A. versicolor. Moreover, it has been argued that phytophagous insects that acquire novel hosts can maintain their fitness on ancestral hosts during phases of host range expansion or shifts (Futuyma et al. 1995; Janz et al. 2001).
The differences observed in laboratory experiments for oviposition preference and larval performance could also be attributed to a larval-host plant learning effect that affects female oviposition behavior, as previously observed in lepidopterans (Papaj 1986; Cunningham et al. 1998). However, the host plant origin of the female had only a minor (i.e., on females from A. tenuifolia) or no effect in our assay (Table 3). The results of the laboratory choice test were similar for females recovered from four different host plants (X. cavanillesii, A. tenuifolia, P. sagittalis, and P. absinthioides) in zone 2, suggesting that the niche experienced during larval development does not influence oviposition behavior.
Taken together (Table 7), our results strongly support the hypothesis that geographical variation in host use has a genetic basis. Differences in oviposition preferences causing host use changes are generally followed by host race formation or geographical differentiation, leading eventually to reproductive isolation (Itami et al. 1998; Wood and Keese 1990; Janz et al. 2006). The mtDNA genetic data clearly support genetic differentiation among the three studied zones, and provide strong support to the idea of a recent population expansion in this species, particularly within zones 1 and 2. This hypothesis was tested using several specific tests, such as Tajima’s D, Fu’s Fs, and Ramos-Onsins and Rozas’ R 2 tests, which are very powerful methods of detecting departures from neutral expectations due to changes in historical population size (Fu 1997; Ramos-Onsins and Rozas 2002). However, when applied to zone 3, these tests yielded nonsignificant results, which could be due to either the low number of segregating sites detected in this sampling area or a strong founder effect following a recent colonization.
The phylogeographic analysis agrees with genetic differentiation caused by restrictions to gene flow in populations of A. versicolor coupled with long-distance colonization events and/or past habitat fragmentation. The fact that zones 2 and 3 shared haplotypes with zone 1 but not with each other (despite being geographically close) means that insects from zone 3 are more closely related to insects from zone 1 than those from zone 2, and suggests two independent routes of colonization.
Three features of the mtDNA data suggest that zone 1 may be ancestral to the others. First, it has the highest nucleotide and haplotype diversity; second, it shares haplotypes with the other two zones; and third, it had a highly divergent haplotype in the maximum parsimony tree, which separates early from the remaining variants. However, it should be noted that haplotypes found in zone 1 are scattered across the haplotype network, whereas those from zone 2 are central and thus could also represent the ancestral type (Fig. 4).
Ecological factors involved in the evolution of geographically related variation in host plant use by A. versicolor
Our mtDNA data analyses showed that the three zones are genetically structured. Thus, we may ask whether the observed geographic structure is related to patterns of host use. Phytophagous, endophytic insects with restricted dispersal ability (Karban 1989; Mopper 1996; Stiling and Rossi 1998; Van Zandt and Mopper 1998) like A. versicolor have a tendency to adapt to host plant species, thus resulting in the development of locally adapted populations (Williams 1966; Kawecki and Ebert 2004). When gene flow among populations does not counteract the forces of selection (Alleaume-Benharira et al. 2006), adaptive divergence takes place. However, other evolutionary processes may also cause differentiation, such as sexual selection (Lande 1981) or conflict (Chapman et al. 2003), chance events due to a small local population or bottlenecks (Choe and Crespi 1997; Arnqvist 1998; Matsubayashi et al. 2010), or the fixation of divergent incompatible alleles (Schluster 2009). Therefore, more studies using unlinked molecular markers would help to disentangle the possible mechanisms involved in the geographic differentiation of A. versicolor.
When divergent selection is caused by ecological sources (Schluter 2000, 2009), the process of host race formation is defined as “ecological speciation” (reviewed in Matsubayashi et al. 2010). Different host plant species can be considered different “environments” because they may vary in their morphology, chemistry or abundance, exposing the insects to divergent selective pressures (Matsubayashi et al. 2010). In the case of A. versicolor, our behavioral data suggests that P. sagittalis is its putative ancestral host, since it was the most commonly used host plant in the field regardless of its availability, and it was the preferred host in the oviposition experiments irrespective of the geographic origin of the females, even though this plant does not occur in zone 3 (Table 2a, b). Apparently, host range evolution in A. versicolor preserved the order of preference (Carriere and Roitberg 1996) (i.e., preference for P. sagittalis is maintained). Species that lay endophytic eggs like A. versicolor are assumed to be more limited in their searches for hosts (Carrière 1998), due to correlations between the size of the insect’s body, the size of the eggs, and the size of the host plant specimen. In fact, it was observed that small host plant specimens, regardless of the plant species, were not attacked by A. versicolor because their narrow stems cannot hold the cerambycid larvae (Logarzo 2007).
Considering that P. sagittalis is the ancestral host, which ecological factors were involved in the host expansion in zone 2 and the host shift in zone 3, as well as the subsequent insect race formation? Host expansion may be explained by an increase in herbivorous fitness due to an improvement in nutrient balance or food abundance (Westoby 1977; Berner et al. 2005) or due to the dilution of poisonous compounds (Freeland and Janzen 1974; Bernays and Mikenberg 1997) caused by feeding on several species or on different parts within the same plant (Johns et al. 2009)—also known as “dietary mixing.” However, the larva of A. versicolor is a stem borer that is confined to a single stem of a host plant specimen until it pupates inside the plant (Logarzo et al. 2002), so it cannot experience the benefits of intra- or interspecific host plant dietary mixing.
Some host plant species traits can exhibit geographical variation in ecological traits like nutritional quality and/or defense traits (Dethier 1954; Scriber and Slansky 1981; Lucas et al. 2000; Singer 2001), or in host plant morphology or chemical traits that provide refuge or natural enemy escape (Jeffries and Lawton 1984; Chew and Courtney 1991). We did not measure the geographical variation of P. sagittalis in relation to nutritional quality or chemical defenses. However, almost all P. sagittalis patches sampled were attacked by A. versicolor (56 out of 58) along a wide geographical distribution, meaning that if there is spatial variation in host traits, it does not affect insect use. In addition, due to its obligate endophagous feeding, A. versicolor larvae are rarely exposed to predators and entomopathogens. In fact, natural enemy mortality in this cerambicyd is mainly due to two species of parasitoids (Nealiolus sp. and Monitoriella sp, Hymenoptera: Braconidae), with a low percentage of attack (7.2% of 2,826 larvae sampled) (GAL, unpublished data).
If A. versicolor expanded or shifted to other host plant species than P. sagittalis to look for enemy-free space, P. sagittalis should be attacked by A. versicolor to a lesser extent in the areas where the expansion or shift was produced, i.e., in Zone 2. However, our data showed that, when present, P. sagittalis is attacked by A. versicolor. Ecological traits can interact, leading to tritrophic interactions (Price et al. 1980; Dicke 2000) and opposite selective pressures, resulting in trade-offs; i.e., larger lepidopteran larvae can eat tougher leaves (Fox and Czesak 2000) but they are more predated (Heimpel et al. 1996; Harvey et al. 2000). Apparently, the ecological advantages provided by the host P. sagittalis seem to counteract other probable selective pressures.
Taking the results together, our study shows that, when present, the host plant species P. sagittalis is always attacked at high rates by A. versicolor, despite some possible geographical variation in its ecological traits or in the enemy-free space provided. An alternative explanation to dietary mixing, host geographical variation or host enemy-free space is that geographic variation in host availability (Nitao et al. 1991) is the ecological factor that promoted local adaptation and divergent selection. Ecological availability of the preferred host P. sagittalis may have caused rapid evolutionary divergence (Schluter 2000) in A. versicolor populations. In northern Argentina (zone 1, north of 27° 20′ S), the perennial herb P. sagittalis is abundant and available throughout the year in frost-free habitats like stream and river shores (Cabrera 1994) (Argentina has no frost-free areas, with the exception of some ecological habitats; Burgos 1963). In central-southern cooler areas (south of 27° 20′ S, zones 2 and 3), P. sagittalis is less abundant or absent and, like the other three perennial hosts (P. absinthioides, A. tenuifolia, and A. scabra) loses aerial biomass during the cold season due to the harsher frosts. Plants in the genus Xanthium are always absent in winter because they are annual (Logarzo and Gandolfo 2005).
The cerambycid seems to have adapted its life cycle, by either losing or acquiring a diapause, to synchronize with the seasonality of the host plants. In our study, differences in host availability and insect life cycle traits, i.e., voltinism versus diapause (Table 7), result in a certain degree of gene flow restriction (Wood and Keese 1990; Glover et al. 1992). For instance, migrants from zones 2 and 3 are at a disadvantage in zone 1 since they are obligate diapausers that have only one generation per year (versus two generations of zone 1 individuals). Also, individuals from zone 3 (restricted to Pluchea spp.) that migrate to zone 2 may suffer fitness reductions due to the scarcity of suitable Pluchea hosts (Table 2, Fig. 2). In addition, preliminary results from mating choice tests showed a much higher mating success of males from P. absinthioides in zone 3 than males in zone 2 (87.5 vs. 12.5% mating success), and of males from Pluchea spp. than males of Xanthium cavanillesii (72.5 vs. 27.5%) (GAL, unpublished data), probably due to particular chemical traits of the genera Pluchea (Tooker et al. 2002; Moayeri et al. 2007) which affect reproductive performance. However, these results should be considered with caution, because captivity conditions may bias mating preferences (Hoffman and Turelli 1985).
Our study provides behavioral data suggesting that intraspecific geographic variation in host use has a genetic basis, and mtDNA data pointing to a recent geographic expansion coupled with subsequent genetic differentiation among geographic zones. Apparently, the ecological availability of the preferred host P. sagittalis triggered genetic differentiation involving host range and life cycle changes, determining gene flow restriction and resulting in geographic races, probably within an ecological speciation process.
No single genotype or host range can be uniformly successful in all environments (Falconer 1952). For instance, monophagy might be advantageous for A. versicolor in zone 1, where the preferred and putative ancestral host P. sagittalis is an abundant and predictable resource, while generalism or host shifts might be advantageous in zones 2 and 3, where P. sagittalis exhibits temporal and spatial unavailability or is absent. Our results echo those of other studies which show that host species range increases when the host species are less predictable, and narrows to monophagy when they are abundant and seasonally stable (Shapiro 1975, 1976; Chew 1977, Jaenicke 1978, Wiklund 1982). Host specialization may represent an adaptive advantage in environments with predictable host availability, whereas generalism or host shifts may provide an adaptive advantage due to the possibility of using alternative hosts in host-restricted environments.
References
Alleaume-Benharira M, Pen IR, Ronce O (2006) Geographical patterns of adaptation within a species’ range: interactions between drift and gene flow. J Evol Biol 19:203–215
Arnqvist G (1998) Comparative evidence for the evolution of genitalia by sexual selection. Nature 393:784–786
Bernays E, Graham M (1988) On the evolution of host specificity in phytophagous arthropods. Ecology 69:886–892
Bernays EA, Mikenberg OPJM (1997) Insect herbivores: different reasons for being a generalist. Ecology 78:1157–1169
Berner D, Blanckenhorn WU, Körner C (2005) Grasshoppers cope with low host plant quality by compensatory feeding and food selection: N limitation challanged. Oikos 111:525–533
Bigger DS, Fox LR (1997) High-density populations of diamondback moth have broader host-plant diets. Oecologia 112:179–186
Bremer K (1994) Branch support and tree stability. Cladistics 10:295–304
Burgos JJ (1963) Las heladas en Argentina. Colección científica de Instituto Nacional de Tecnología Agropecuaria (INTA), vol III, 1st edn. INTA, Buenos Aires
Burnham KP, Anderson DR (2002) Model selection and multimodel inference: a practical information-theoretical approach, 2nd edn. Springer, New York
Bush GL (1994) Sympatric speciation: new wine in old bottles. Trends Ecol Evol 9:285–288
Cabrera AL (1994) Enciclopedia Argentina de Agricultura y Jardinería, Tomo II, Fascículo 1. Regiones fitogeográficas argentinas. ACME, Buenos Aires
Chapman T, Arnqvist G, Bangham J, Rowe L (2003) Sexual conflict. Trends Ecol Evol 18:41–47
Chew FS (1977) Coevolution of Pierid butterflies and their cruciferous hostplants. II The distribution of eggs on potential foodplants. Evol 31:568–579
Chew FS, Courtney SP (1991) Plant apparency and evolutionary escape from insect herbivory. Am Nat 138:729–750
Choe J, Crespi BJ (1997) The evolution of mating systems in insects and arachnids. Cambridge University Press, Cambridge
Clement M, Posada D, Crandall KA (2000) TCS: a computer program to estimate gene genealogies. Mol Ecol 9:1657–1660
Craig TP, Itami JK, Abrahamson WG, Horner JD (1993) Behavioural evidence for host-race formation in Eurosta solidaginis. Evolution 47:1696–1710
Crawley MJ (2005) Statistics: an introduction using R. Wiley, Chichester
Crespi BJ, Sandoval CP (2000) Phylogenetic evidence for the evolution of ecological specialization in Timema walking-sticks. J Evol Biol 13:249–262
Cunningham JP, West SA, Wright DJ (1998) Learning in the nectar foraging behavior of Helicoverpa armigera. Ecol Entomol 23:363–369
Dethier VG (1954) Evolution of feeding preferences in phytophagous insects. Evolution 8:33–54
Dicke M (2000) Chemical ecology of host-plant selection by herbivorous arthropods: a multitrophic perspective. Biochem Syst Ecol 28:601–617
Ehrlich PR, Raven PH (1964) Butterflies and plants: a study in coevolution. Evolution 18:586–608
Erwin TL (1982) Tropical forests: their richness in Coleoptera and other arthropod species. Coleopt Bull 36:74–75
Excoffier L, Smouse PE, Quattro JM (1992) Analysis of molecular variance inferred from metric distance among DNA haplotypes: application of human mitochondrial DNA restriction date. Genetics 136:343–359
Excoffier L, Laval G, Schneider S (2005) Arlequin ver 3.1. An integrated software package for population genetics data analysis. Evol Inform Online 1:47–50
Falconer DS (1952) The problem of environment and selection. Am Nat 86:293–298
Felsenstein J (1985) Phylogenies and the comparative method. Am Nat 125:1–15
Fox CW, Czesak ME (2000) Evolutionary ecology of progeny size in arthropods. Annu Rev Entomol 45:341–369
Fraenkel GS (1959) Raison d’etre of secondary plant substances. Science 129:1466–1470
Freeland WJ, Janzen DH (1974) Strategies in herbivory by mammals: the role of plant secondary compounds. Am Nat 108:269–289
Fu YX (1997) Statistical tests of neutrality of mutations against population growth, hitchhiking and background selection. Genetics 147:915–925
Funk DJ, Bernays EA (2001) Geographic variation in host specificity reveals host range evolution in Uroleucon ambrosiae aphids. Ecology 82:726–739
Futuyma DJ (1983) Selective factors in the evolution of host choice by phytophagous insects. In: Ahmad S (ed) Herbivorous insects: host-seeking behavior and mechanism. Academic, New York
Futuyma DJ (2001) Ecological specialization and generalization. In: Fox CW, Roff DA, Fairbairn DJ (eds) Evolutionary ecology: concepts and case studies. Oxford University Press, London, pp 177–189
Futuyma DJ, Keese MC (1992) Evolution and coevolution of plants and phytophagous arthropods. In: Rosenthal GA, Berenbaum M (eds) Herbivores: their interactions with plant secondary metabolites. VII: Evolutionary and ecological processes. Academic, San Diego, pp 439–475
Futuyma DJ, Moreno G (1988) The evolution of ecological specialization. Annu Rev Ecol Syst 19:207–233
Futuyma DJ, Keese MC, Scheffer SJ (1993) Genetic constraints and the phylogeny of insect–plant associations: responses of Ophraella communa (Coleoptera: Crysomelidae) to host plants of its congeners. Evolution 47:888–905
Futuyma DJ, Keese MC, Funk DF (1995) Genetic constraints on macroevolution: the evolution of host affiliation in the leaf beetle genus Ophraella. Evolution 49:797–809
Gandolfo DE, Logarzo GA, Cordo HA (1997) Apagomerella versicolor (Coleoptera: Cerambycidae) candidato para el control biológico de Xanthium strumarium en USA. Estimación del daño de Xanthium cavanillesii en laboratorio. Rev Soc Entomol Argent 56:147–150
Glover TJ, Robbins PS, Eckenrode CJ, Roelofs WL (1992) Genetic control of voltinism characteristics in European corn borer races assessed with a marker gene. Arch Insect Biochem Physiol 20:107–117
Goloboff PA, Farris JS, Nixon KC (2008) TNT, a free program for phylogenetic analysis. Cladistics 24:1–13
Groman JD, Pellmyr O (2000) Rapid evolution and specialization following host colonization in a yucca moth. J Evol Biol 13:223–236
Harvey JA, Kadash K, Strand MR (2000) Differences in larval feeding behavior correlate with altered developmental stages in two parasitic wasps: implications for the size–fitness hypothesis. Oikos 88:621–629
Heimpel GE, Rosenheim JA, Mangel M (1996) Egg limitation, host quality, and dynamic behavior by a parasitoid in the field. Ecology 77:2410–2420
Hoffman AA, Turelli MI (1985) Distribution of Drosophila melanogaster on alternative resources: effects of experience and starvation. Am Nat 126:662–679
Hudson RR (1990) Gene genealogies and the coalescent process. Oxf Surv Evol Biol 7:1–44
Hudson RR, Boos DD, Kaplan NL (1992) A statistical test for detecting geographic subdivision. Mol Biol Evol 9:138–151
Itami JK, Craig TP, Horner JD (1998) Factors affecting gene flow between the host races of Eurosta solidaginis. In: Mopper S, Strauss S (eds) Genetic structure and local adaptation in natural insect populations: effects of ecology, life history and behavior. Chapman and Hall, New York, pp 375–407
Jaenicke J (1978) On optimal oviposition behavior in phytophagous insects. Theor Popul Biol 14:350–356
Jaenike J (1990) Host specialization in phytophagous insects. Annu Rev Ecol Syst 21:243–273
Janz N, Nyblom K, Nylin S (2001) Evolutionary dynamics of host-plant specialization: a case study of the tribe Nymphalini. Evolution 55:783–796
Janz N, Nylin S, Wahlberg N (2006) Diversity begets diversity: host expansions and the diversification of plant-feeding insects. BMC Evol Biol 6:4. doi:10.1186/1471-2148-6-4
Jeffries MJ, Lawton JH (1984) Enemy-free space and the structure of ecological communities. Biol J Linn Soc 23:269–286
Johns R, Quirin DT, Lapointe R, Lucarotti CJ (2009) Foliage-age mixing within balsam fir increases the fitness of a generalist caterpillar. Ecol Entomol 34:624–631
Karban R (1989) Fine-scale adaptation of herbivorous thrips to individual host plants. Nature 340:60–61
Kawecki TJ, Ebert D (2004) Conceptual issues in local adaptation. Ecol Lett 7:1225–1241
Kelley ST, Farrell BD (1998) Is specialization a dead end? The phylogeny of host use in Dendroctonus bark beetles (Scolytidae). Evolution 52:1731–1743
Lande R (1981) Models of speciation by sexual selection on polygenic characters. Pro Natl Acad Sci USA 78:3721–3725
Librado P, Rozas J (2009) DnaSP v5: A software for comprehensive analysis of DNA polymorphism data. Bioinformatics 25:1451–1452
Logarzo GA, Gandolfo DE (2005) Análisis de voltinismo y la diapausa en poblaciones de Apagomerella versicolor (Coleoptera: Cerambycidae) en el gradiente latitudinal de su distribución en la Argentina. Rev Soc Entomol Argent 64:143–146
Logarzo GA, Gandolfo DE, Cordo HA (2002) Biology of Apagomerella versicolor (Coleoptera: Cerambycidae) in Argentina, a candidate for biological control of cocklebur (Xanthium spp.) in US. Biol Control 25:22–29
Lucas PW, Turner IM, Dominy NJ, Yamashita N (2000) Mechanical defenses to herbivory. Ann Bot 86:913–920
Matsubayashi KW, Ohshima I, Nosil P (2010) Ecological speciation in phytophagous insects. Entomol Exp Appl 134:1–27
Mitter CB, Farrell B, Wiegmann B (1988) The phylogenetic study of adaptive zones has phytophagy promoted insect diversification? Am Nat 132:107–128
Moayeri HRS, Ashouri A, Brodsgaard HF, Enkegaard A (2007) Males of the predatory mirid bug Macrolophus caliginosus exploit plant volatiles induced by conspecifics as a sexual synomone. Entomol Exp Appl 123:49–55
Mopper S (1996) Adaptive genetic structure in phytophagous insect populations. Trends Ecol Evol 11:235–238
Mopper S, Micheal B, Simberlof D, Stiling P (1995) Local adaptation and agents of selection in a mobile insect. Evolution 49:810–815
Morrone JJ (2006) Biogeographic areas and transition zones of Latin America and the Caribbean islands based on panbiogeographic and cladistic analyses of the entomofauna. Annu Rev Entomol 51:467–494
Neu CW, Byers CB, Peek JM (1974) A technique for analysis of utilization-availability data. J Wildl Manag 38:541–545
Nitao JK, Ayres MP, Lederhouse RC, Scriber JM (1991) Larval adaptation to lauraceous hosts: geographic divergence in the spicebush swallowtail butterfly. Ecology 72:1428–1435
Panchal M (2007) The automation of nested clade phylogeographic analysis. Bioinformatics 23:509–510
Papaj DR (1986) Conditioning of leaf-shape discrimination by chemical cues in the butterfly, Battus philenor. Anim Behav 34:1281–1288
Piccinali RV, Marcet PL, Noireau F, Kitron U, Gürtler RE, Dotson EM (2009) Molecular population genetics and phylogeography of the Chagas disease vector Triatoma infestans in South America. J Med Entomol 46:796–809
Posada D, Crandall KA, Templeton AR (2000) GeoDis: a program for the cladistic nested analysis of the geographical distribution of genetic haplotypes. Mol Ecol 9:487–488
Price PW, Bouton CE, Gross P, Mcpheron BA, Thompson JN, Weis AE (1980) Interactions among three trophic levels: influence of plants on interactions between insect herbivores and natural enemies. Annu Rev Ecol Syst 11:41–65
Prokopy RJ, Diehl SR, Cooley SS (1988) Behavioral evidence for host races in Rhagoletis pomonella flies. Oecologia 76:138–147
Ramos-Onsins SE, Rozas J (2002) Statistical properties of new neutrality tests against population growth. Mol Biol Evol 19:2092–2100
Schluter D (2000) The ecology of adaptive radiation. Oxford University Press, New York
Schluter D (2009) Evidence for ecological speciation and its alternative. Science 323:737–741
Scriber JM, Slansky F (1981) The nutritional ecology of immature insects. Annu Rev Entomol 26:183–211
Shapiro AM (1975) Ecological and behavioral aspects of coexistence in six crucifer-feeding pierid butterflies in the central Sierra Nevada. Am Midl Nat 93:424–433
Shapiro AM (1976) The role of watercress (Nasturtium officinale) as a host of native and introduced pierid butterflies in California. J Res on the Lepid 14:158–168
Singer MS (2001) Determinants of polyphagy by a woolly bear caterpillar: a test of the physiological efficiency hypothesis. Oikos 93:194–204
Singer MC, Ng D, Vasco D, Thomas CD (1992) Rapidly evolving associations among oviposition preferences fail to constrain evolution of insect diet. Am Nat 139:9–20
Singer MC, Thomas CD, Singer M (1993) Rapid human-induced evolution of insect–host associations. Nature 366:681–683
Smith DC (1988) Heritable divergence of Rhagoletis pomonella host races by seasonal asynchrony. Nature 336:66–67
Stiling P, Rossi AM (1998) Deme formation in a dispersive gall-forming midge. In: Mopper S, Strauss SY (eds) Genetic structure and local adaptation in natural insect populations. Chapman and Hall, New York, pp 22–36
Tajima F (1989) Statistical methods to test for nucleotide mutation hypothesis by DNA polymorphism. Genetics 123:585–595
Templeton AR, Crandall KA, Sing CF (1992) A cladistic analysis of phenotypic associations with haplotypes inferred from restriction endonuclease mapping of DNA sequence data. III: Cladogram estimation. Genetics 132:619–633
Thompson JN (1993) Preference hierarchies and the origin of geographic specialization in host use in swallowtail butterflies. Evolution 47:1585–1594
Thompson JN (1998) The evolution of diet breadth: monophagy and polyphagy in swallowtail butterflies. J Evol Biol 11:563–578
Tooker JF, Koenig WA, Hanks LM (2002) Altered host plant volatiles are proxies for sex pheromones in the gall wasp Antistrophus rufus. Proc Natl Acad Sci USA 99:15486–15491 (see http://www.pnas.org/cgi/reprint/99/24/15486)
Van Zandt P, Mopper S (1998) A meta-analysis of adaptive deme formation in phytophagous insect populations. Am Nat 152:597–606
Via S (1984) The quantitative genetics of polyphagy in an insect herbivore. II. Genetic correlations in larval performance within and among host plants. Evolution 38:896–905
Via S, Lande R (1985) Genotype–environment interaction and the evolution of phenotypic plasticity. Evolution 39:505–522
Westoby M (1977) What are the biological bases of varied diets? Am Nat 112:627–631
Wiklund C (1982) Generalist versus specialist utilization of host plants among butterflies. In: Wisser JH, Minks AK (eds) Insect–plant relationships. PUDOC, Wageningen, pp 181–192
Williams GC (1966) Adaptation and natural selection: a critique of some current evolutionary thought. Princeton University Press, Princeton
Wood TK, Keese MC (1990) Host plant induced assortative mating in Enchenopa treehoppers. Evolution 44:619–628
Zuccon A, Zuccon D (2010) MrEnt v.2.1. http://www.mrent.org
Acknowledgments
We thank Laura Varone, Florencia Palotini, Vanina Varni, and Javier Jara for their excellent technical assistance. We thank Juan Briano, Arabella Bugliani, Laura Yadhjian, Jesse de León, and Luis Calcaterra for reading and improving the manuscript. Appreciation is also extended to Daniel Vazquez, Anibal Carbajo, Alejandro Sosa, and Hugo Cordo for comments on earlier drafts. Finally, we thank the anonymous reviewers and the editor for thoughtful comments that greatly improved the manuscript. EH and RVP are members of Carrera de Investigador Científico (Scientific Research Career) (CONICET, Argentina). All experiments performed in this article comply with the current laws of Argentina.
Author information
Authors and Affiliations
Corresponding author
Additional information
Communicated by Roland Brandl.
Rights and permissions
About this article
Cite this article
Logarzo, G.A., Casalinuovo, M.A., Piccinali, R.V. et al. Geographic host use variability and host range evolutionary dynamics in the phytophagous insect Apagomerella versicolor (Cerambycidae). Oecologia 165, 387–402 (2011). https://doi.org/10.1007/s00442-010-1782-2
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00442-010-1782-2