
Abstract
Leaf traits related to the performance of invasive alien species can influence nutrient cycling through litter decomposition. However, there is no consensus yet about whether there are consistent differences in functional leaf traits between invasive and native species that also manifest themselves through their “after life” effects on litter decomposition. When addressing this question it is important to avoid confounding effects of other plant traits related to early phylogenetic divergences and to understand the mechanism underlying the observed results to predict which invasive species will exert larger effects on nutrient cycling. We compared initial leaf litter traits, and their effect on decomposability as tested in standardized incubations, in 19 invasive-native pairs of co-familial species from Spain. They included 12 woody and seven herbaceous alien species representative of the Spanish invasive flora. The predictive power of leaf litter decomposition rates followed the order: growth form > family > status (invasive vs. native) > leaf type. Within species pairs litter decomposition tended to be slower and more dependent on N and P in invaders than in natives. This difference was likely driven by the higher lignin content of invader leaves. Although our study has the limitation of not representing the natural conditions from each invaded community, it suggests a potential slowing down of the nutrient cycle at ecosystem scale upon invasion.
Similar content being viewed by others

Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
Litter decomposition is a key process in nutrient cycling (Swift et al. 1979; Hobbie 1992; Sayer 2006). Rates of litter decomposition can greatly influence nutrient availability and thereby other ecosystem processes (Vitousek and Howarth 1991). At the community level, decomposition rates are controlled by species or functional type composition (Hättenschwiler and Vitousek 2000; Garnier et al. 2004; Quested et al. 2005). Therefore, the introduction or invasion of alien species into new ecosystems can cause profound changes in community structure and function (Mack et al. 2000; Kolar and Lodge 2001), including changes in litter decomposition and nutrient cycling (Allison and Vitousek 2004; Ashton et al. 2005). To determine the direction of these changes, several studies have compared litter decomposition between invasive alien species and natives. However, results differ greatly between studies, taxa, and regions (Ehrenfeld 2003) and no generalization (a frequent objective in the study of biological invasion) has arisen so far. Faster decomposition of litter from invasive alien species compared to native litter has been reported when invaders were N2-fixers and native species were not (Liao et al. 2008), when nutrient concentration in the exotic litter was higher than in the native one (Allison and Vitousek 2004) and when the specific leaf area (SLA) of invaders was higher than that of native species (Castro-Díez et al. 2009; Kueffer et al. 2008). By contrast, slower decomposition of invasive alien species’ leaf litter was found when it had a higher polyphenolic content (Knight et al. 2007), higher lignin content or higher C/N ratio than native litter (Drenovsky and Batten 2007).
Differences between studies may be partly due to over-representation of particular regions (e.g. Hawaii islands; Allison and Vitousek 2004) or functional types (e.g. at least 1/3 of the articles involve N2-fixers; Levine et al. 2003), and they have generally included only a few species pairs, which have not accounted for phylogenetic and life history differences. Comparing large numbers of invasive and native species by means of phylogenetic independent contrasts is necessary to unveil general trends whilst avoiding confounding effects of phylogeny.
As important as avoiding confounding effects, is to understand the mechanisms underlying the direction of the change upon invasion. This will potentially help to predict which invasive alien species are likely to exert larger effects on nutrient cycling and which communities are likely to suffer larger impacts (Levine et al. 2003). Invasive species possessing leaf litter traits similar to natives will be functionally redundant in the nutrient cycling mechanisms and no changes may be expected. By contrast, invasive species with leaf litter traits largely different from those of natives, may impact the nutrient cycling. Thus, the direction and magnitude of the change will depend on the trait difference between these groups (Dukes and Mooney 2004; Strauss et al. 2006). Under Mediterranean soil conditions, where the availability of N and P is low (Gallardo and Parama 2007), we may predict that leaf litter from invasive alien species with traits that retard decomposition will inhibit and slow down nutrient cycling, while those promoting decomposition will have no effect (because of microorganism immobilization in cases of low nutrient soil concentration) or will slightly speed up nutrient cycling (Knorr et al. 2005).
By using this less frequently employed approach, we have the additional opportunity to link ecosystem impacts to functional strategies of invasive species, since many leaf litter traits are directly related to plant performance strategies such us competitive ability (e.g. variation in N content), defence against herbivores (e.g. variation in polyphenolic compounds) or resource use efficiency (e.g. variation in lignin content). Thus, while many studies have tried to link species traits with invasiveness, the importance of studying these traits is more related to differences in ecosystem impacts than in invasiveness.
The objective of the present study was to compare leaf litter decomposability between invasive alien and native species from Mediterranean ecosystems in Spain by means of an experimental design using phylogenetically independent contrasts (Armstrong and Westoby 1993). We addressed the following questions:
-
1.
Do leaf litter properties of invasive alien species differ consistently from those of native ones?
-
2.
Does leaf litter of invasive alien species decompose faster than that of native species?
-
3.
Do initial leaf litter traits influence the decomposition process in the same way in invasive and native species?
To address these questions, a set of 19 invasive-native co-familial pairs were selected and subjected to an experimental standardized screening for leaf litter decomposability together with physical–chemical analyses.
Materials and methods
Species selection
Our species set comprised introduced species that are clearly invasive (Pyšek et al. 2004), local dominants in some native ecosystems (Valéry et al. 2008) and with potential impact in the invaded ecosystems (Richardson et al. 2000). Species compiled by The Atlas of the Invasive Alien Species in Spain (Sanz Elorza et al. 2004) matched our criteria of invasive species definition. Thus, it was used to select the invasive alien species and the localities where the senesced leaf material was collected, ranging from central to eastern Spain (Table 1). The final selection of invasive species was done on the basis of leaf litter availability and aimed to represent of a broad diversity of growth forms (annual and biannual herbs, lianas, shrubs, and trees), invaded habitats (woodlands, shrublands, grasslands, and riparian areas), and different life histories. Nomenclature followed the Iberian Flora (Castroviejo 1986–2008). Each of the selected species was paired with one closely related native species based on two criteria: (1) within each pair, the native species had to belong to the same family as the invasive species; and (2) the two species had to belong to the same growth form (i.e. herbaceous vs. woody species).
The selection of phylogenetically independent invasive-native pairs for methodological reasons may induce biases in the ecological meaning of our results. To minimize this problem, we conducted a screening to select native species with a common occurrence throughout Spain and especially in the eastern part, which hosts the highest diversity and abundance of invasive alien species (Gasso et al. 2009). Native species with a small distribution range, and those which are rare or endangered were excluded. Additionally, we discarded those native species occurring in habitats different from those where alien species invade or with occurrence in successional community stages different from natives. The extended herbarium database at the Complutense University of Madrid was used to select, within pairs, native species recorded in the same localities or next to where the alien species pair invade. This procedure has permitted us to retain the potential ecological meaning of our results in the whole range of the invasive species, except for the case of Ailanthus altissima vs. Cneorum tricoccon (Table 1), which only has ecological sense in the Balearic Islands and north of Cataluña.
Collection of leaf litter
Recently senesced, undecomposed and herbivore-damage-free leaf litters of invasive alien species and their native relatives were collected following the criteria of Cornelissen (1996). Leaf litter from deciduous woody species was collected throughout the autumn 2005, while leaf litter of evergreen woody and herbaceous species was collected in the spring 2006, according to their natural leaf senescence period. Ten mature individuals for each species of the same population at full-sun exposure (in the case of trees) were preferentially chosen for senesced leaf collection. Large variation in the proportion of petioles to lamina was found between species (6.7 ± 5.3%), but petioles from all species were discarded to assess the decomposition ability of leaf litter, since the variability within pairs was low (1.2 ± 0.4%). Approximately 40 g of senesced leaves per species was obtained after pooling the leaves from all individuals. They were transported to the laboratory, air-dried at 20–25°C and stored in open paper bags until further analysis.
Litter decomposition experiment set up
For each species, four subsamples of 1.0 ± 0.1 g air-dried senesced leaves were used to calculate their initial water content by weighing them again after 3 days in the oven at 60°C. Sixteen samples of air-dried leaves were extracted from each species pool, weighed (1.0 ± 0.1 g) and corrected for water content. Then each of these samples was sealed into a 1-mm-mesh nylon bag to allow free movement of macro- and meso-decomposers into the bags (Cornelissen 1996). Large leaf size variation between species lead to variation in bag size in order to spread out whole leaves in one single layer, since surface access to soil decomposers is a determinant of decomposition rate (Cornelissen 1996). Litter bags were buried randomly in a purpose-built litter decomposition bed (7 m × 3.5 m) at Alcalá University Botanical Garden (Madrid province, Central Spain). This bed consisted of a thoroughly mixed 5-cm litter layer collected from a nearby mature Quercus spp. forest. We selected this material trying to create an incubation matrix of natural non-invaded forest communities representative of Spain with close-to-natural microbial and soil fauna activity. Additionally, we covered the bed with a single layer of senesced Platanus hispanica leaves. The decomposition bed was displayed below a shade frame covered by a neutral shade mesh, which removed 30% of the solar radiation, in order to avoid rapid water loss, especially in summer. On 23 December 2005, the litter bags of the autumn-deciduous invasive-native pairs (pair numbers 1, 4, 8 and 18; Table 1) were placed in the litter decomposition bed at 2-cm depth in the matrix, while the rest of the 15 pairs (where at least one of the species pair shed its leaves in spring) were placed there on 22 June 2006. Two harvests of eight litterbags per species were collected, the first after 147 and 143 days of incubation (for the December and June pairs, respectively), and the final after 289 and 231 days. The first harvest date was chosen by expert criteria to select a steady-state of mean litter decomposition around 50% of mass loss taking into consideration all species, while the final harvest date was chosen when the species with faster litter decomposition (Colutea arborescens, Anagyris foetida) were close to full decomposition. Eight additional litterbags of Acer campestre, Lonicera japonica and Ulmus pumila were placed in each of the two periods of our experiment to assess the effect of starting date on the decomposition rate. At each harvest, the material remaining in each litterbag was carefully brushed clean, dried at 60°C during 3 days, and weighed to calculate the percentage of litter mass loss. For contamination by dust particles see below.
At the beginning of the experiment, three 2.5 × 2.5-cm anion and two 2.5 × 2.5-cm cation exchange resin membranes (types I-100 and I-200; Electropure excellion, Laguna Hills, Calif.) fixed to a plastic label were introduced into the soil at three equidistant points of the litter bed at 4-cm depth, and left for 15 days to evaluate inorganic N and P availability (Abrams and Jarrell 1992; Subler et al. 1995). Resin membranes had been previously conditioned in the lab by immersing them in demineralised water at 82–90°C for 48 h. Ions were extracted from the resins using 20–25 ml KCl and ammonia, nitrate and phosphate contents were assessed following Sims et al. (1995). Finally, local air temperature, available photon flux density (PPFD) and soil moisture within the first 15-cm soil depth were registered every 15 min during the litter decomposition experiment with a data logger (HOBO model H08-006-04; Onset, Pocasset, Mass.) and measured by external sensors [LI-COR, Lincoln, Neb.; ThetaProbe sensors (Delta-T Devices, Cambridge, UK)]. The litter decomposition bed was occasionally watered during dry periods, when soil moisture dropped below 60% gravimetric water content (GWC).
Measurements
The initial area of the senesced leaves was measured in eight subsamples of eight leaves each per species (leaf area meter device; Delta-T ), which were then oven-dried at 60°C for 3 days and weighed to calculate their SLA (= leaf area/leaf dry mass). The remaining leaf litter, after removing samples for litter decomposition and SLA assessment, was ground in a single pool per species, using a Cullati mill (1-mm particle size). Chemical analyses to determine litter leaf traits were conducted at the laboratory of the Department of Systems Ecology, Vrije Universiteit, Amsterdam, The Netherlands. Total initial concentrations of organic C, N, P, Ca, K, Mg and pH were calculated from a set of four ground subsamples per species, while initial lignin and cellulose contents were assessed in one ground sample per species. C and N were also measured in four ground samples of the final harvest. Finally, in four subsamples per species, ash content was measured (mass loss on ignition, 575°C) to correct litter sample dry mass for possible sand or clay contamination. Since some plants can contain significant amounts of silica and other minerals in their tissues (Epstein 1994), we only corrected for ash fractions greater than 2–3% for species where we observed or suspected contamination by dust.
Total C and N were determined by dry combustion with a NA1500 series II elemental analyser (Carlo Erba, Rodana, Italy). After digestion (of ground leaf material) in a 1:4 mixture of 37% (v/v) HCl and 65% (v/v) HNO3 (Sneller et al. 1999) P was measured colorimetrically, Ca and Mg were measured with AAS and K with AES (both: Perkin Elmer 1100, Waltham, Mass.). pH was measured using a narrow (5 mm diameter) SenTix Mic electrode connected to an Inolab Level 2 pH meter (both: WTW, Weilheim, Germany). Initial litter lignin and cellulose concentration was determined following Poorter and Villar (1997): in short, after several extraction steps to ensure that only cellulose and lignin made up the composition of the residue of the sample, the C and N concentrations of this residue were used to calculate the lignin concentration, based upon the difference in C content between cellulose and lignin.
Statistics
We applied Statistica 6.1 (StatSoft) and R package (library “stats”; R Development Core Team 2009) for all analyses below. Percentage of litter mass loss at the first and final harvests (with initial mass as the reference for both harvests) was arcsine square-root(×/100) transformed to approach normal distributions and homogeneity of variance (Quested et al. 2003). Given that some families were over-represented in our dataset (e.g. Fabaceae; Table 1) and that the 19 species pairs were not randomly selected, a Mantel test was conducted to test whether differences in litter decomposition within pairs were influenced by pair selection. The phylogenetic distance from one species to another between all the species pairs was calculated through the first common ancestor to both species using the plant phylogenetic supertree described by Soltis et al. (2000) and modifications by Bremer et al. (2003).
Pairwise Student t-tests compared initial physical–chemical leaf litter traits and percentage of litter mass loss across all pairs of invasive alien species and the native co-familial counterpart, at the first and final harvests, respectively. The effect of the start date of incubation (autumn or spring) on the percentage of litter mass loss was assessed for the three control species (A. campestre, L. japonica and U. pumila) by a two-way ANOVA using time and species as main independent factors.
In order to assess the relative importance of different predictors for the litter mass loss, a four-way ANOVA was conducted taking as the main independent variables, Growth form, Family nested in Growth form—because none of the families comprised both woody and herbaceous species—Leaf type (deciduous or evergreen) and Plant status (invasive or native). Additionally, multiple-regression models were conducted to evaluate which senesced leaf traits better predicted the percentage of litter mass loss in each species group (invasive alien and native species) at each harvest. Pearson correlations were conducted to check for co-linearity between predictors. Predictors which were not correlated or were poorly correlated were chosen (r < 0.10), while among those highly correlated (r > 0.70) only one predictor with a high ecological meaning was selected (Fowler and Cohen 1992). Therefore, initial lignin concentration was discarded because it was highly correlated with the initial C concentration in both species sets (invasive species r > 0.80 and native species r > 0.75). Different models were fitted selecting different sets of predictive variables by the approach based on information theory (Akaike information criteria; AIC; Burnham and Anderson 2002). AIC was calculated for each model and used to choose the best fitted. A value of difference in AIC (ΔAIC) higher than 10 represents a poorly fitted model compared to the best model, whereas a value of less than 2 indicates that both models are equivalent (Burnham and Anderson 2002).
Results
Litter bed characteristics
The most abundant form of available mineral N in the litter decomposition bed was NO3 −–N (1.82 μg m−2 day−1), which was two times more than NH4 +–N (0.86 μg m−2 day−1). The availability of PO4 −–P was 0.72 μg m−2 day−1, and thus the N/P ratio of the available pool was 1.9. Mean (±SE) air temperature, soil GWC and daily PPFD during the first incubation period for deciduous autumn pairs (see “Materials and methods”) were 10.21 (±7.06)°C, 68.74 (±3.46)% and 24 mol m−2 day−1, respectively, whereas for the final incubation period with the rest of the pairs the, respective, means were 16.84 (±9.77)°C, 65.12 (±5.73)% and 32 mol m−2 day−1.
Leaf litter traits, decomposition rates and decomposition predictors
SLA did not differ between invasive alien and native species (Table 2). Initial Ca and lignin concentrations were significantly higher in invasive species. Total C concentration was marginally higher in invasive species. Cumulative mass loss at the final harvest was significantly higher in invasive species of a pair than in their co-familial native species, whereas a similar trend was not significant at the first harvest (Table 2). Mean values of all measured traits for each species are shown in the Electronic supplementary material (Table S1). Mantel test R (% mass loss − phylogeny = 1.34; P = 0.529), showed that differences in % litter mass loss between invasive and native species were not influenced by the phylogenetic structure of the invasive-native pair selection. In addition, two-way ANOVA showed that the effects of the start date of incubation were comparable because significant differences were only found for species (F 2,90 = 41.68, P < 0.001), but not for time period (F 1,90 = 0.09, P = 0.769) or the time period by species interaction (F 2,90 = 1.66, P = 0.206).
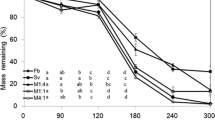
Four-way ANOVA showed that growth form was the most important categorical factor explaining the variation of final litter mass loss across the whole data set, mass loss being faster in herbaceous than in woody species. Family was the second best predictor, followed by plant status (invasive-native), native species decompose faster than invasive species, and finally leaf type, deciduous decomposing faster than evergreen species. No significant effect was found in the interaction between plant status and the other categorical factors (Fig. 1).

Four-way ANOVA assigning the relative importance of Family, Growth form (woody/herbaceous), Leaf type (evergreen deciduous) and Status (invasive/native) on percentage mass of leaf litter, based on the 19 invasive-native co-familial pairs (see Table 1). Mass loss % was arcsine [square-root(×/100)] transformed prior to analysis. Family, as a predictor, was nested in Growth form because there was no Family with both woody and herbaceous species. Explained/residual variance corresponds with the F-values of the ANOVA
Leaf litter decomposition processes
Total initial concentrations of C, N, P, cations (sum of Ca + K + Mg), pH, cellulose and SLA were the independent predictors selected to assess the relative importance of leaf litter traits in litter decomposition of both harvests and for the invasive and native species groups separately (Table 3). Initial lignin was excluded due to its high correlation with initial C concentration (see “Materials and methods”). Multiple-regression models revealed that the predictors were different between invasive and native species and between harvests (Table 3). N was related positively to litter decomposition in all cases and its relative importance increased at the final harvest for invasive species. P was the most important positive predictor of % litter mass loss, but only for invasive species at the final harvest. C and thus lignin were negatively related to litter mass loss in native species across the whole incubation period, while only in the first harvest in invasive species (Table 3). pH was a positive predictor of native litter decomposition rate at the final harvest and of invasive species only at the first harvest. A positive relation between decomposition rate and SLA only emerged at final harvest for both groups of species. Finally, cellulose was slightly negatively related to decomposition rate in native species at both harvests (Table 3).
The goodness-of-fit of the models (r 2) was higher for native than for invasive species. Regression models generally predicted litter decomposition rate better at the final than at the first harvest for invasive species (Table 3).
Discussion
Leaf litter characteristics and mass loss rates
Leaf litter of alien species invading Mediterranean ecosystems in Spain tended to decompose slightly more slowly than that of the co-familial native species. This could be due to a large initial lignin content, which was significantly higher in invasive alien species across invasive-native species pairs. This well-studied relationship (Swift et al. 1979) was recently confirmed within multiple sites worldwide (Cornwell et al. 2008). Invasive alien species combine higher lignin with higher Ca initial content (Table 2). Ca plays an important role in lignin synthesis and a deficit of this macroelement in soils limits lignification of plant tissues (Lautner et al. 2007). Indeed, the soils of the locations where leaf litter was collected (Table 1) are characterized by a high availability of Ca (Rubio and Escudero 2005), which apparently does not limit the tissue lignification of invasive alien species. Although plant species trade off plant growth for tissue lignification (Villar et al. 2006), the elevated C assimilation capacity for growth commonly described in invasive species (Pyšek and Richardson 2007) and the higher levels of litter lignin found in our study, may reflect a C availability sufficient to maximize both strategies. This overall C strategy apparently improves the growth rate and protection of leaf tissues which leads to a higher nutrient use efficiency, another strategy also reported in invasive alien species (Pyšek and Richardson 2007).
Our results do not match those from previous studies comparing leaf litter properties of invasive and native species, as these studies showed that, in general, invasive alien species tended to exhibit higher SLA and higher nutrient concentrations [see Ehrenfeld (2003) for a detailed description]. However, these differences seemed to be biased by the selection of species and regions (mostly N2-fixing invasive species in oligotrophic soils; Levine et al. 2003; Yelenik et al. 2007).
Predictors of litter decomposition rate
N content in litter was related to litter decomposition in all species groups and in both harvests (Table 3). This agrees with the well-known fact that high N content in litter and in soil promotes decomposition (Ehrenfeld 2003; Cornwell et al. 2008). However, its relative importance was higher in invasive alien species in the final harvest (Table 3), which may be attributed to their higher lignin content. Lignin, which forms complex associations with litter proteins, inhibits the action of decomposing organisms and delays the decomposition processes during the first months (Gallardo and Merino 1993). Thus, N, unlike lignin, becomes an important predictor later on, when soil organism activity overcomes the barrier of the lignin complex and starts to release N from litter proteins (Gallardo and Merino 1992). In addition, P content in leaf litter exhibited a great positive influence on the decomposition of invasive alien litter, but not on native litter (Table 3). Both results are in agreement with Bubb et al. (1998) and Xuluc-Tolosa et al. (2003), who demonstrated that litter with a high lignin content needs a high N content in leaves and soil for its decomposition, and that this process becomes P dependent. Consequently it may become important as a limiting factor for the decomposition of the lignin-rich litter of invasive alien species when lignin degradation by micro-organisms takes off. Perhaps the lower lignin content of the litter of the native species makes them less dependent on P and on N (Table 3; Knorr et al. 2005).
Overall, leaf traits predicted litter decomposition rate better for native than for exotic species, especially at the first harvest. This suggests that other unquantified litter traits of exotic species might initially exert a tighter control on the decomposition process than the measured traits. For instance, high levels of relatively mobile polyphenolic compounds have been commonly reported in invasive species (Ehrenfeld 2003; Knight et al. 2007). These C-based defences may be useful under the full-sun habitat conditions of the early community succession stages, where aliens mainly invade because they provide protection from UV light, and defence against generalist herbivores, but not against the specialist ones (see references in Marko et al. 2008). Such an ecological strategy of invasive species remains in the senesced leaves, influencing the initial processes of litter decomposition (Hättenschwiler and Vitousek 2000).
Consequences of invasions for nutrient cycling
Lignin is an immobile C-based substance that influences nutrient cycles at the ecosystem scale by slowing down litter decomposition (Swift et al. 1979; Cornwell et al. 2008). Contrary to most other studies on litter decomposition rates by aliens versus natives, our results have demonstrated that invasive alien species produce less decomposable leaf litter compared to co-familial native Mediterranean plants, probably due to their higher lignin content. Our results, therefore, suggest that the soil nutrient turnover, and thereby nutrient availability, will not be enhanced upon invasion in Spanish Mediterranean ecosystems; actually they might even be slightly inhibited. However, our experiment has the limitation of not representing natural soil and litter layer conditions in the habitats where invasive species are becoming abundant. In addition, our decomposition bed litter matrix was composed of species (Quercus spp.) that generate low decomposition rates (Gallardo and Merino 1993) and possibly select for organisms adapted to the decomposition of lignin-rich litter but reduce soil fauna and microbial diversity (Gallardo and Merino 1992). Thus, changes in decomposer community, soil conditions, or both caused by invasion processes may feedback or moderate the decomposition patterns reported here. For instance, although litters from invasive Acacia species are hard to decompose (see “Electronic supplementary material”, Table S1; Yelenik et al. 2007) the N2-fixing strategy of these species highly augments N soil availability and microbial soil activity, accelerating litter decomposition upon invasion (Yelenik et al. 2007). High soil N and P availability by natural or human-mediated actions may lead to the same results (Knorr et al. 2005). However, Spanish Mediterranean soils are in general characterized by their low N and P content (Gallardo and Parama 2007) and their low microbial activity and biomass (Gallardo and Merino 1992). Accordingly, a progressive increment of invasive plant species characterized by their lignin-rich litter and without a significant increment in available N and P in soil, would cause a slowing down of nutrient cycling by reducing the litter quality and the abundance and diversity of decomposer communities (Gallardo and Merino 1992; Chigineva et al. 2009). In this sense, Cotrufo et al. (1998) demonstrated that litter of impoverished quality due to an increment in lignin content and lignin:N ratio was avoided by soil fauna in a food choice experiment.
Our results also suggest that under the scenario of a progressive displacement of native species by invasive ones, altering the spectra of growth forms in an ecosystem, invokes the highest relative impact on the litter decomposition rate (the relative impact depending on whether the native community is completely displaced or invasive and native species coexist; Fig. 1). Consequently, changes in community structure from herbaceous to less decomposable invasive woody species, which have been reported for different Mediterranean-type ecosystems worldwide (Di Tomaso 1998; Bousquet-Melou et al. 2005) could augment this negative feedback on litter decomposition. Our data also highlight the importance of Family and, to a lesser extent, Leaf type on the litter decomposition rate (Fig. 1). The impoverishment of taxonomic diversity in native woody communities due to the introduction in forestry of exotic evergreen trees, which belong to a few selected taxonomic groups (e.g. Pinus, Pinaceae; Eucalyptus, Myrtaceae; see “Electronic supplementary material”, Table S1 for mean mass loss values), could also augment this negative feedback. There may be worse consequences for nutrient cycling of the invasive plant species on establishment when both effects occur at the same time, as has been reported in the Cape region of South Africa (Manders and Richardson 1992).
In summary, invasive species have the potential to slow down nutrient cycling in Spanish Mediterranean ecosystems through slower litter decomposition. The high lignin content that drives this change is a reflection of the features displayed by alien species which allow them to become invasive. The degree of the impact will be highest when invasive species have a higher lignin content than natives, which include a few genera of woody evergreen alien species.
References
Abrams MM, Jarrell WM (1992) Bioavailability index for phosphorous using ion-exchange resin impregnated membranes. Soil Sci Soc Am J 56:1532–1537
Allison SD, Vitousek PM (2004) Rapid nutrient cycling in leaf litter from invasive plants in Hawaii. Oecologia 141:612–619
Armstrong DP, Westoby M (1993) Seedlings from large seeds tolerate defoliation better—a test using phylogenetically independent contrast. Ecology 74:1092–1100
Ashton IW, Hyatt LA, Howe KM, Gurevitch J, Lerdau MT (2005) Invasive species accelerate decomposition and litter nitrogen loss in a mixed deciduous forest. Ecol Appl 15:1263–1272
Bousquet-Melou A, Louis S, Robles C, Greff S, Dupouyet S, Fernandez C (2005) Allelopathic potential of Medicago arborea, a Mediterranean invasive shrub. Chemoecology 15:193–198
Bremer B, et al (2003) An update of the Angiosperm Phylogeny Group classification for the orders and families of flowering plants: APG II. Bot J Linn Soc 141:399–436
Bubb KA, Xu ZH, Simpson JA, Saffigna PG (1998) Some nutrient dynamics associated with litterfall and litter decomposition in hoop pine plantations of southeast Queensland, Australia. For Ecol Manage 110:343–352
Burnham KP, Anderson DR (2002) Model selection and multimodel inference. Springer, New York
Castro-Díez P, González-Muñoz N, Alonso A, Gallardo A, Poorter L (2009) Effects of exotic invasive trees on nitrogen cycling: a case study in Central Spain. Biol Invasions 11:1973–1986
Castroviejo S (1986–2008) Flora Ibérica. Plantas vasculares de la Península Ibérica e Islas Baleares. (todos los vols). Real Jardín Botánico-CSIC, Madrid
Chigineva NI, Aleksandrova AV, Tiunov AV (2009) The addition of labile carbon alters litter fungal communities and decreases litter decomposition rates. Appl Soil Ecol 42:264–270
Cornelissen JHC (1996) An experimental comparison of leaf decomposition rates in a wide range of temperate plant species and types. J Ecol 84:573–582
Cornwell WK, et al (2008) Plant species traits are the predominant control on litter decomposition rates within biomes worldwide. Ecol Lett 11:1065–1071
Cotrufo MF, Briones MJI, Ineson P (1998) Elevated CO2 affects field decomposition rate and palatability of tree leaf litter: importance of changes in substrate quality. Soil Biol Biochem 30:1565–1571
Di Tomaso JM (1998) Impact, biology and ecology of saltcedar (Tamarix spp.) in the southwestern United States. Weed Technol 12:326–336
Drenovsky RE, Batten KM (2007) Invasion by Aegilops triuncialis (barb goatgrass) slows carbon and nutrient cycling in a serpentine grassland. Biol Invasions 9:107–116
Dukes JS, Mooney HA (2004) Disruption of ecosystem processes in western North America by invasive species. Rev Chil Hist Nat 77:411–437
Ehrenfeld JG (2003) Effects of exotic plant invasions on soil nutrient cycling processes. Ecosystems 6:503–523
Epstein E (1994) The anomaly of silicon in plant biology. Proc Natl Acad Sci USA 91:11–17
Fowler J, Cohen L (1992) Practical statistics for field biology. Wiley, London
Gallardo A, Merino J (1992) Nitrogen immobilization in leaf litter at two Mediterranean ecosystems of SW Spain. Biogeochemistry 15:213–228
Gallardo A, Merino J (1993) Leaf decomposition in two Mediterranean ecosystems of southwest Spain, influence of substrate quality. Ecology 74:152–161
Gallardo A, Parama R (2007) Spatial variability of soil elements in two plant communities of NW Spain. Geoderma 139:199–208
Garnier E, et al (2004) Plant functional markers capture ecosystem properties during secondary succession. Ecology 85:2630–2637
Gasso N, et al (2009) Exploring species attributes and site characteristics to assess plant invasions in Spain. Divers Distrib 15:50–58
Hättenschwiler S, Vitousek PM (2000) The role of polyphenols in terrestrial ecosystem nutrient cycling. Trends Ecol Evol 15:238–243
Hobbie SE (1992) Effects of plant-species on nutrient cycling. Trends Ecol Evol 7:336–339
Knight KS, Kurylo JS, Endress AG, Stewart JR, Reich PB (2007) Ecology and ecosystem impacts of common buckthorn (Rhamnus cathartica): a review. Biol Invasions 9:925–937
Knorr M, Frey SD, Curtis PS (2005) Nitrogen additions and litter decomposition: a meta-analysis. Ecology 86:3252–3257
Kolar CS, Lodge DM (2001) Progress in invasion biology: predicting invaders. Trends Ecol Evol 16:199–204
Kueffer C, Klingler G, Zirfass K, Schumacher E, Edwards PJ, Gusewell S (2008) Invasive trees show only weak potential to impact nutrient dynamics in phosphorus-poor tropical forests in the Seychelles. Funct Ecol 22:359–366
Lautner S, Ehlting B, Windeisen E, Rennenberg H, Matyssek R, Fromm J (2007) Calcium nutrition has a significant influence on wood formation in poplar. New Phytol 173:743–752
Levine JM, Vila M, D’Antonio CM, Dukes JS, Grigulis K, Lavorel S (2003) Mechanisms underlying the impacts of exotic plant invasions. Proc R Soc Lond B Biol Sci 270:775–781
Liao CZ, et al (2008) Altered ecosystem carbon and nitrogen cycles by plant invasion: a meta-analysis. New Phytol 177:706–714
Mack RN, Simberloff D, Lonsdale WM, Evans H, Clout M, Bazzaz FA (2000) Biotic invasions: causes, epidemiology, global consequences, and control. Ecol Appl 10:689–710
Manders PT, Richardson DM (1992) Colonization of Cape fynbos communities by forest species. For Ecol Manage 48:277–293
Marko PD, Gross EM, Newman RM, Gleason FK (2008) Chemical profile of the North American native Myriophyllum sibiricum compared to the invasive M. spicatum. Aquat Bot 88:57–65
Poorter H, Villar R (1997) The fate of acquired carbon in plants: chemical composition and construction costs. In: Bazzaz FA, Grace J (eds) Plant resource allocation. Academic Press, San Diego, pp 39–72
Pyšek P, Richardson DM (2007) Traits associated with invasiveness in alien plants: where do we stand? In: Caldwell MM (ed) Biological invasions, vol 193. Springer, Berlin Heidelberg, pp 97–125
Pyšek P, Richardson DM, Rejmánek M, Webster GL, Williamson M, Kirschner J (2004) Alien plants in checklist and floras: towards better communication between taxonomist and ecologist. Taxon 53:131–143
Quested HM, et al (2003) Decomposition of sub-arctic plants with differing nitrogen economies: a functional role for hemiparasites. Ecology 84:3209–3221
Quested HM, Callaghan TV, Cornelissen JHC, Press MC (2005) The impact of hemiparasitic plant litter on decomposition: direct, seasonal and litter mixing effects. J Ecol 93:87–98
R Development Core Team (2009) R: a language and environment for statistical computing. R foundation for Statistical Computing, Vienna. ISBN 3-900051-07-0. http://www.R-project.org
Richardson DM, Pyšek P, Rejmanek M, Barbour MG, Panetta FD, West CJ (2000) Naturalization and invasion of alien plants: concepts and definitions. Divers Distrib 6:93–107
Rubio A, Escudero A (2005) Effect of climate and physiography on occurrence and intensity of decarbonation in Mediterranean forest soils of Spain. Geoderma 125:309–319
Sanz Elorza M, Dana Sanchez D, Sobrino Vesperinas E (2004) Atlas de las Plantas Alóctonas Invasoras en España. Ministerio de Medio Ambiente, Madrid
Sayer EJ (2006) Using experimental manipulation to assess the roles of leaf litter in the functioning of forest ecosystems. Biol Rev 81:1–31
Sims GK, Ellsworth TR, Mulvaney RL (1995) Microscale determination of inorganic nitrogen in water and soil extracts. Commun Soil Sci Plant Anal 26:303–316
Sneller FEC, et al (1999) Toxicity of arsenate in Silene vulgaris, accumulation and degradation of arsenate-induced phytochelatins. New Phytol 144:223–232
Soltis DE, et al (2000) Angiosperm phylogeny inferred from 18S rDNA, rbcL, and atpB sequences. Bot J Linn Soc 133:381–461
Strauss SY, Webb CO, Salamin N (2006) Exotic taxa less related to native species are more invasive. Proc Natl Acad Sci USA 103:5841–5845
Subler S, Blair JM, Edwards CA (1995) Using anion-exchange membranes to measure soil nitrate availability and net nitrification. Soil Biol Biochem 27:911–917
Swift MJ, Heal OW, Anderson JM (1979) Decomposition in terrestrial ecosystems. In: Studies in ecology, vol 5. Blackwell, Oxford
Valéry L, Fritz H, Lefeuvre JC, Simberloff D (2008) In search of a real definition of the biological invasion phenomenon itself. Biol Invasions 10:1345–1351
Villar R, Robleto JR, De Jong Y, Poorter H (2006) Differences in construction costs and chemical composition between deciduous and evergreen woody species are small as compared to differences among families. Plant Cell Environ 29:1629–1643
Vitousek PM, Howarth RW (1991) Nitrogen limitation on land and in the sea—how can it occur? Biogeochemistry 13:87–115
Xuluc-Tolosa FJ, Vester HFM, Ramirez-Marcial N, Castellanos-Albores J, Lawrence D (2003) Leaf litter decomposition of tree species in three successional phases of tropical dry secondary forest in Campeche, Mexico. For Ecol Manage 174:401–412
Yelenik SG, Stock WD, Richardson DM (2007) Functional group identity does not predict invader impacts: differential effects of nitrogen-fixing exotic plants on ecosystem function. Biol Invasions 9:117–125
Acknowledgements
Thanks are due to Margarita Costa-Tenorio, Antonio Gallardo, Rob Broekman and Manuel Godoy for field help, supplying ion exchange resin membranes and lab assistance, respectively. Financial support was provided by the Spanish Ministry for Education and Science grants RASINV GL2004-04884-C02 02/BOS (as part of the coordinate project RINVE) and CGL2007-61873/BOS.
Author information
Authors and Affiliations
Corresponding author
Additional information
Communicated by Stephan Hattenschwiler.
Electronic supplementary material
Below is the link to the electronic supplementary material.
Rights and permissions
About this article
Cite this article
Godoy, O., Castro-Díez, P., Van Logtestijn, R.S.P. et al. Leaf litter traits of invasive species slow down decomposition compared to Spanish natives: a broad phylogenetic comparison. Oecologia 162, 781–790 (2010). https://doi.org/10.1007/s00442-009-1512-9
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00442-009-1512-9