
Abstract
We investigated the relationship between plant diversity and ecological function (production and nutrient cycling) in tropical tree plantations. Old plantations (65–72 years) of four different species, namely Araucaria cunninghamii, Agathis robusta, Toona ciliata and Flindersia brayleyana, as well as natural secondary forest were examined at Wongabel State Forest, in the wet tropics region of Queensland, Australia. Two young plantations (23 years) of Araucaria cunninghamii and Pinus caribaea were also examined. The close proximity of the older plantations and natural forests meant they had similar edaphic and climatic conditions. All plantations had been established as monocultures, but had been colonised by a range of native woody plants from the nearby rainforest. The extent to which this had occurred varied with the identity of the plantation species (from 2 to 17 species in 0.1 ha blocks). In many cases these additional species had grown up and joined the forest canopy. This study is one of the few to find a negative relationship between overstorey plant diversity and productivity. The conversion of natural forest with highly productive, low-diversity gymnosperm-dominated plantations (young and old Araucaria cunninghamii and Pinus caribaea) was found to be associated with lower soil nutrient availability (approximately five times less phosphorus and 2.5 times less nitrogen) and lower soil pH (mean = 6.28) compared to the other, less productive plantations. The dominant effects of two species, Araucaria cunninghamii and Hodgkinsonia frutescens, indicate that ecosystem functions such as production and nutrient availability are not determined solely by the number of species, but are more likely to be determined by the characteristics of the species present. This suggests that monoculture plantations can be used to successfully restore some functions (e.g. nutrient cycling and production), but that the level to which such functions can be restored will depend upon the species chosen and site conditions.
Similar content being viewed by others

Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
Nearly 150 years ago, Darwin (1859) proposed that diverse plant communities were more productive. Over the last 20 years, ecologists have focussed on determining the nature of the relationship between biodiversity and ecosystem function, but Darwin’s proposals have proven controversial as well as difficult to verify (Huston et al. 2000; Kaiser 2000; Loreau et al. 2001; Mouquet et al. 2002; Naeem et al. 1994; Schwartz et al. 2000; Walker 1992).
Studies using experimental communities have largely found evidence of a positive relationship between plant diversity and productivity (Balvanera and Aguirre 2006; Cardinale et al. 2006; Hector et al. 1999; Pfisterer and Schmid 2002; Reich et al. 2001; Symstad and Tilman 2001; Tilman and Downing 1994; Tilman et al. 1996, 1997a). In these cases, experimental communities differing in plant diversity were defined as communities that had been specifically assembled for research. When this same relationship is investigated within natural and managed (plantation) forest communities, the findings are not as straightforward. If we take, for example, eight studies that specifically focussed on forest communities, three found a negative relationship (Huston 1980; Lugo 1992; Wardle et al. 1997), four found a positive relationship (Caspersen and Pacala 2001; Erskine et al. 2006; Troumbis and Memtsas 2000; Vila et al. 2007) and one found no relationship (Vila et al. 2003). The natural and managed communities used in these studies had not been assembled for research purposes and included monocultures and mixed species forest plantations.
This difference between experimental and natural or managed community studies has resulted in a vigorous debate amongst researchers (Hughes and Petchey 2001; Kaiser 2000; Mittelbach et al. 2001). The debate revolves around the interpretation of the positive results found in experimental community studies. Two hypotheses have been suggested to explain the observations: the “niche complementarity” hypothesis and the “sampling effect” hypothesis.
The niche complementarity hypothesis proposes that species-rich communities are able to more efficiently access and utilise limiting resources because they contain species with a diverse array of ecological attributes. The ecosystem is thought to be more functionally complete because these species complement each other, allowing them to optimise the use of resources. As a consequence, more diverse ecosystems are also more productive and have also been described as having less nutrients available because, overall, the uptake is more efficient (Hector 1998; Loreau 1998; Tilman et al. 1997b). On the other hand, the sampling effect hypothesis proposes that more biologically diverse communities have increased productivity because they are more likely to contain at least one species that is particularly efficient in resource usage. That is, only one or two species within the community may be largely responsible for most of the production. It follows from this that simple monocultures of these species would probably achieve the same high levels of production as some multi-species communities (Aarssen 1997; Huston 1997, 1980; Tilman et al. 1997b).
The relationship between diversity and nutrient cycling is also complicated. Some experimental studies have found that increasing diversity can be accompanied by a reduction in the availability of soil nutrients. That is, there is a net transfer of nutrients from soils to plant biomass as more species with different functional traits are added to the community. In these circumstances, nutrients are conserved and nutrient cycling has become more “sustainable” (Tilman and Downing 1994; Tilman et al. 1996, 1997a). By contrast, other studies have found that nutritional responses to increased diversity are maximised with a relatively small number of species or that the soil nutritional status is primarily influenced by the functional characteristics of the dominant species, rather than simply the number of species involved (Ewel et al. 1991; Hooper et al. 2005). The complexity arises because the definition of a “sustainable ecosystem” will vary greatly depending on the type of habitat (grassland, forest, pasture), the original conditions of the site (productive or unproductive), the environmental heterogeneity over time, and the future plans for the site(s) in question, including whether it will be used for resource extraction or for conservation. Therefore, it is essential that studies examining the relationship between diversity and nutrient cycling are highly explicit in how they define “sustainable”.
There have been very few studies conducted in communities as rich in species as tropical forests (Schlapfer and Schmid 1999; Vandermeer et al. 2002; Vila et al. 2003). Extensive tracts of these areas have been deforested in recent decades, but large areas have also been reforested. Most of this reforestation has used monoculture plantations although some even-aged mixed species plantations have also been used (Dobson et al. 1997; Erskine et al. 2006; Lamb 1998; Lamb and Gilmour 2003; Parrotta 1992; Sayer et al. 2004). The niche complementarity hypothesis suggests that mixed species plantations might restore ecosystem function more effectively than monocultures (Kelty 1992; Pretzsch 2005; Rothe and Binkley 2001). On the other hand, traditional plantation establishment has usually relied on a few key species and is based on the operation of mechanisms inherent to the “sampling” effect hypothesis. These methods have created productive commercial timber resources, but may not have restored other ecosystem functions.
We investigate the relationships between plant diversity, production and nutrient cycling in tropical forest ecosystems in order to clarify which of the two hypotheses is operational. The design of this study is unique because the sites are a series of 60–70 year-old monoculture timber plantations, all of which are growing adjacent to natural rainforest. Over time, the plantations have been colonised by a large number of native forest species. Some of these have grown up, joined the canopy layer and transformed the simple monocultures into a series of more structurally complex, species-rich forests. Therefore, this study is a novel hybrid between natural and managed community studies and experimental community studies because we have been able to use a series of plantations whose levels of biodiversity have been driven by natural processes and are not experimentally derived.
We hypothesised that the less diverse plantation ecosystems would be more productive than the species-rich natural forest, which supports the notion of the sampling effect hypothesis. We also hypothesised that the soil nutrient availability would be higher with greater levels of tree diversity. This supports the notions of the complementarity hypothesis because, contrary to the grassland ecosystem such as those studied by Tilman and Downing (1994), Tilman et al. (1997) and Tilman (1996), having a high-level nutrient availability in the soil is defined as sustainable because the original soils in this region were characteristically fertile, being derived from basalt. The low-diversity stands are hypothesised to have a low soil nutrient availability in comparison to the high-diversity stands because of a slower nutrient turnover rate associated with the poor-quality litter.
Methods
Study area
This study was conducted within the plantations of Wongabel State Forest, which is located 5 km south of the town of Atherton in tropical north-east Queensland (17°32′S, 145°50′E). The plantations and surrounding natural forest cover 635 ha. The mean annual rainfall ranges from 1,460 to 1,690 mm, elevation ranges from 740 to 780 m and the distance from the coast is around 50 km. The soil is described as well drained and formed from young, weakly weathered basaltic lavas (Laffan 1988). Webb et al. (1984) classified the original forest of this area as complex notophyll vine forest type 5b. This forest type once covered most of the region around Atherton, but has been extensively cleared for agriculture, largely because of its occurrence on nutrient-rich soils. Today, less than 2% of the original forest remains, with Wongabel State Forest containing some of the largest fragments. As a result, the remaining complex notophyll vine forest type 5b has been classified as an “endangered” plant community.
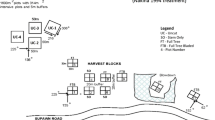
Wongabel State Forest was selected as the study site because of the existence of old plantations of four different species, namely Araucaria cunninghamii (Family: Araucariaceae, gymnosperm, native), Agathis robusta (Family: Araucariaceae, gymnosperm, native), Toona ciliata (Family: Meliaceae, angiosperm, endemic) and Flindersia brayleyana (Family: Rutaceae, angiosperm, endemic), as well as natural secondary forest (hereafter the plantations are identified by only the genus). In the 1930s and 1940s, rainforest was cleared and plantations of these species were established within Wongabel for research and wood production. During plantation establishment, remnant areas of the adjacent rainforest were selection-harvested for valuable timber species such as Toona ciliata and Castanospermum australe and then left to regenerate naturally. Remnant rainforest strips were also left around and between the plantations to act as a fire break.
The close proximity of the different forest types and their similarity in age provided a unique opportunity for a comparative analysis of production and nutrient cycling, as any effects from climatic, edaphic and successional differences are reduced to a minimum. The plantations selected for study ranged in age from 65 to 72 years, and the natural secondary forest was selection-harvested around the same time as plantation establishment (Table 1). All plantations, once established, received little stand maintenance (except for the Toona ciliata plantation) and presently have a narrow range of stand density, 500–800 stems/ha (Table 1).
Two young plantations of Pinus caribaea var. hondurensis (Family: Pinaceae, gymnosperm, exotic) and Araucaria cunninghamii (Family: Araucariaceae, gymnosperm, native) were also studied. Both were 23 years in age and located adjacent to each other and within 500 m of the older plantations. The reason for the inclusion of these young plantations was that Pinus caribaea var. hondurensis and Araucaria cunninghamii are the species the current management agency (the Queensland Department of Primary Industries) favours for future planting and harvesting.
Stand composition and productivity
Three 50 m × 20 m blocks were randomly established and surveyed within each forest type. In the strict sense of the term these were not true replicates but pseudo-replicates. We did not have access to other stands of this age in the vicinity since most plantations established around the same time have been harvested because of their high-value timber. In each block, all trees ≥10 cm in diameter at breast height (DBH) were identified and the DBH measured. These measurements were used to calculate biodiversity (as species richness and Simpson’s biodiversity index) and production [calculated as basal area (m2/ha)]. Basal area is highly correlated with tree biomass (Satoo and Madgwick 1982) and was used as an index of production in these plantations because accurate volume equations and wood density data were unavailable for most species.
Understorey biodiversity levels were assessed by randomly locating a 10 m2 plot within each of the 50 m × 20 m overstorey tree blocks and all woody plants less than 10 cm DBH were identified and recorded. Simpson’s biodiversity index was used to represent biodiversity after testing found that it was closely related to both species richness and the Shannon–Weaver index. This choice is supported by Magurran (1988) who, in a detailed analysis of different biodiversity indices, suggests that the Simpson’s biodiversity index can be a more reliable measure than the Shannon–Weaver index or species richness. Regression analysis was also used to determine the relationship between plant diversity and function, and these analyses were performed with the statistical package R, version 2.2.1.
Nutrient availability
Soil nitrogen and phosphorus availability was assessed using ion exchange resin (IER) bags. IER bags were made from plastic screen material manufactured by Swiss Screens PE 48GG (365 μm pore size; Seven Hills, NSW, Australia). The plastic material was cut and assembled into bags 5 cm2 in size. Each bag was filled with 5 g fresh weight of mixed bed resin (Dowex MR-3, Sigma, St. Louis, MO, USA) (Erskine et al. 1998). Five bags were buried within each of the 21 blocks (7 forest types, 3 replicates per forest type), two at a depth of 20 cm, two at a depth of 5 cm and one just below the leaf litter. The bags were left in situ for a total of 21 days, from the end of May to the beginning of June. During this time frame, steady rainfall occurred daily. This time of year is considered the “drizzle” quarter in the Atherton area, which is characterised by consistent but low-intensity rainfall.
Once removed from the ground, each IER bag was stored separately in a plastic bag, labelled and refrigerated at 4 °C until laboratory analysis was possible. The resin was then extracted with 20 ml of 1 M KCl solution and shaken for 30 min on an orbital shaker. The eluate was decanted then filtered, and the above steps were repeated five times to ensure thorough extraction. The bulked elution for each IER bag was analysed using three separate colorimetric methods for ammonium (NH4 +) concentration: indophenol blue (Bolleter et al. 1961); nitrate (NO −3 ) concentration: cadmium–copper reduction to nitrite (Wood et al. 1967); and phosphorus (P) concentration: malachite green (Ohno and Zibilske 1991; Vanveldhoven and Mannaerts 1987). Soil pH was measured in 1:5 soil:water suspensions.
An index of available soil nitrogen values were calculated by summing the mean ammonium and nitrate concentrations at three levels within the soil profile, specifically at 0, 5, and 20 cm. Available soil phosphorus values were also calculated by summing the mean concentrations found at each of these three levels. The summed value of the concentrations found at each of the three levels was used as an index of soil nitrogen and phosphorus availability because the A horizons of the soils at Wongabel State Forest have been described as deep (Laffan 1988), and tree roots, particularly within the older plantations, will exist throughout the soil profile and not just at one depth. Therefore, nitrogen and phosphorous availability was deemed to be more accurately assessed with an index that combined concentrations of nutrients available throughout the profile, as opposed to just one depth.
The computer software package Primer (Plymouth routines in multivariate ecological research) was used to conduct a principal component analysis (PCA) using three normalised environmental variables, including soil-available inorganic nitrogen and phosphorus in the top 20 cm and soil pH in water.
Results
Recruitment of plant diversity
All five of the older monoculture plantations had been colonised by native rainforest species from the adjoining natural forests, although large differences were found in the diversity of species now present (Table 2). Of the older plantations, Araucaria was found to have the lowest overstorey species richness and Simpson’s biodiversity values, while the Flindersia plantation and the natural secondary forest had the highest. The order from lowest to highest overstorey plant diversity was found to be Araucaria, Agathis, Toona, Flindersia, and the natural secondary forest (Table 2).
The levels of diversity in the understorey vegetation were quite different to those in the overstorey vegetation. In this case, most of the plantations and the natural forest had similar numbers of plant species (between four and eight species per 10 m2 plot). However, diversity levels varied and were low in sites with large amounts of overstorey diversity, such as the Flindersia plantation and the natural forest, but high in sites such as the Araucaria plantations which had much lower numbers of overstorey species. This difference is related to the dominance of Hodgkinsonia frutescens within the understorey of the natural secondary forest and Flindersia plantations, where it represents more than 90% of all individuals. The dominance of this understorey shrub is a distinct characteristic of the endangered forest type, complex notophyll vine forest type 5b; this shrub is found in only one other forest type in Australia (Mabi Forest Working Group 2001).
Relationship between biodiversity and productivity
There is a significant negative relationship between the diversity (measured as Simpson’s biodiversity index) of overstorey tree species and the stand’s production as measured by total basal area (Fig. 1). As the diversity of species within the overstorey increased, production levels decreased steeply. The Araucaria and Agathis plantations had basal area values greater than 60 m2/ha, but Simpson’s biodiversity index values of mostly less than 0.3. Toona and Flindersia plantations and natural secondary forests had basal area values of mostly less than 65 m2/ha but ranged between 0.50 and 0.92 in Simpson’s biodiversity index values.

The relationship between Simpson’s biodiversity index (overstorey) in 0.1 ha plots and production (basal area). AC, Araucaria cunninghamii; YAC, young Araucaria cunninghamii; PC, Pinus caribaea; AR, Agathis robusta; TC, Toona ciliata; FB, Flindersia brayleyana; NF, natural secondary forest. The numbers refer to blocks 1, 2, 3 within each forest type
Relationship between biodiversity and nutrient availability
A positive relationship was found between the diversity of canopy trees and soil nutrient availability (Fig. 2). A stronger negative relationship was found, however, between the diversity of understorey plants and available soil nitrogen and phosphorus (Fig. 3). This suggests some relationship between increased soil nutrient availability and the recolonisation of the sites by the characteristic understorey of the area, which is dominated by the shrub Hodgkinsonia frutescens (Rubiaceae). This species is present within the Agathis and Toona plantations, but is particularly common in the Flindersia plantation and natural secondary forest (Table 2).

The relationship between canopy tree diversity and soil nitrogen and phosphorus availability in the top 20 cm, as measured using IER bags. AC, Araucaria cunninghamii; YAC, young Araucaria cunninghamii; PC, Pinus caribaea; AR, Agathis robusta; TC, Toona ciliata; FB, Flindersia brayleyana; NF, natural secondary forest. The numbers refer to blocks 1, 2, 3 within each forest type

The relationship between understorey tree and shrub diversity and soil nitrogen and phosphorus availability in the top 20 cm, as measured using IER bags. AC, Araucaria cunninghamii; YAC, young Araucaria cunninghamii; PC, Pinus caribaea; AR, Agathis robusta; TC, Toona ciliata; FB, Flindersia brayleyana; NF, natural secondary forest. The numbers refer to blocks 1, 2, 3 within each forest type
The natural secondary forest, Toona, and Flindersia plantations all have similar degrees of species richness and abundance in the overstorey and similar high levels of available soil nitrogen and phosphorus (Tables 2, 3). These four forest types differ from the young and old Araucaria and Pinus plantations which have different floristic assemblages in their overstorey [see “Electronic supplementary material” (ESM)] and lower levels of soil nitrogen and phosphorus availability. The Agathis plantation also has a floristically distinct species assemblage in the overstorey in comparison with the other forest types (see “ESM”). However, like the natural secondary forest, Toona, and Flindersia forest types the availability of soil nitrogen and phosphorus is relatively high.
The Fischer’s Individual Error Rate post hoc test was conducted on the characteristics of the soil collected within each block of each forest type (Table 3). The results suggest that the Araucaria and the Pinus plantations differed significantly from the Agathis, Toona, and Flindersia plantations and the natural forest, particularly with regard to soil phosphorus availability (a similar difference was found between these species with respect to soil nitrogen availability). The soil in the young and old Araucaria and the Pinus plantations tended to be slightly more acidic and to have lower nitrogen and phosphorus availability then the other more floristically diverse forest types.
The similarity and dissimilarities between the forest types based on normalised nutrient availability data collected at each block is shown in Fig. 4, a two-dimensional PCA ordination. The data included available soil inorganic nitrogen (ammonium-N and nitrate-N) in the top 20 cm, available soil phosphorus in the top 20 cm, and soil pH. This ordination shows the close association between the Agathis, Toona, Flindersia and natural secondary forest types because of their proximity on the two-dimensional plane. PC1 (x-axis) and PC2 (y-axis) together account for 71.6% of the total variability between the forest sites, with PC1 describing 46.9% of the variance alone. Figure 4 also shows the difference between the old and young Araucaria and Pinus forest types because of their distance from each other and the other forest types.

Two-dimensional PCA ordination of three normalised environmental variables, including soil-available inorganic nitrogen and phosphorus in the top 20 cm and soil pH in water for all seven forest types and 21 blocks (% variance explained = 71.6%). AC, Araucaria cunninghamii; YAC, young Araucaria cunninghamii; PC, Pinus caribaea; AR, Agathis robusta; TC, Toona ciliata; FB, Flindersia brayleyana; NF, natural secondary forest. The numbers refer to blocks 1, 2, 3 within each forest type
Discussion
Recruitment of plant diversity
Over time, all of the plantations studied have changed from monocultures to more diverse communities because of the colonisation of native plant species from the surrounding rainforest buffers. This makes our experimental design a hybrid between natural and managed and experimental community studies, which is both an advantage and a limitation for exploring the central question of the relationship between biodiversity and ecosystem function. Its advantage is that each plantation has experienced different rates of recruitment within both the overstorey and the understorey, and this recruitment was driven largely by the characteristics of the plantation species and stochastic forces. Different rates of recruitment have subsequently altered tree densities, overstorey cover and hence microenvironments. Natural ecosystems are more likely to acquire diversity in this manner being driven by the frequency and intensity of disturbances and the conditions left by current or previous species on the site (Connell 1978; Connell and Slatyer 1977). This arguably makes our study one of only a few able to assess the function of diversity in a forest ecosystem that has undergone the complex successional dynamics that characterise natural environments (Vila et al. 2007; Caspersen and Pacala 2001; Troumbis and Memtsas 2000; Vila et al. 2003; Wardle et al. 1997), as opposed to an artificially derived system where the species have been selected and then assembled at the same time.
However, this experimental design is also a limitation, as the diversity found within these plantations is not an independent variable, but is instead a function of the production rate and nutrient use of the plantation species themselves. But the five older plantations surveyed have been growing with different degrees of diversity for more than 65 years. Therefore, it is probable that the acquired diversity has been established for long enough that it has influenced the functioning of these systems.
The highest amount of new plant diversity was found in the Flindersia and Toona plantations, while much lower recruitment was recorded in the gymnosperm plantations of Araucaria and Pinus. The recruitment under the third gymnosperm species, Agathis, was intermediate between these two groups. This suggests that the level of recolonisation by native plant species is strongly influenced by the characteristics of the plantation species. Specifically, the broad-leafed gymnosperm species (Agathis) showed a greater level of recruitment than the needle-like gymnosperm species (Araucaria and Pinus). Keenan et al. (1997), Parrotta (1995) and Lugo (1992) found similar results in studies conducted within tropical forest communities.
Another limitation of our experimental design is that the Agathis and Toona plantations were thinned (Table 2), while the Araucaria and Flindersia plantations were not. This thinning may account for the different levels of biodiversity within the overstorey and understorey, particularly when comparing the two high-production gymnosperm plantations, Agathis and Araucaria. It may be that the Agathis plantation has recruited more diversity within both its overstorey and understorey because of the thinning to 400 trees/ha in 1970 (Table 2). Information is not available, however, on the amount of timber extracted from these plantations during thinning. To explore this further we examined the relationship between the average basal area/ha and the average stems/ha (all species not just the original plantation species) found in each forest type using regression analysis and found that there was no relationship (R 2 = 0.014, P = 0.717, n = 15). We applied the same regression analysis to data from the Araucaria and Agathis plantations, but again did not find a significant relationship (R 2 = 0.58, P = 0.078, n = 6). However, the correlation between the number of stems per ha and the basal area was high, which suggests that we may have found a significant result with more data points. Different thinning regimes is an important confounding effect to note, but does not entirely explain the varying rates of acquisition of diversity into the overstorey of the plantations.
The biodiversity–productivity relationship
Overall, the differences in the biodiversity–production balance were strongly related to the productivity of the plantation species that dominated each multi-species community. This provides evidence of the potentially influential role of the “sampling effect” in generating the positive relationship found within experimental communities (Huston 1980; Lugo 1992; Wardle et al. 1997). For example, Wardle (1997) attributed his finding of a negative relationship between plant diversity and production on islands in the northern boreal forest zone of Sweden to the dominance of one gymnosperm species, Pinus sylvestris. On islands where this species predominated, the forest ecosystems were characterised by increased productivity and reduced plant diversity. Lugo (1992) compared overstorey and understorey diversity in different tropical tree plantations and natural secondary forest and also found less production in forests with greater levels of diversity. He attributed increased production levels found in low diversity plantations to the dominance of an especially productive gymnosperm species, Pinus caribaea var. morelet.
However, not all studies in natural or managed communities have found a negative relationship between biodiversity and production. Troumbis and Memtsas (2000) examined the functional role of diversity in natural communities in the shrublands of the Mediterranean and found increasing production with increased biodiversity. This contrasting relationship might be a consequence of the generally low soil fertility at these sites. In this case, additional species may have proven advantageous by increasing the amount of nitrogen fixed or mobilising nutrients.
Caspersen and Pacala (2001) and Vila et al. (2007) also found a positive relationship between species richness and productivity in forest communities. Caspersen and Pacala (2001) analysed the extensive United States Service Forest Inventory and Analysis Database, which comprises data from across the USA. Unfortunately, their results are highly confounded as they did not consider environmental heterogeneity in their analysis. Vila et al. (2007) also used inventory data from the National Forest Inventory of Spain, but concentrated on the region within Catalonia. They overcame the shortcomings of Caspersen and Pacala (2001) by incorporating differences in environmental conditions, forest structure and management as explanatory variables within their analysis. They found that on average forest plantations with multiple species were 30% more productive, but the results also suggested that dominant tree species strongly influenced the level of productivity that was finally achieved. More specifically, the presence of deciduous species (Quercus genera) positively affected the productivity of the forest stand, which they suggest as evidence of the sampling effect. Vila et al. (2007) also suggest successional stage, although not considered in their study, as another possible determinant of the relationship between diversity and productivity, and that the functioning of young plantations may be positively influenced by increased species richness.
Erskine et al. (2006) analysed an inventory data collected for an Australian federal government initiative called the Community Rainforest Restoration Program and also found a positive relationship between production and increasing species richness. This initiative saw the establishment of more than 1,782 ha of tropical forest plantations varying from monocultures to diverse mixtures on private land in the 1990s. These plantations were established in the same tropical region as those included in this study, but are examples of early successional communities when compared to the plantations in this study that have been growing for over 65 years. It may be the relationship between diversity and production changes at the later stages of successional development as suggested by Vila et al. (2007).
An explanation for the disparity between studies may be differences in ecological scale. Several reviews have suggested the need for a distinction in the results of studies within and between communities, particularly communities defined by different scales of ecological association (Crawley and Harral 2001; Loreau et al. 2001; Mittelbach et al. 2001; Naeem and Wright 2003). Two detailed analyses of the literature, Mittelbach et al. (2001) and Waide et al. (1999), suggested that the nature of the relationship between biodiversity and production may vary with the scale of the study undertaken. More specifically, it may depend on whether the relationship was examined within communities (small spatial scale) or between communities (large spatial scale), but there is disagreement as to which relationship characterises which scale. Mittelbach et al. (2001) found that the probability of a positive relationship increased with spatial scale, which may explain the positive results of Vila et al. (2007) and Erskine et al. (2006); while Waide et al. (1999) found that the probability of a hump-shaped positive relationship increased with a larger spatial scale. Waide et al. (1999) also found that less than 12% of studies pointed to a negative relationship between diversity and productivity.
The different levels of species diversity that characterise the communities under investigation may also explain the nature of these different relationships. Troumbis and Memtsas (2000) and Vila et al. (2007) investigated Mediterranean forest communities, although the study by Vila et al. (2007) included forests from a more diverse geographic region, including Mediterranean, sub-Mediterranean, Euro-Siberian and even Boreo-alpine chorology. Neither study investigated the diversity–productivity relationship in regions where the potential species pools are as diverse as in rainforest ecosystems. The negative relationship we found suggests that the relationship between biodiversity and ecosystem function may not be generalised across ecosystems or even across forest ecosystems, which further suggests that the relationship is likely to be influenced heavily by site conditions, including the size of the species pool, climatic conditions, as well as the spatial scale chosen. Erskine et al. (2006) studied mixed species plantations in the same tropical region as the present study. The species richnesses found in the younger plantations studied by Erskine et al. (2006) were, however, much lower than those found in the present study (eight as opposed to 17 tree species). Also, the mixtures of species were planted at the same time, whereas the acquisition of diversity in this study was driven by natural processes.
Biodiversity–nutrient availability relationship
Plantations at Wongabel that had acquired a high amount of native plant diversity in the overstorey also had high levels of soil nutrient availability, similar to those in the species-rich natural forest. This finding seems to agree with the hypothesis of this study, namely that there is a positive relationship between overstorey biodiversity and nutritional availability. The IER bags are assumed to provide an indication of nutrient availability in the soil as the resin exchanges cations and anions with soil water in a similar manner to tree roots (Binkley 1984; Binkley and Matson 1983; Mo et al. 2003). However, the alternative explanation—that nutritional availability is largely a consequence of the attributes of the dominant species in each type—also has some relevance here. There are several pieces of evidence supporting this suggestion. One is the close relationship between high nutrient availability and a high relative density of the common understorey plant, Hodgkinsonia frutescens. The underlying mechanism for this relationship is not clear, although there seems to be a stronger relationship between the density of Hodgkinsonia frutescens and the index of available phosphorus than with the index of available nitrogen. A second piece of evidence comes from our analyses of soil nutrient availability and soil pH (Table 3), which indicated that the conifer-dominated forest types generally had lower levels of nitrogen and phosphorous than the other forests, and that the genera with needle-like leaves (Araucaria and Pinus) had lower levels than the broad-leafed conifer (Agathis).
There are several possible explanations for these observations. One is that the Araucaria plantations are more productive than the others, as the basal area was approximately 1.5 times higher than in most of the other forest types (Table 1), suggesting that more of the nutrient capital of the site is stored in this aboveground biomass. However, the young Araucaria and the young Pinus plantations, which are fast-growing but are yet to develop large aboveground biomass also had low soil nutrient availability levels, like the old Araucaria plantation.
Another explanation concerns the differential quality of the litters being produced. Since Pinus and Araucaria are gymnosperm species, the microbial activity and litter decomposition is likely to be less than in broad-leafed species, leading to reductions in the availability of phosphorus, nitrate and ammonium (Carney et al. 2004; Hattenschwiler 2005; Tinker 1984). The soil conditions created by gymnosperm leaf litter have been found to be deleterious to soil microbial communities, perhaps because of increased soil acidity (Lugo 1992; Mo et al. 2003). This was the case in this study, where the soil from both the young and old Araucaria and the Pinus plantations was found to be more acidic than that from the other forest types (Table 3). Lugo (1992) also concluded that differences in the quality of gymnosperm litter and angiosperm litter, rather than tree diversity, accounted for the differences in nutrient cycling between tropical tree plantations and natural secondary forest.
If the plant diversity within a gymnosperm-dominated plantation is increased, the availability of nutrients may be increased. The soil sampled at the Agathis plantation was found to have a high nitrogen and phosphorus availability, despite Agathis being a gymnosperm species of similar stature to Araucaria and also having leaves with waxy cuticles (Ash 1983; Boland et al. 1997). An obvious difference between these two forest types was the understorey of the Agathis plantation, which was found to contain species that were more characteristic of the natural secondary forest and the original complex notophyll vine forest 5b, particularly Hodgkinsonia frutescens (Fig. 3, “ESM”). The results do not show the mechanism behind the increased diversity found within the Agathis plantation. It may have been driven by the characteristics of the dominant species or the fact that this plantation was thinned and the Araucaria plantation was not. Prescott (2002) found that nitrogen levels in the soil increased in canopy openings, which suggests that thinning operations may have even increased the availability of soil nutrients initially, permitting the colonisation of a more characteristic understorey of the local area.
The results also suggest that the angiosperm-dominated understorey within the Agathis plantation may have improved the quality of the leaf litter layer, thus accounting for the increased level of available soil nitrogen and phosphorus. Hattenschwiler (2005) and Reich et al. (2005) have noted that this commonly (though not invariably) occurs in mixtures of litter of conifer and broad-leaved species. Yao et al.(2000), Carney et al. (2004) and Smith et al. (1998) also found that differences in plant communities could affect soil microbial communities and subsequently nutrient availability indirectly through variations in the type, complexity and amount of organic matter.
The diverse understorey within the Agathis plantation may have also improved soil nutrient availability because of differences in rooting morphologies and mycorrhizae, which have altered the capacity of the community to explore soil resources and have led to higher soil heterogeneity and increased microbial diversity (Yao et al. 2000; Smith et al. 1998; Carney et al. 2004; Rothe and Binkley 2001). In a long-term common garden experiment, Reich et al. (2005) found that tree species can affect the chemistry of soils directly because of varying leaf litter chemistry and indirectly by affecting the activity of earthworms. They also found that gymnosperm and angiosperm species had divergent effects on the soil, which was driven by varying levels of calcium in the litter (Reich et al. 2005).
Based on our findings, the relationship between biodiversity and ecosystem function is not as straightforward as originally proposed by Darwin and found in experimental community studies. If a species-rich tropical forest is replaced by a species-poor plantation, production and the sustainability of nutrient cycling will be affected. This will not be simply because of low species richness, but because of the characteristics of the species that are present, the stage of successional development and the history of management actions. Species such as Araucaria cunninghamii and Hodgkinsonia frutescens were found to have a significant impact on the functioning of forest plantations. Plantations established with gymnosperm species such as Araucaria cunninghamii are likely to have a high level of production (basal area = 82.5 m2/ha) and a low soil nutrient availability (roughly five times less phosphorus and 2.5 times less nitrogen than the natural forest), possibly because of the effects the leaf litter has on the resident microbial and detritivore community. The presence of a species more indicative of the natural forest, such as Hodgkinsonia frutescens, in the understorey of a plantation established using another highly productive gymnosperm species Agathis robusta (basal area = 68.8 m2/ha) may improve leaf litter quality, thus contributing to an increase in soil nutrient availability (5.5 times more phosphorus and double the nitrogen compared to the old Araucaria plantation).
Monocultural plantations can be used to successfully restore some ecological functions if the most appropriate species are selected and some local biodiversity is present. Plantations of Toona and Flindersia were found to facilitate the recruitment of an increasingly diverse range of native plant species within the overstorey and understorey (on average seven and 13 overstorey species per plot, respectively), while Agathis was found to recruit an understorey characteristic of the original forest type. These three plantations were also found to have levels of soil phosphorus and nitrogen availability comparable to those of the natural secondary forest. The Araucaria and Pinus plantations were found to have recruited much lower levels of native plant diversity within the overstorey and understorey (on average 2–4 overstorey species, respectively) and were also found to have created environments with low levels of soil nitrogen and phosphorus availability. Therefore, plantation species that allow for the recolonisation of native species are also likely to show an increase in the diversity of organic material within the leaf litter, root characteristics and ultimately the heterogeneity of the soil, which in turn will ensure the maintenance of the original edaphic conditions.
References
Aarssen LW (1997) High productivity in grassland ecosystems: effects by species diversity or productive species? Oikos 80:183–184
Ash J (1983) Growth rings in Agathis robusta and Araucaria cunninghamii from tropical Australia. Aust J Bot 31:269–275
Balvanera P, Aguirre E (2006) Tree diversity, environmental heterogeneity, and productivity in a Mexican tropical dry forest. Biotropica 38:479–491
Binkley D (1984) Ion-exchange resin bags—factors affecting estimates of nitrogen availability. Soil Sci Soc Am J 48:1181–1184
Binkley D, Matson P (1983) Ion-exchange resin bag method for assessing forest soil-nitrogen availability. Soil Sci Soc Am J 47:1050–1052
Boland DJ et al. (1997) Forest trees of Australia. CSIRO Publishing, Collingwood, VIC, Australia
Bolleter WT, Bushman CJ, Tidwell MD (1961) Spectrophotometric determination of ammonium as indophenol. Anal Chem 33:592–594
Cardinale BJ et al. (2006) Effects of biodiversity on the functioning of trophic groups and ecosystems. Nature 443:989–992
Carney KM, Matson PA, Bohannan BJ (2004) Diversity and composition of tropical soil nitrifiers across a plant diversity gradient and among land-use types. Ecol Lett 7:684–694
Caspersen JP, Pacala SW (2001) Successional diversity and ecosystem function. Ecol Res 16:895–903
Connell JH (1978) Diversity in tropical rain forests and coral reefs. Science 199:1302–1310
Connell JH, Slatyer RO (1977) Mechanisms of succession in natural communities and their role in community stability and organization. J Ecol 111:1119–1144
Crawley MJ, Harral JE (2001) Scale dependence in plant diversity. Science 291:864–868
Darwin C (1859) The origin of species by means of selection. Murray, London
Dobson AP, Bradshaw AD, Baker AJM (1997) Hopes for the future: restoration ecology and conservation biology. Science 277:515–522
Erskine PD, Bergstorm DM, Schmidt S, Stewart G, Tweedie CE, Shaw J (1998) Subantartic Macquarie Island—a model ecosystem for studying animal derived nitrogen sources using 15N natural abundance. Oecologia 117:187–193
Erskine P, Lamb D, Bristow M (2006) Tree species diversity and ecosystem function: can tropical multi-species plantations generate greater productivity? For Ecol Manag 233:205–210
Ewel J, Mazzarino MJ, Berish CW (1991) Tropical soil fertility changes under monocultures and successional communities of different structure. Ecol Appl 3:289–302
Hattenschwiler S (2005) Effects of tree species diversity on litter quality and decomposition. In: Scherer-Lorenzen M, Korner C, Schulze ED (eds) Forest diversity and function: temperate and boreal systems. Springer, Berlin, pp 149–162
Hector A (1998) The effect of diversity on productivity: detecting the role of species complementarity. Oikos 82:597–599
Hector A et al. (1999) Plant diversity and productivity experiments in European grasslands. Science 286:1123–1127
Hooper DU et al. (2005) Effects of biodiversity on ecosystem functioning: a consensus of current knowledge. Ecol Monogr 75:3–35
Hughes JB, Petchey OL (2001) Merging perspectives on biodiversity and ecosystem function. Trends Ecol Evol 16:222–227
Huston M (1980) Soil nutrients and tree species richness in Costa-Rican Forests. J Biogeogr 7:147–157
Huston M (1997) Hidden treatments in ecological experiments: re-evaluating the ecosystem function of biodiversity. Oecologia 110:449–460
Huston MA et al. (2000) No consistent effect of plant diversity on productivity. Science 289:1255
Kaiser J (2000) Rift over biodiversity divides ecologists. Science 289:1282–1283
Keenan R, Lamb D, Woldring O, Irvine T, Jensen R (1997) Restoration of plant biodiversity beneath tropical tree plantations in Northern Australia. For Ecol Manag 99:117–131
Kelty MJ (1992) Comparative productivity of monocultures and mixed-species stands. In: Kelty MJ, Larson BC, Oliver CD (eds) The ecology and silviculture of mixed-species forests. Kluwer, Dordrecht, pp 125–141
Laffan MD (1988) Soils and land use on the Atherton Tableland, North Queensland. CSIRO, Melbourne
Lamb D (1998) Large-scale ecological restoration of degraded tropical forest lands: the potential role of timber plantations. Restor Ecol 6:271–279
Lamb D, Gilmour D (2003) Rehabilitation and restoration of degraded forests. IUCN and WWF, Gland, Switzerland
Loreau M (1998) Biodiversity and ecosystem function: a mechanistic model. Proc Natl Acad Sci 95:5632–5636
Loreau M et al. (2001) Biodiversity and ecosystem functioning: current knowledge and future challenges. Science 294:804–808
Lugo AE (1992) Comparison of tropical tree plantations with secondary forests of similar age. Ecol Monogr 62:1–41
Mabi Forest Working Group (2001) Vanishing vegetation of far North Queensland Mabi (5b) Forest. http://www.treat.net.au/mabi/Mabi_Forest.pdf. Cited 30 August 2007
Magurran AE (1988) Ecological diversity and its measurement. Chapman and Hall, London
Mittelbach GG et al. (2001) What is the observed relationship between species richness and productivity? Ecology 82:2381–2396
Mo J, Brown S, Peng S, King G (2003) Nitrogen availability in disturbed, rehabilitated and mature forests of tropical China. For Ecol Manag 175:573–583
Mouquet N, Moore JL, Loreau M (2002) Plant species richness and community productivity: why the mechanisms that promotes coexistence matters. Oikos 5:56–65
Naeem S, Wright JP (2003) Disentangling biodiversity effects on ecosystem functioning: deriving solutions to a seemingly insurmountable problem. Nature 6:567–579
Naeem S, Thompson LJ, Lawler SP, Lawton JH, Woodfin RM (1994) Declining biodiversity can alter the performance of ecosystems. Nature 368:734–736
Ohno T, Zibilske LM (1991) Determination of low concentrations of phosphorous in soil extracts using Malachite green. Soil Sci Soc Am J 55:892–895
Parrotta J (1992) The role of plantation forests in rehabilitating degraded tropical ecosystems. Agri Eco Envir 41:115–133
Parrotta J (1995) Influence of overstorey composition on understorey colonization by native species in plantations on a degraded tropical site. J Veg Sci 6:627–636
Pfisterer AB, Schmid B (2002) Diversity-dependent production can decrease the stability of ecosystem functioning. Nature 416:84–86
Pretzsch H (2005) Diversity and productivity in forests: evidence from long-term experimental plots. In: Scherer-Lorenzen M, Korner C (eds) Forest diversity and function, vol 176. Springer, Berlin, pp 41–64
Prescott CE (2002) The influence of the forest canopy on nutrient cycling. Tree Physiol 22:1193–1200
Reich P et al. (2001) Plant diversity enhances ecosystem responses to elevated CO2 and nitrogen deposition. Nature 410:809–812
Reich PB et al. (2005) Linking litter calcium, earthworms and soil properties: a common garden test and 14 tree species. Ecol Lett 8:811–818
Rothe A, Binkley D (2001) Nutritional interactions in mixed species forests: a synthesis. Can J For Res 31:1855–1870
Satoo T, Madgwick HAI (1982) Forest biomass. Martinus Nijhoff/Dr W. Junk, The Hague
Sayer J, Chokkalingam U, Poulsen J (2004) The restoration of forest biodiversity and ecological values. For Ecol Manage 201:3–11
Schlapfer F, Schmid B (1999) Ecosystem effects of biodiversity: a classification of hypotheses and exploration of empirical results. Ecol Appl 9:893–912
Schwartz MW, Brigham CA, Hoeksema JD, Lyons KG, Mills MH, van Mantgem PJ (2000) Linking biodiversity to ecosystem function: implications for conservation ecology. Oecologia 122:297–305
Smith CK, Gholz HL, Oliveira F (1998) Soil nitrogen dynamics and plant-induced soil changes under plantations and primary forest in lowland Amazonia, Brazil. Plant Soil 200:193–204
Symstad AJ, Tilman D (2001) Diversity loss, recruitment limitation, and ecosystem functioning: lessons learned from a removal experiment. Oikos 92:424–435
Tilman D, Downing JA (1994) Biodiversity and stability in grasslands. Nature 367:363–364
Tilman D, Wedin D, Knops J (1996) Productivity and sustainability influenced by biodiversity in grassland ecosystems. Nature 379:718–721
Tilman D, Knops J, Wedin D, Reich P, Ritchie M, Siemann E (1997a) The influence of functional diversity and composition on ecosystem processes. Science 277:1300–1302
Tilman D, Lehman C, Thomson K (1997b) Plant diversity and ecosystem productivity: theoretical considerations. Proc Natl Acad Sci USA 94:1857–1861
Tinker P (1984) The role of micro-organisms in mediating and facilitating the uptake of plant nutrients from soil. Plant Soil 76:77–91
Troumbis AY, Memtsas D (2000) Observational evidence that diversity may increase productivity in Mediterranean shrublands. Oecologia 125:101–108
Vandermeer JH, Lawrence D, Symstad AJ, Hobbie S (2002) Effect of biodiversity on ecosystem functioning in managed ecosystems. In: Loreau M, Naeem S, Inchausti P (eds) Biodiversity and ecosystem functioning, synthesis and perspectives. Oxford University Press, Oxford, pp 221–233
Vanveldhoven PP, Mannaerts GP (1987) Inorganic and organic phosphate measurements in the nanomolar range. Anal Biochem 161:45–48
Vila M, Vayreda J, Gracia C, Ibanez JJ (2003) Does tree diversity increase wood production in pine forests? Oecologia 135:299–303
Vila M, Vayreda J, Comas L, Ibanez JJ, Mata T, Obon B (2007) Species richness and wood production: a positive association in Mediterranean forests. Ecol Lett 10:241–250
Waide RB, Willig MR, Steiner CF, Mittelbach GG, Gough L, Dodson SI, Juday GP, Parmenter R (1999) The relationship between productivity and species richness. Annu Rev Ecol Syst 30:257–300
Walker BH (1992) Biodiversity and ecological redundancy. Conserv Biol 6:18–23
Wardle DA, Zackrisson O, Hornberg G, Christiane G (1997) The influence of island area on ecosystem properties. Science 277:1296–1299
Webb LJ, Tracey JG, Williams WT (1984) A floristic framework of Australian rainforest. Aust J Ecol 9:169–198
Wood ED, Armstrong FA, Richards FA (1967) Determination of nitrate in sea water by cadmium-copper reduction to nitrite. J Mar Biol Ass UK 47:23–28
Yao H, He Z, Wilson M, Campbell CD (2000) Microbial biomass and community structure in a sequence of soils with increasing fertility and changing land use. Microb Ecol 40:223–237
Acknowledgments
We would like to thank Mark Hunt and Nick Kelley from the Queensland Department of Primary Industries for their support financially and operationally with the field study component of this project. We would also like to thank Tony Irvine, Mila Bristow and Kerry Hanrahan (Department of Primary Industries, Forest Operations) for their assistance and expertise, as well as two anonymous reviewers whose comments greatly improved a previous draft of this document. Finally, we would like to thank Martina Langi, whose work on rainforest plantations in Northern Queensland sparked the idea for our separate study. We declare that this study complies with the current laws of Australia, where it was performed.
Author information
Authors and Affiliations
Corresponding author
Electronic supplementary material
Below is the link to the electronic supplementary material.
Rights and permissions
About this article
Cite this article
Firn, J., Erskine, P.D. & Lamb, D. Woody species diversity influences productivity and soil nutrient availability in tropical plantations. Oecologia 154, 521–533 (2007). https://doi.org/10.1007/s00442-007-0850-8
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00442-007-0850-8