
Abstract
Bird-dispersed seeds are difficult to track, especially in the case of long-distance dispersal events. To estimate the oak dispersal distance and the seed shadow generated by the European jay (Garrulus glandarius), we inserted radio-transmitters in 239 acorns, placed them in bird-feeders and then located them by radio-tracking. Using this methodology we located the exact caching site of 94 Quercus ilex and 54 Q. suber acorns and determined the caching habitat characteristics (vegetation type, distance, spatial distribution). The results show that: (1) there is no differences in the dispersal distance distribution between the different acorn species or sizes, (2) dispersal distances range from approximately 3 m up to approximately 550 m (mean = 68.6 m; median = 49.2 m), (3) recently abandoned fields and forest tracks were the sites preferred by jays to cache acorns, whereas fields and shrublands were avoided and (4) seed shadows showed acorn aggregation zones (i.e. clusters of caches) close to the feeder as well as isolated caches at longer distances. The results also suggest that radio-transmitters are a cheap and reliable way to determine seed shadows and quantify both seed dispersal and post-dispersal seed predation for medium to large seeds.
Similar content being viewed by others
Avoid common mistakes on your manuscript.
Introduction
A long-standing challenge in ecology is the characterisation of plant dispersal, which has implications on plant regeneration, gene flow and evolution (Herrera 1995; Cain et al. 2000; Wilson and Traveset 2000). For animal-dispersed plants, characterising the dispersal curve is complicated by the difficulty of following dispersers and locating the transported seeds, especially in the case of long-distance dispersal events (Cain et al. 2000; Nathan 2006). Furthermore, seed shadow size and shape may be dependent on the landscape configuration, as animals may show preferences for distinct landscape attributes (Kollmann and Schneider 1997; Russo et al. 2006).
Inferring seed shadows from seedling recruitment has limitations not only because of differential germination and survival but also because of the difficulties involved in unambiguously identifying the mother plant (Cain et al. 2000). For fleshy-fruited endozoochorous species, a simple method based on applying a fluorescent dye to the fruits and recovering it in faecal material has been successfully applied (Levey and Sargent 2000; Levey et al. 2005; Reiter et al. 2006). For plants with large seeds that are dispersed by scatter-hoarding animals (e.g. oaks), magnetic locators (Iida 1996), plastic tags (Xiao et al. 2004; Pons and Pausas 2007a) and radioisotopes (Vander Wall 2002) have been tested for their reliability to quantify acorn movement by rodents (Forget and Wenny 2005); however, these methods are not appropriate for long-distance dispersal (e.g., by birds) as the search surface increases exponentially with distance, thus making it very difficult to locate extreme long-distance dispersal events.
A very important step forward in dispersal ecology has been the use of maternity analysis (Dow and Ashley 1996; Godoy and Jordano 2001; Grivet et al. 2005). This usually involves the genetic analysis of both the dispersed seeds and the potential mother plants. However, in addition to the difficulties involved in performing genetic analyses in oaks (Ziegenhagen et al. 2003), it is also difficult to locate the dispersed acorns because most are buried by jays (e.g. the European jay Garrulus glandarius, Bossema 1979; Blue jay Cyanocitta cristata, Darley-Hill and Johnson 1981), which are known to travel large distances. Alternatively, the maternity analysis can be performed on recruits, but then the results reflect not only the dispersal but also the outcome of predation, germination and establishment on the given location (i.e. effective dispersal or realised recruitment; Nathan et al. 2003; Greene et al. 2004; Hardesty et al. 2005). In some oaks, woodpeckers store acorns in tree trunk granaries, where it is easy for researchers to collect them and study acorn movements using genetic markers (Grivet et al. 2005), although the relation between woodpecker acorn movements and dispersal is still poorly understood. In other cases, acorn dispersal has been inferred from observations of jay flights, thereby providing a first estimate of acorn dispersal curves (Gómez 2003). However, the precise location of the acorn caches is subject to visibility conditions.
In this paper we (1) estimate the distribution of acorn dispersal distances, (2) determine the shape of the dispersal shadow (i.e. we ask to what extent jay-hidden acorns are either aggregated or randomly distributed), (3) determine which vegetation types jays prefer for hiding acorns and, finally, (4) infer the suitability of the methodology for studying acorn seed shadows and jay dispersal characteristics. To achieve these objectives, we followed individual acorns dispersed by the European jay (Garrulus glandarius) by inserting a radio-transmitter inside the acorn, leaving the tagged acorn in a bird-feeder, and then locating the acorn by radio-tracking. This method, which has seldom been used prior to the present study, has enabled us to estimate primary dispersal – i.e. distance from feeders (simulated tree) to jay caches. Previous uses of radio tags include acorn dispersal by mice (Sone and Kohno 1996) and gut passage rates and seed dispersal by flightless cassowaries (Mack and Druliner 2003). By using this method, our primary aim was to obtain direct data on the pattern of acorn primary dispersal by the European jay in a Mediterranean landscape.
Materials and methods
Study area
We performed this study close to the village of La Pobla Tornesa, in the eastern Iberian Peninsula (40°5′N, 0°0′W, Castelló province, Valencia region, Spain). The area has a Mediterranean climate with dry summers and mild, wet winters. Two bedrock types are common, Bundsandstein sandstone (the most common) and Palaeolithic slates. The vegetation is a typical Mediterranean mosaic of shrubs, pines, abandoned fields of different ages and fields still in use. Pines (Pinus halepensis and P. pinaster) are the dominant trees in the area. Oaks (Holm oak, Quercus ilex subsp. ballota and cork oak Q. suber) are present in low densities (dispersed or in small groups).
Acorn processing and tracking
We used acorns from Q. ilex subsp. ballota and Q. suber obtained from nearby oak populations and conserved in the refrigerator. Sound acorns were visually selected, opened and partially emptied, and a transmitter (model TW-4; Biotrack, Wareham, Dorset, UK; weight: 2.2 g; mean life span: 3 months) was inserted longitudinally in each acorn, with the antenna rolled up inside (tagged acorns). The acorn was then closed and sealed (superglue 3; Loctite, Warrington, Pa.). The length, diameter (0.1 mm) and weight (0.5 g) of each tagged acorn were recorded. Because jays cannot detect 10% weight variations in acorns (Bossema 1979), no better weight estimation was necessary. Due to size limitations, we introduced the transmitter into relatively large (>5 g) acorns. We did not use smaller transmitters because of their limited battery life-span.
In autumn 2004, we set up three feeders (feeders 1–3) to form a triangle with inter-feeder distances of between 210 and 225 m. Each feeder consisted of a 40 × 40-cm seed tray mounted at the top of a 1.7-m pole. We performed the acorn tracking experiment on these feeders in early spring (March–April 2005) and early winter (December 2005–January 2006). Two additional feeders (feeders 4–5) were located more than 2 km away from feeders 1–3 (i.e. used by different jays) and set up in the spring of 2006 (March–April). All feeders were located in relatively flat areas with gentle slopes. We did not perform the experiments in the autumn because jays did not visit our feeders when natural acorns were available. In each feeder we included about one tagged acorn for every nine untagged acorns to reduce the number of tagged acorns rejected by jays. Acorn location was carried out by radio-tracking (Mariner 57 portable receiver; Biotrack), and for each retrieved acorn we recorded the Global Positioning System position (GPS 76S; Garmin Ltd, George Town, Cayman Islands) and the surrounding vegetation type (fields, recently abandoned fields, old-fields, shrubland, pine forest and roads or forest paths). We recorded the status of each recovered acorn as: (1) cached – acorn buried by jays; (2) opened – acorn found open (or only remains of the tagged acorn found), (3) abandoned – tagged acorn abandoned (probably because the jay detected the transmitter) on the soil surface or in a tree branch. Abandoned acorns were not considered any further in the analysis.
Data analysis
Differences in retrieval time between opened and cached acorns were compared by means of the Mann–Whitney U (MWU)-test as the data were not normally distributed. Dispersal distance distribution was tested against four models (normal, log-normal, Weibull and exponential) using the maximum likelihood approach implemented in MASS (Venables and Ripley 2002); Akaike Information Criteria (AIC) was used to compare the fits. In addition, the Kolmogorov–Smirnov test (KS) was also used to compare the four fitted distributions with the four theoretical distributions. We compared both the distances (using the t-test) and the distance distributions (using the KS test) between different acorn fates (cached vs. opened), different dispersal season (spring vs. late winter), different acorn species (Q. ilex and Q. suber) and different feeders. For distance comparisons, we used log-transformed as the log-normal model was the best to fit the data (see Results).
We performed a density analysis with the spatial analysis module of ArcView ver. 3.2 using a search radius of 17.5 m and a cell unit of 1 m. This allowed us to develop the kernel shadows and, afterwards, to visually identify all cache clusters – i.e. areas with high cache density. All spatial analyses were performed on each feeder independently, except for feeder 5 which had very few jay visits, thus precluding any analysis.
The vegetation composition within a 1-km radius of each feeder was obtained from the 2003 aerial photograph archives, and the necessary corrections were made during fieldwork (2005). The data were subsequently transferred to a GIS for analysis. Vegetation categories were aggregated into six vegetation types (the dominant landscape units): (1) fields, (2) recently abandoned fields (abandoned fields with low plant cover), (3) oldfields (abandoned fields with high plant cover), (4) shrublands, (5) pine forest and (6) forest roads and tracks. We tested whether the number of acorns found in each vegetation type was different than that expected from the proportion of vegetation types available around each feeder using a chi-squared test. The test was performed at different distances to the feeders (0–40, 40–80 and 80–120 m). The residuals of this test were used as indicators of preference (positive residuals) or avoidance (negative residuals). For this analysis, we consider a maximum distance of 120 m, which includes most acorns (88.6%) and all cache clusters in all feeders.
Results
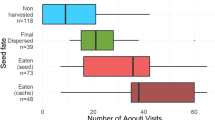
A total of 239 tagged acorns were tracked, 158 (64.0%) of which were cached; the remainder were opened by the jay (26.7%), abandoned close to the feeder (1.6%) or their radio-tracking signal was lost (4.5%). Because radio-interference in the study area was high, retrieval time varied from 1 to 93 days (mean = 5.4 days; SD = 9.9; n = 152). Signal detection was lower (usually about tens of meters) than transmitter specifications (hundreds of meters) due to the fact that the transmitter antenna was rolled up inside the acorn. Mean retrieval time – i.e. the average time between acorn placement in feeder and cache detection – tended to be shorter for opened than for cached acorns (2.8 vs. 6.6 days; P = 0.029; MWU test) because the opening of the acorn allowed the antenna to unfold to some degree (usually to its full length), thereby increasing signal detection. No differences in any of the acorn size parameters (length, width or weight) were observed between cached and opened acorns (t-test, P > 0.8), and no association between species (Q. suber vs. Q. ilex) and acorn status (cached vs. opened) was found (χ2 tests, P = 0.82). In some cases (8.3%; and only in the spring trials), the jay-cached acorns were pilfered by rodents, and the transmitters were located deep underground or inside the stone walls of field terraces. Jays tended to open more acorns in the spring (34.3%) than in the winter (22.1%; χ2 = 3.980, P = 0.046).
Dispersal distances
The distance distribution of cached acorns was right-skewed and leptokurtic (Fig. 1; skewness = 3.39, Kurtosis = 15.41). The distance distribution that best fit the data (with the lowest AIC) was the log-normal distribution (AIC = 1606.0) with a mean of 3.844 (SE = 0.068; location parameter) and a standard deviation of 0.851 (SE = 0.048; shape parameter) (values in log-scale). The other fitted dispersal distributions showed a worse performance: normal (AIC = 1,791.1), exponential (AIC = 1,637.7) and Weibull (AIC = 1,633.3). The distance distribution data were not significantly different from a log-normal distribution (KS test: W = 0.992, P = 0.516), but they differed from normal (P < 0.0001), exponential (P < 0.0001) and Weibull (P = 0.0359) distributions (KS tests). Similar results were obtained when analysing the different feeders separately, i.e. for all feeders (feeders 1–4) the best fit was the log-normal distribution, and there was no significant differences (P > 0.3, KS test) between the observed distance distributions and a log-normal distribution.

Distance distribution for cached and opened acorns (white and grey bars, respectively). The distribution of cached acorns does not differ from a log-normal distribution. Inset Shapiro–Wilk test, W = 0.99; P value = 0.55
The distributions of cached and opened acorns were significantly different (Figs. 1, 2; Table 1). The mean distance of cached acorns was 68.6 m (SD = 73.57), with values ranging from 2.8 up to about 545.4 m (25, 50, 75, 90 and 95 percentiles were 25.87, 49.15, 75.97, 149.60 and 196.71 m, respectively). Opened acorns were moved over much shorter distances, from 2.2 up to 137.5 m, with a mean of 37.6 m (SD = 34.1; 25, 50 and 75 percentiles were 11.50, 30.61 and 46.15, respectively; n = 66). No differences in either the mean or the distribution of distances were found between Q. suber and Q. ilex (Table 1). The mean distance did not differ between spring and winter (89.6 vs. 66.0 m; P = 0.152), but the distance distribution did show differences (median = 63.5 vs. 48.1 m, respectively; Table 1). There were also differences in mean distance and distribution among feeders, mostly because of shorter distances at feeders 2 and 4 (Table 2). None of the feeders showed a positive relation between the order that the acorns were cached and the distance to the caching site. Acorn size (length, diameter or weight) were not correlated with dispersal distance (Pearson’s correlation, n = 220, P > 0.6).

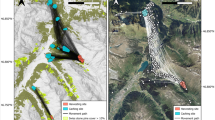
Spatial location of the three feeders that are close together, with straight lines from each cache to its corresponding feeder
Seed shadow
Feeders 1–3 were located close together. Although most (89.4%) acorns were found closer to the source feeder than to the other feeders, 10.6% showed the opposite pattern, evidencing the overlap between seed shadows (Fig. 2).
Density analysis revealed from one to five high-density areas of cached acorns close to each feeder (Fig. 3), with the exception of feeder 5. These caches clusters account for 60 (feeder 1) to 89% (feeder 3) of the total cached acorns, but their areas range from only 3 (feeder 1) to 19% (feeder 3) of the total area covered by all the acorns cached at each feeder (Table 2). Cache density at the cluster ranged between 19.5 and 75.0 (mean: 60.9) tagged acorns/ha. Extrapolation of these data to all the acorns left at the feeder (tagged and untagged acorns) suggest that cluster densities raged between 174 and 675 (mean: 419.8) acorns/ha (corrected densities in Table 2). At feeder 5, no such clusters appeared, as this feeder had very few jay visits (no spatial analysis was performed for this feeder). Isolated caches were found farther away than clusters of caches for all the feeders (KS test P < 0.01; Table 2).

Map of the cached acorns (dots) at feeders 1–4. The dimensions of each quadrat are 120 × 120 m, and thus some isolated caches fall outside the map (see distances in Table 1). Black lines delimit the clusters of caches (i.e. zones of acorn hoarding with >27 tagged acorns/ha), white crosses represent feeder position. Vegetation type legend: white fields, pattern of vertical lines recently abandoned fields (i.e. with low plant cover), pattern of horizontal lines long-abandoned fields (i.e. with high plant cover), dark grey shrubland, circle pattern pine forest, white corridors field roads and tracks
Jays did not randomly select vegetation types for hoarding acorns; we detected a trend in their vegetation-type selection (χ2 test for 0–40 m, P = 0.080; for 40–80 m, P = 0.032; for 80–120 m, P = 0.054). They preferred recent oldfields and forest tracks and avoided shrublands, irrespective of distance (Fig. 4). For fields, oldfields and pine woodlands, we found an interaction with distance in such a way that fields and oldfields were preferred at short distances and avoided at greater distances, while the opposite was true for pine woodlands (avoided at short distances and preferred at greater distances).

Residuals of the chi-squared test for each vegetation type at three distances from the feeder (P values are 0.080, 0.032 and 0.054 for 0–40, 40–80 and 80–120 m, respectively). Positive values indicate preference, negative values indicate avoidance
Discussion
Acorn dispersal by jays
Jay caches were located at distances of approximately 3 m up to approximately 550 m from feeders, often aggregated in preferred areas (cache clusters). These primary dispersal distances are shorter than those previously reported by Gómez (2003) probably because of the different landscape structure. In fact, we found significant differences between our feeders in terms of distance distribution, vegetation selection and number and distribution of preferred areas for caching acorns. Feeders 1 and 4 were in pine woodland patches with no recent oldfields (preferred patches to hoard) nearby. Thus, jays cached the acorns in the same pine woodland, preferentially on paths or forest roads. The preference for recent oldfields can be clearly seen in the data from feeder 2, where many of the other vegetation classes were in the vicinity. In contrast, feeder 3 was located inside a non-preferred habitat (oldfields and shrublands), and acorns were cached in cleared areas of the shrub patch or in medium distance small woodlands (Fig. 3). Thus, we suggest that jay dispersal-distances and seed-shadows are closely related to landscape composition and configuration and that the reason for this may be that landscape structure determines jay territory composition and use. Altogether, our findings suggest that more attention should be devoted to gaining an understanding of the role of landscape composition, configuration (Kollmann and Schneider 1997) and structures (Pausas et al. 2006a) in seed dispersal studies.
The dispersal distribution follows a log-normal distribution, which suggests the importance of the “skip distance” (Greene et al. 2004) – i.e. that the maximum seed rain is not at the seed source but a few meters away (Fig. 1). Similar results have been observed in many species, and include both wind- (Greene et al. 2004) and animal- (Russo et al. 2006) dispersed seeds. This distance distribution may represent a generalised dispersal curve for one dimension (distance). However, when the two spatial dimensions were considered, we found two clearly defined cached-acorn spatial arrangements (clusters vs. isolated scattered caches; Howe 1989, Russo et al. 2006), from which it may be possible to derive important consequences for spatial models of oak regeneration (Ribbens et al. 1994; Pausas 2006; Purves et al. 2007) and for future developments of afforestation techniques based on facilitating natural regeneration (Pausas et al. 2004; Vallejo et al. 2006). The fact that the dispersal distances did not increase with the order that the acorns were cached suggests that at the dispersal densities inferred at the cache clusters, the jays had not reached a hoard saturation; in other words, our results suggest that jays are capable of caching at densities higher than 400 acorn/ha in one season in their preferred habitats (Table 2). This point needs further testing as it is possible that these cache clusters will be recached further by jays (reallocations after “rapid sequestration”; Jenkins et al. 1995).
Jays preferred caching acorns in open habitats (recently abandoned fields, forest tracks); this preference progressively decreased as soil cover increased (oldfields, shrublands), which is in agreement with previously reported observations on jay seed caching behaviour (Bosssema 1979; Gómez 2003). This habitat preference also concurs with the seedling regeneration densities in Mediterranean mosaic landscapes (Pausas et al. 2006b; Pons and Pausas 2006). Moreover, in low-density oak populations, regeneration by rodent dispersal is usually found at a very short distance from the acorn source (Xiao et al. 2005; Xiao and Zhang 2006; Pons and Pausas 2007a). Thus, we suggest that the Quercus regeneration pattern is primarily driven by jay dispersal, at least in our mosaic landscape. However, in light of the finding that our jays dispersed some acorns to shrublands but that we failed to find any significant regeneration in this habitat (Pausas et al. 2006b; Pons and Pausas 2006), some other processes could be affecting seedling densities in this habitat (e.g. high acorn post-dispersal predation by rodents in shrublands; Pons and Pausas 2007a). On the other hand, it is also possible that the regeneration process is not the same in all shrublands, as we observed some dispersal and reduced predation in Erica shrublands but not in Ulex-dominated shrublands (unpublished observations). Recently abandoned fields have low plant cover and are preferred by the jay for caching acorns. In these conditions, the acorn predation risk (by mice) is also low (Torre and Díaz 2004; Pons and Pausas 2007a). Shrublands have very high plant cover and the predation risk by mice is very high; thus, jays avoid caching in such conditions. Oldfields are in an intermediate position with respect to plant cover and are selected at short distances only (Fig. 4), probably because the robbery risk is counteracted by the low cost of caching at short distances. On the other hand, the positive relation between distance and pine woodlands preference (Fig. 4) may be driven by jays using pines as a landmark (Bossema 1979) when flying to a suitable habitat to hoard acorns.
Rodent robbery of the jay-cached acorns was detected only in the spring trials and was lower than expected (8.3% of all cached acorns). Although these data are preliminary, to our knowledge this is the first time that predation on jay-cached acorns has been evaluated, and the proposed methodology should facilitate further research on this topic. Our results suggest that jays may reduce the possibility that rodents will pilfer the acorn by caching preferentially in open habitats where rodent activity is lower (Pons and Pausas 2007a). Moreover, during the jay caching period (autumn–winter), the wood mouse (Apodemus sylvaticus, the main post-dispersal acorn predator in the study area; Pons and Pausas 2007a) is at its lowest annual population density (unpublished data). In our study we re-used the tagged acorns very shortly after caching; to obtain more reliable conclusions on post-dispersal predation it would be necessary to monitor jay-cached acorns during the natural dispersal season over longer time periods.
We found no differences in dispersal distance and fate between the two acorn species nor between the different acorn sizes, even though differences in acorn nutrient content determine differential selection by rodents (Pons and Pausas 2006) and jays (Pons and Pausas 2007b). This suggests that, after selection, jays treated all acorns in a similar way.
Methodological issues
Acorn tracking can be used to unambiguously monitor large seeds dispersed by birds over a range of tens to hundreds of metres, thus improving the quantification and delimitation of seed shadows and (especially) the fate of long-distance dispersal events, which have been underestimated in most previous studies (Cain et al. 2000). Seed radio-tracking is relatively easy and cheap, and it does not significantly change acorn weight. Our results indicate that neither acorn species nor acorn weight modify the distance that jays move acorns. Moreover, radio-tracking permits the exact location of the cache site to be determined. Acorn radio-tracking is less time-consuming than surface-searching methods (i.e. magnetic systems), especially in long-distance dispersal events. In addition, this technique is not conditioned by vegetation, topography (as in visual flight tracking) or pilfering and belowground acorn movements by rodents. However, seed-tracking remains limited by the size and life-span of current state-of-the-art transmitters and batteries, respectively, and thus only seeds of intermediate (>1 g) to large (e.g., acorns) size are feasible for use (Sone and Kohno 1996; Mack and Druliner 2003).
The proportion of opened acorns should be treated with caution as it may not be a good estimation of predation; jays may open the acorn if they detect that it is a “false acorn”. Our results also suggest that the season of the experimental set-up may have some effect on dispersal distance. A slight reduction in acorn dispersal distances occurs between acorns cached in December–January (towards the end of the winter period) and those cached in February–April (beginning of the reproductive period; Table 1). This may be mediated by a reduction in jay territory size towards the reproductive season (Rolando 1998), which suggests a movement of acorns towards the reproductive core area. Consequently, the dispersal distances obtained in this study (especially those corresponding to spring) can be considered to be slight overestimations in comparison with those expected during the natural dispersal period (September–December).
The seed radio-tracking approach has a great potential for providing insights into acorn reallocations by jays and pilfering by rodents. The usual methods for estimating post-dispersal predation are based on artificially sown seeds in the different habitats (Gómez 2004); however, these experiments do not resemble real acorn dispersal shadows. Radio-tracking of the dispersed seeds permits marking the place and following the acorn fate in a “non-invasive” way, thus facilitating a more true assessment of post-dispersal predation. If the tagged seed is left in the field and its position tested periodically, this methodology would also allow us to study seed relocations and secondary dispersal (Van der Wall 2002).
Nonetheless, the seed radio-tracking method has shortcomings; the most important of these is that the experiments need to be performed outside the peak acorn season to force the jay to take acorns from the artificial feeders. However, improvements in the methodology may allow tracking acorns even in autumn, i.e., by inter-mixing the marked acorns with the naturally available acorns. This seems advisable as the size of the jay territory decreases towards the reproductive season (Rolando 1998) and some changes in caching behaviour could occur (Clayton 1996).
Concluding remarks
We report here for the first evidence of a spatially explicit seed shadow for oaks. This application of a seed radio-tracking method allowed us to obtain realistic acorn distributions and densities as well as jay preferences for caching acorns. Because jays selected some vegetation types and the configuration of the vegetation patches was diverse, jay dispersal distances varied greatly with the landscape context. Based on our results, variations in the jay dispersal curve among landscapes deserve future research. The methodology used here may be useful both to evaluate dispersal curves in different conditions and to elucidate some of the open questions in acorn dispersal ecology.
References
Bossema I (1979) Jays and oaks: an eco-ethological study of a symbiosis. Behaviour 70:1–117
Cain ML, Milligan BG, Strand AE (2000) Long-distance seed dispersal in plant populations. Am J Bot 87:1217–1227
Clayton NS, Mellor R, Jackson A (1996) Seasonal patterns of food storing in the jay Garrulus glandarius. Ibis 138:250–255
Darley-Hill S, Johnson WC (1981) Acorn dispersal by blue jays (Cyanocitta cristata). Oecologia 50:231–232
Dow BD, Ashley MV (1996) Microsatellite analysis of seed dispersal and parentage of saplings in bur oak, Quercus macrocarpa. Mol Ecol 5:615–627
Forget PM, Wenny DG (2005) How to elucidate seed fate? A review of methods used to study seed removal and secondary seed dispersal. In: Forget PM, Lamber JE, Hulme PE, Vander Wall SB (eds) Seed Fate: predation, dispersal and seedling establishment, CAB Int, Wallingford, pp 379–394
Godoy JA, Jordano P (2001) Seed dispersal by animals: exact identification of source trees with endocarp DNA microsatellites. Mol Ecol 10:2275–2283
Gómez JM (2003) Spatial patterns in long-distance dispersal of Quercus ilex acorns by jays in a heterogeneous landscape. Ecography 26:573–584
Gómez JM (2004) Importance of microhabitat and acorn burial on Quercus ilex early recruitment: non-additive effects on multiple demographic processes. Plant Ecol 172:287–297
Greene DF, Canham CD, Coates KD, LePage PT (2004) An evaluation of alternative dispersal functions for trees. J Ecol 92:758–766
Grivet D, Smouse PE, Sork VL (2005) A novel approach to an old problem: tracking dispersed seeds. Mol Ecol 14:3585–3595
Hardesty BD, Dick CW, Kremer A, Hubbell S, Bermingham E (2005) Spatial genetic structure of Simarouba amara Aubl (Simaroubaceae), a dioecious, animal-dispersed Neotropical tree, on Barro Colorado Island, Panama. Heredity 95:290–297
Herrera CM (1995) Plant-vertebrate seed dispersal systems in the mediterranean ecological, evolutionary, and historical determinants. Annu Rev Ecol Syst 26:705–727
Howe HF (1989) Scatter and clump-dispersal and seedling demography: hypothesis and implications. Oecologia 79:417–426
Iida S (1996) Quantitative analysis of acorn transportation by rodents using magnetic locator. Vegetation 124:39–43
Jenkins SH, Rothstein A, Green WCH (1995) Food hoarding by Merriam's kangaroo rats: a test of alternative hypotheses. Ecology 76:2470–2481
Kollmann J, Schneider B (1997) Effects of landscape structure on seed dispersal of fleshy-fruited species along forest edges. Bull Geobot Inst ETH 63:77–86
Levey DJ, Sargent S (2000) A simple method for tracking vertebrate-dispersed seeds. Ecology 81:267–274
Levey DJ, Bolker BM, Tewksbury JJ, Sargent S, Haddad NM (2005) Effects of landscape corridors on seed dispersal by birds. Science 309:146–148
Mack AL, Druliner G (2003) A non-intrusive method for measuring movements and seed dispersal in cassowaries. J Field Ornithol 74:193–196
Nathan R (2006) Long-distance dispersal of plants. Science 313:786–788
Nathan R, Perry G, Cronin JT, Strand AE, Cain ML (2003) Methods for estimating long-distance dispersal. Oikos 103:261–273
Pausas JG (2006) Simulating mediterranean landscape pattern and vegetation dynamics under different fire regimes. Plant Ecol 187:249–259
Pausas JG, Bladé C, Valdecantos A, Seva JP, Fuentes D, Alloza JA, Vilagrosa A, Bautista S, Cortina J, Vallejo R (2004) Pines and oaks in the restoration of mediterranean landscapes in Spain: new perspectives for an old practice – a review. Plant Ecol 171:209–220
Pausas JG, Bonet A, Maestre FT, Climent A (2006a) The role of the perch effect on the nucleation process in mediterranean semi-arid Oldfields. Acta Oecol 29:346–352
Pausas JG, Ribeiro E, Dias SG, Pons J, Beseler C (2006b) Regeneration of a marginal Cork oak (Quercus suber) forest in the eastern Iberian Peninsula. J Veg Sci 17:729–738
Pons J, Pausas JG (2006) Oak regeneration in heterogeneous landscapes: the case of fragmented Quercus suber forests in the eastern Iberian Peninsula. For Ecol Manage 231:196–204
Pons J, Pausas JG (2007a) Rodent acorn selection in a mediterranean oak landscape. Ecol Res. doi:10.1007/s11284-006-0053-5
Pons J, Pausas JG (2007b) Not only size matters: acorn selection by the European jay (Garrulus glandarius). Acta Oecol 31:353–360. doi:10.1016/j.actao.2007.01.004
Purves DW, Zavala MA, Ogle K, Prieto F, Rey Benayas JM (2007) Environmental heterogeneity, bird-mediated directed dispersal, and oak woodland dynamics in Mediterranean Spain. Ecol Monogr 77:77–97
Reiter J, Curio E, Tacud B, Urbina H, Geronimo F (2006) Tracking bat-dispersed seeds using fluorescent pigment. Biotropica 38:64–68
Ribbens E, Silander JJA, Pacala SW (1994) Seedling recruitment in forests: calibrating models to predict patterns of tree seedling dispersion. Ecology 75:1794–1806
Rolando A (1998) Factors affecting movements and home ranges in the jay (Garrulus glandarius). J Zool 246:249–257
Russo SE, Portnoy S, Augspurger CK (2006) Incorporating animal behavior into seed dispersal models: implications for seed shadows. Ecology 87:3160–3174
Sone K, Kohno A (1996) Application of radiotelemetry to the survey of acorn dispersal by Apodemus mice. Ecol Res 11:187–192
Torre I, Díaz M (2004) Small mammal abundance in mediterranean post-fire habitats: a role for depredators? Acta Oecol 25:137–143
Vallejo R, Aronson J, Pausas JG, Cortina J (2006) Mediterranean woodlands. In: van Andel J, Aronson J (eds) Restoration ecology: the new frontier. Blackwell, Oxford, pp 193–207
Vander Wall SB (2002) Secondary dispersal of Jeffrey pine seeds by rodent scatter-hoarders: the roles of pilfering, recaching and a variable environment. In: Levey DJ, Silva (eds) Seed dispersal and frugivory: ecology, evolution and conservation. CAB Int, Wallingford, pp193–208
Venables WN, Ripley BD (2002) Modern applied statistics with S, 4th edn. Springer, New York
Wilson M, Traveset A (2000) The ecology of seed dispersal. In: Fenner M (ed) Seeds: the ecology of regeneration in plant communities. CAB Int, Wallingford, pp 85–110
Xiao ZS, Zhang ZB (2006) Nut predation and dispersal of harland tanoak Lithocarpus harlandii by scatter-hoarding rodents. Acta Oecol 29:205–213
Xiao ZS, Zhang ZB, Wang YS (2004) Dispersal and germination of big and small nuts of Quercus serrata in a subtropical broad-leaved evergreen forest. For Ecol Manage 195:141–150
Xiao ZS, Zhang ZB, Wang YS (2005) Effects of seed size on dispersal distance in five rodent-dispersed fagaceous species. Acta Oecol 28:221–229
Ziegenhagen B, Liepelt S, Kuhlenkamp V, Fladung M (2003) Molecular identification of individual oak and fir trees from maternal tissues of their fruits or seeds. Trees 17:345–350
Acknowledgments
We thank P. Jordano for helpful comments on a draft version of this paper, Aina Blasco for her kind contribution to the fieldwork and J. Cortina for lending us part of the radio-tracking material. Comments from the three anonymous referees helped to improve the paper. This work has been financed by the EU project CREOAK (QLRT-2001-01594) and the Spanish project VARQUS (CGL2004-04325/BOS). The CEAM is funded by Generalitat Valenciana and Bancaixa.
Author information
Authors and Affiliations
Corresponding author
Additional information
Communicated by Fernando Valladares.
Rights and permissions
About this article
Cite this article
Pons, J., Pausas, J.G. Acorn dispersal estimated by radio-tracking. Oecologia 153, 903–911 (2007). https://doi.org/10.1007/s00442-007-0788-x
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00442-007-0788-x