
Abstract
The mechanisms regulating the build-up of invertebrate assemblages on ephemeral detritus patches are still poorly understood. Here, the daily colonization of decaying reed leaves by vagile macroinvertebrates was monitored in an brackish lake in Italy. The highly variable abundance patterns of dominant taxa were analysed by spectral and geostatistical techniques to test for nonrandomness and to further determine whether they were related to body size. Comparisons between two contrasting sites allowed an assessment of the generality of our observations. At both sites, the macroinvertebrate assemblage was dominated by three detritivorous taxa, i.e. the isopod Lekanesphaera monodi, the amphipod Microdeutopus gryllotalpa and the polychaete Neanthes caudata. Overall, their abundance patterns were characterised by short-term fluctuations of a nonrandom, autocorrelated nature. In addition, a significant covariation was observed between the average body mass of each taxon and the complexity of the respective abundance pattern, expressed by the fractal dimension D. The covariation was observed at both study sites, notwithstanding the diverging outcomes of bivariate pattern comparisons for similar-sized taxa. Our findings indicate that the size of macroinvertebrates is strongly related to the short-term dynamics of their abundance patterns on reed detritus, suggesting that the interaction between vagile consumers and ephemeral resource patches might be influenced by individual energetics. The implications of size-related constraints for the coexistence of species on decaying detrital patches are discussed.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
The importance of plant detritus to the functioning of an ecosystem is widely acknowledged (e.g. Cebrian and Lartigue 2004 and literature cited therein). Only recently, however, has the intrinsic variability characterising the detrital matrix at multiple spatial and temporal scales (the “ontogeny” of detritus, sensu Moore et al. 2004) been suggested to affect individuals, communities and whole ecological systems, ultimately promoting species coexistence in detritus-based food webs (Moore et al. 2004 and literature cited therein). Indeed, several studies have demonstrated how detritus heterogeneity, in terms of both availability and quality, influences the structure of the associated microheterotrophic community (e.g. Rossi et al. 1983; Bärlocher 1992; Mille-Lindblom and Tranvik 2003; Moore et al. 2004; Nikolcheva and Bärlocher 2005). On the other hand, several hypotheses have been formulated concerning the role of detritus ontogeny in the coexistence of invertebrate detritivores, ranging from facilitation (the “shredder–collector facilitation hypothesis”; Heard and Buchanan 2004 and literature cited therein), to classic niche partitioning across a mosaic of microfungal resource patches (Rossi 1985; Basset and Rossi 1990; Maraun et al. 2003 and literature cited therein). To date, however, there has been an apparent lack of a general corpus of experimental evidence on the mechanisms determining the coexistence of invertebrate species on ephemeral detritus patches.
Among other factors, differences in body size have long been considered crucial to promoting intra- and interspecific coexistence in natural communities (e.g. Hutchinson 1959; MacArthur 1972; Levinton 1982; Basset 1995 and literature cited therein). Early discussions focussed on the concept of niche partitioning, implying that coexisting organisms of different sizes interact through the use of a static, coarse-grained mosaic of different resources. In the last two decades, however, optimal foraging theories have been progressively incorporated into the original niche framework (e.g. see Basset 1995 and literature cited therein; Tokeshi 1999), converging within a hierarchical, spatially explicit perspective in which body size has maintained its centrality, since coexistence results from the interaction of the spatiotemporal patchiness of the habitat in which vagile species interact with their sizes and related characteristics (e.g. dispersal; Tilman and Kareiva 1997; Tokeshi 1999). An increasing body of literature on terrestrial vertebrate and invertebrate assemblages is providing experimental support for this theoretical framework (Brown and Bowers 1984; Fierer and Kotler 2000 and literature cited therein; Yu et al. 2004). For aquatic benthic habitats, on the other hand, the available information is limited to stream benthic systems and emphasises the interplay of body size with abiotic factors such as water flow and the physical complexity of the interstitial environment in regulating species’ coexistence within invertebrate communities (Schmid et al. 2000 and literature cited therein; Schmid et al. 2002; Taniguchi and Tokeshi 2004).
In the present study, we monitored the build-up of an assemblage of benthic invertebrate taxa on decaying patches of reed leaf detritus. The investigation was carried out in Lake Alimini Grande, a brackish lake in Puglia (SE Italy) at daily temporal resolution for 40 days. The main objective was to explore the fine-scale temporal dynamics of reed detritus colonization by vagile macrozoobenthos and to identify whether the body size of dominant invertebrate taxa was related to their colonization patterns and coexistence on detritus patches. The simple null hypothesis (H0) is for invertebrates aggregating on detrital patches to show no predictable short-term responses to the synergistic interplay of abiotic and biotic constraints characterising the benthic environment. Rejection of H0 takes two different forms: if the size of each taxon is crucial to determining its pattern of abundance variation, then a constant relationship is expected, unaffected by either density-independent (i.e. the abiotic characteristics of the benthic environment) or density-dependent (e.g. population abundance) site-specific constraints (H1). Conversely, if patterns are not size-determined, the effect is expected to vary in response to site-specific ecological constraints (H2). To discriminate between the two alternative hypotheses (H1 versus H2), the study was carried out at two experimental sites with contrasting chemical–physical characteristics of the water column and differences in the structure of the macrobenthic assemblage.
Materials and methods
Study sites and experimental set-up
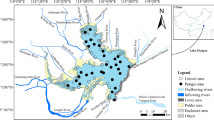
The study was carried out in Lake Alimini Grande, a brackish coastal lake located in Puglia (SE Italy) on the Adriatic Sea. The lake is ∼3 km long, with an approximate surface area of 1.26 km2 and a maximum depth of 3.5 m. Continuous water exchange with the sea is provided by a 10-m wide channel, although episodic interruptions of the water flow do occur due to siltation events. Freshwater inputs in the NW area of the basin are provided by the Zuddeo Channel and other ephemeral drainage ditches and underwater springs. Lake Alimini Piccolo, a water body connected to the main basin by a 1,300-m-long channel, represents the main freshwater source in the southern area. The riparian vegetation is dominated by the reed Phragmites australis (Cav.) Trin. ex Steudel, although Cladium mariscus (L.) Pohl, Scirpus holoschoenus L. and Cyperus longus L. can be locally abundant.
The study was undertaken in autumn 1998 at an experimental site (site “S1” hereafter) located on the NW coast of the lake near the mouth of the Zuddeo Channel. A second experimental site (site “S2”) was located on the NE coast of the lake. Notwithstanding the closeness of the two sites (∼400 m linear distance), freshwater inputs within the S1 area result in peculiar chemical–physical conditions for the shallow littoral waters compared to S2 (see Mancinelli et al. 2005 for chemical–physical data referring to the period covering the present investigation), which affects the structure of the macrobenthic community and, ultimately, reed detritus processing rates (Sangiorgio 2004; Mancinelli et al. 2005).
Senescing leaves of P. australis were collected during winter 1997 and air-dried. Blades were cut into 10-cm-long fragments; only the central section of the blade was used to minimise interpack heterogeneity of the leaf material. Aliquots of leaf fragments were weighed after drying at 60 °C for at least 72 h (“leaf packs” hereafter: 3.000±0.001 g initial dry mass) and enclosed in ballasted net bags (0.5 cm mesh size) to facilitate their manipulation and retrieval.
A total of 160 leaf packs were placed at each experimental site on 17 October 1998. Leaf packs were randomly deployed close to the reed stands within a ∼20-m long, 1-m-wide area at a water depth of ∼0.5 m. Previous observations (F. Sangiorgio, personal communication) indicated that leaf material accumulates within this area soon after shedding in a mosaic of detritus patches in response to the morphology of the bottom.
Sampling and laboratory procedures
Four leaf packs per site were collected haphazardly along the deployment area four days after the start of the experiment; a boat was used to avoid disturbances during retrieval operations. Pack collection was carried out daily for the subsequent 39 days, adopting the same sampling criteria. In the field, each leaf pack was carefully enclosed underwater in a plastic bag and subsequently transferred at 4 °C to the laboratory, where it was washed with tap water to remove organic and inorganic particles together with invertebrates. Sessile macrofauna were removed by hand and collected. Leaf fragments were dried (60 °C for at least 72 h) and weighed to the nearest mg (dry mass, DM hereafter). Invertebrates were identified to the lowest taxon necessary for trophism determination and enumerated. The DM (60 °C, >72 h) of each individual was measured to the nearest μg.
Data analysis
When not stated otherwise, values in the text are expressed as mean ± 1 SE with n=4 replicates. For univariate parametric analyses, the assumption of homogeneity of variances was checked using Cochran’s C-test, and log-transformations were performed when necessary. Statistical significance was always evaluated at α=0.05.
A repeated-measures ANOVA (RM-ANOVA hereafter) with “site” as a fixed factor and “time” as a repeated-measures factor was performed to test for intersite differences in daily estimations of leaf pack DM. The assumption of sphericity was tested using Mauchly’s test, and violations were corrected by a Huynh–Feldt adjustment when necessary (von Ende 1993). Since a significant “site” effect was observed (see “Results”), the following analyses were carried out on daily abundance data expressed as the number of individuals g−1 leaf pack DM.
To verify the general null hypothesis H0, RM-ANOVAs were used to test within each site for overall time effects on daily abundances of vagile taxa characterised by a “quantitative dominance” (QDN)>5%. The significance of the F statistics obtained was tested using a randomization procedure (Manly 1991). In brief, Monte Carlo simulations were run, repeating for each taxon the RM-ANOVA on 9999 random replications of the original data set under the null hypothesis of a completely random distribution of the total number of collected specimens over sampling times and replicates. P-values were calculated by comparing the F statistics from the original data with the empirical distributions of the generated F statistics.
Subsequently, the Kolmogorov–Smirnov d test (K–S test hereafter) was performed on abundance patterns to determine if nonrandom periodicity was present across the 40-day experimental period. Semi-variance analysis was used to investigate the autocorrelative structure of invertebrate abundance variations over time. To this end, the daily mean abundance of each invertebrate taxon was treated as a temporally referenced regionalised variable Z(X) (Legendre and Legendre 1998); the one-dimensional semi-variogram was calculated as:
where γ(k) = semi-variance at time lag k, N(k) = number of data points in the record, Z(X i ) = taxon abundance at time X i , and Z(X j ) = taxon abundance at time j, which is within a lag k of time i. Semi-variances were determined over a maximum time lag of 20 days; standardization by the variance of the data was performed to allow for further intertaxon comparisons. Drift effects were checked through visual examination of semi-variograms, and were ignored when the time lag at which the effect became significant (indicated by a near-parabolic increase in semi-variances) was greater than 20 days. The model characterised by the highest coefficient of correlation r was fitted to semi-variograms. In addition, to investigate joint interspecific variability in taxon abundances, cross semi-variograms were calculated as:
where X and Y are the abundances of two taxa; further notation as in Eq. 1. Since the scope of the analysis was to detect bivariate relationships between data series, only linear models were fitted to cross semi-variograms. To avoid underestimating cross semi-variances at the shortest time lags, zero pairs in data series were not included in the analyses.
To discriminate between hypotheses H1 and H2, Welch t-tests were used to test for differences in the mean size of the dominant taxa; differences in the size distributions were verified using chi-square tests on individual mass data pooled over the experimental period and allocated to a geometric (×2) mass scale of ten size classes (0–0.02 to 5.12–10.24 mg). Size distributions were used to calculate standard descriptive parameters (i.e. mean, median, mode, maximum and minimum, 25th and 75th percentiles). Finally, semi-variograms were used to determine the fractal dimension D of the abundance pattern of each taxon, providing an indirect yet quantitative estimation of pattern complexity in the time domain. Fractal distributions are characterised by the equation:
corresponding to a power model without a sill. In the equation, H is defined as the Hurst exponent, related to the fractal dimension D by the relationship:
(Burrough 1981), where β is the slope of the time lag (h) versus variogram γ(h) plotted on a log–log scale. The experimental semi-variograms were log–log-transformed and fitted via least squares by a no-sill power law model. Slopes were preventively compared among taxa using ANCOVA procedures followed by planned orthogonal contrasts, and the fractal dimension was calculated according to Eq. 3. Parametric regression analyses were performed against the total abundances of taxa and standard descriptive parameters of the size distributions to identify the best predictor of D.
Results
The composition of the leaf pack invertebrate assemblage
During the study period, 1,144 individuals belonging to 13 vagile invertebrate taxa and 2,744 specimens belonging to two sessile taxa were collected on leaf packs. In particular, ∼90% of the vagile macrofauna at both experimental sites were made up of three detritivore species individually characterised by a QDN>5%, i.e. the isopod Lekanesphaera monodi, the amphipod Microdeutopus gryllotalpa and the polychaete Neanthes caudata (Table 1). L. monodi was the most abundant taxon at site S1, while M. gryllotalpa was twice as abundant as the isopod at the other study site. The numerical contribution of N. caudata was low, although a higher abundance of the polychaete was observed at site S2 compared to site S1 (Table 1). Other detritivores not considered further were the amphipod Corophium insidiosum and the gastropod Bittium reticulatum (cumulative QDN=7.78%).
The sessile macrofauna found on leaf packs were dominated by Balanus sp. At site S2, barnacles were collected from day 10, reaching a maximum abundance of 177 individuals per leaf pack at day 41. Conversely, at site S1 only 67 barnacles were collected in total and were not included in the following analyses. Similarly, the other sessile taxon, Mytilus sp., was characterised by negligible occurrence (11 and six specimens, sites S1 and S2, respectively) and was not considered in the following analyses.
Size of dominant vagile taxa
Overall, L. monodi was the largest vagile taxa (0.97±0.07 mg mean individual DM, n=403), followed by M. gryllotalpa (0.53±0.02 mg, n=485) and N. caudata (0.33±0.03 mg, n=140). This size-based hierarchy was confirmed at both experimental sites, where significant interspecific differences in average body size (individual DM: Welch t test, P≤0.001 for all pairwise comparisons) and size–frequency distribution were observed; noticeably, size–frequency distributions were characterised by a higher interspecific similarity at site S1 than at site S2 (Table 2).
Specifically, L. monodi individuals collected at site S1 were significantly smaller than those collected at site S2 (0.82±0.07 mg vs. 1.27±0.14 mg mean individual DM; Welch t test: t=2.89, P=0.004 for 205.9 approx. df), and significant intersite differences in the size–frequency distribution of individual DM were observed (Fig. 1). In contrast, M. gryllotalpa did not show significant intersite differences in body mass, either on average (0.52±0.02 mg vs. 0.53±0.02 mg mean individual DM at sites S1 and S2, respectively; Welch t test: t=0.28, P=0.78 for 466.9 approx. df) or in terms of frequency distribution (Fig. 1). Similarly, N. caudata was characterised by marginally nonsignificant intersite variation in average body mass (site S1: 0.27±0.02 mg; site S2: 0.36±0.04 mg; Welch t test: t=1.95, P=0.053 for 127.9 approx. df), and negligible differences in the size–frequency distributions (Fig. 1).

Size–frequency distributions of Lekanesphaera monodi, Microdeutopus gryllotalpa and Neanthes caudata at sites S1 (filled rectangles) and S2 (open rectangles). Text in the graphs refers to results of intersite chi-square tests
Mass loss of detrital patches and colonization by invertebrates
After 43 days of immersion, leaf packs lost more than one-third of their initial mass at both experimental sites. In particular, decay was significantly slower at site S2 compared to site S1 (1.92±0.04 g vs. 1.72±0.03 g final DM), and a significant difference was observed for the DM remaining in leaf packs (RM-ANOVA, factor “site”: F (1,6)=21.69, P=0.003; interaction factor “site × time”: F (39,234)=1.37, P=0.08).
Overall, leaf pack colonization patterns of L. monodi, M. gryllotalpa and N. caudata were characterised by intense short-term fluctuations throughout the experimental period (Fig. 2). Yet, RM-ANOVAs followed by complete randomizations of individual occurrence among sampling times and replicates indicated that the observed variations were significantly different from those expected by chance (Table 1). Moreover, these variations were characterised by a nonrandom structure (K–S test: P always <0.05 for all combinations species/sites). In contrast, the sessile Balanus sp. at site S2 was characterised by an almost monotonic abundance variation over time (Fig. 3), with random daily fluctuations (K–S test, d=0.13, P>0.05).

Abundance patterns (expressed as N individuals g−1 leaf pack DM) of Lekanesphaera monodi (filled circles), Microdeutopus gryllotalpa (open circles) and Neanthes caudata (open squares) at the two experimental sites. Bars = ±1 SE

Abundance pattern (expressed as N individuals g−1 leaf pack DM) of the sessile barnacle Balanus sp. at site S2. Bars = ±1 SE. The inset shows the semi-variogram of temporal abundance variation calculated for the barnacle
Without exception, a substantial portion of the abundance variation of L. monodi, M. gryllotalpa and N. caudata was time-dependent (semi-variogram analysis; Table 3). The occurrence of negligible relationships at an intersite level was highlighted for each taxon by further cross semi-variance analyses (P always >0.05 for all comparisons). On the other hand, interspecific comparisons between M. gryllotalpa and both L. monodi and N. caudata abundances attested to the occurrence at site S1 of negative relationships, while at site S2 the relationships were positive (Table 3). In addition, L. monodi and N. caudata abundances were positively related at both sites.
Balanus sp. did not show any autocorrelated structure, besides an exceptionally strong drift effect occurring at a time lag of >4 days (see inset in Fig. 3).
Complexity of abundance patterns and relationships with body size
Significant interspecies differences were observed among the slopes of linearised power models fitted to log–log transformed semi-variograms (Fig. 4; contrast analysis after significant type-III ANCOVA interactions: P always <0.05 for all paired comparisons). The fractal dimension D computed from the slope data was characterised by maximum values for L. monodi and minimum values for N. caudata, while for M. gryllotalpa intermediate values were determined (Table 4). This pattern was observed at both sites (Table 4), and corresponded to a positive relationship between body mass parameters of taxa and the fractal Ds of their abundance patterns (Fig. 5). Further bivariate comparisons with total abundance data and the other size-related descriptive parameters indicated that mean individual DM was the only parameter significantly related to D (site S1: max r = 0.986 for 25th percentile; site S2: max r = 0.956 for 75th percentile; P=0.106 and P=0.189, respectively; one df for all correlations).

Semi-variograms of temporal abundance variation calculated for Lekanesphaera monodi (filled circles), Microdeutopus gryllotalpa (open circles) and Neanthes caudata (open squares) at the two lake sites, fitted via least squares by the power function log(semi-variance/sample variance) = α + β log(time lag) on a log–log scale. The corresponding slopes β (±1 SE) and coefficients of regression r of the power models are reported. ***P<0.001

Mean individual body mass (expressed as mg DM; open squares) of Lekanesphaera monodi, Microdeutopus gryllotalpa and Neanthes caudata and the fractal dimension D calculated for their respective abundance patterns at sites S1 (top) and S2 (bottom). Box: ±1 SE for both axes. The median values (full circles), 25th and 75th percentiles (whiskers) and minimums and maximums (asterisks and numbers within the graphs) of body mass distributions are shown
Discussion
In our study, the vagile macrobenthos colonising reed leaf packs in Lake Alimini was dominated by three invertebrate taxa whose abundance patterns showed strong short-term fluctuations. These variations were demonstrated to be of a nonrandom, autocorrelated nature, disproving the original null hypothesis H0. Furthermore, whereas site-differentiated bivariate relationships were observed among coexisting taxa, the complexity of their abundance patterns was positively related to the respective individual average mass at both experimental sites, fulfilling the criteria for the alternative hypothesis H1, but not supporting hypothesis H2.
Our study is based on an assumption of ergodicity (sensu Herben et al. 2000), i.e. at each site the 160 leaf packs prepared behaved so homogeneously during the experimental period that the average parameters calculated for packs at each sampling time always equalled the average that could have been calculated for the remaining packs.
Leaf packs were carefully manipulated during deployment and retrieval operations to avoid bias in attractiveness and artificial increases in the abundance variability that naturally occurs among replicates. In addition, the results of both RM-ANOVAs followed by full randomization of specimen occurrence, together with K–S test results, provide sound a posteriori support for the assumption. The fulfilment of the assumption of ergodicity in our experimental system means that the abundance fluctuations for the dominant taxa could not have been determined by simple interpack stochastic variability, and this poses two key questions: what caused the nonrandom patterns of variation in the abundance of invertebrate taxa in the benthic environment of Lake Alimini, and what are the implications of the site-invariant relationship between the complexity of abundance patterns and body size?
Nonrandom abundance patterns and the coexistence of taxa on detrital patches
The nonrandomicity of the abundance variations of taxa, besides disproving the original null hypothesis H0, implies deterministic constraints on the structuring of the macrobenthic assemblage, such as predation, passive dispersion, or active movement (Levinton and Kelaher 2004 and literature cited therein). Indeed, the negligible abundance of invertebrate predators inside the leaf packs excludes direct (consumption) or indirect (escape responses) predatory effects. Since access to leaf packs was precluded by net bags, direct effects due to predatory fish can also be excluded. Indirect influences cannot be excluded, but they are likely to have only dampened the abundance fluctuations of invertebrates, reducing their mobility across leaf patches (e.g. Wooster and Sih 1995; but see below in this section). Additionally, the intersite asynchrony in the abundance patterns of taxa excludes the influence of large-scale abiotic factors such as lateral passive advection due to tides. The effects of factors acting on a local scale (e.g. discharge variations of the Zuddeo channel at site S1) cannot be rejected in principle, but it is unlikely that they could have had the necessary periodicity to produce the observed abundance fluctuations. Thus, although it was neither directly observed or measured, active movement of individuals remains the most plausible determinant of the abundance fluctuations observed in our study, as well as their nonrandom nature. It is interesting to note in this regard that the abundance variations of the sessile barnacle were stochastic and did not show any autocorrelated structure beside a strong drift effect. Significant, in-phase coherence between nonrandom abundance fluctuations of L. monodi and short-term variations in the ash-free DM of the detrital resource has been shown elsewhere (Mancinelli et al. 2005), indicating that a trophic constraint can actually determine short-term abundance patterns of invertebrate detritivores. Thus, the nonrandom autocorrelated abundance variations may have been determined, at least for L. monodi, by specific responses to microhabitat differences in trophic attractiveness between prepared leaf packs and the natural detrital accumulations available in the benthic system of our study sites. The hypothesis implies that some factor(s) must counteract the tendency of the detrital patch to decrease its trophic attractivity due to mass loss. Heterotrophic microflora are widely recognised to be a key factor in enhancing and extending the trophic attractivity of detritus to detritivores over time (Suberkropp 1992; Graça 2001 for freshwater environments; Maraun et al. 2003 and literature cited therein for terrestrial habitats), and, more generally, in counteracting the natural ontogeny of the detrital resource towards a condition of low-quality, highly recalcitrant residue (Moore et al. 2004). Moreover, rapid compensatory after-grazing growth of the microflora has been repeatedly observed, both in aquatic benthic systems (Morrison and White 1980; Graça et al. 2000 and literature cited therein) and in terrestrial forest floors (e.g. Bengtsson et al. 1993 and literature cited therein; Hedlund and Augustsson 1995). From this perspective, considering the high spatial patchiness of detrital accumulation in benthic systems (Schmid 2000 and literature cited therein; Moore et al. 2004), the observed abundance patterns of invertebrates indirectly testified to a colonisation/emigration dynamic of consumers actively selecting or rejecting the prepared leaf packs from a mosaic of natural detrital patches in the search for favoured trophic conditions (giving-up behaviour, in Charnov 1976; Hildrew and Townsend 1982; Malmqvist 1993; Basset 1995). In addition, the observed intersite variations in the signs of cross semi-variograms calculated between similar-sized taxa provide further evidence that body size might have direct implications for the coexistence of consumers on the detrital patch, structuring the way their respective abundance patterns intertwine in the time domain.
At site S1, where differences in abundance and size between similar-sized species pairs were low compared to site S2, negative cross semi-variance relationships were observed. Conversely, at site S2, under higher interspecific differentiation in body size and abundance, cross semi-variance comparisons showed positive relationships between species pairs. Our results are fully consistent with a scenario where, at both experimental sites, a competitive hierarchy has determined the coexistence of taxa within a classical, Hutchinsonian size-based partitioning of the resource in the time domain (Hutchinson 1959), mediated by size-related optimal foraging mechanisms of patch selection and giving-up behaviour among resource patches (Basset 1995; Tokeshi 1999). Nonetheless, inferring competition from census data necessarily requires the interaction to be experimentally verified (e.g. see Pfister 1995 among others). Further laboratory and field investigations are needed to provide more complete insight into interspecific, size-dependent interactions occurring among detritivores.
Relationship between body size of invertebrate taxa and the complexity of their abundance patterns
Our investigation indicated that the complexity of abundance patterns on leaf packs was species-specific, and that for each taxon the individual average mass of individuals was the best predictor of this complexity. Each of the three dominant taxa had nonrandom abundance oscillations on the leaf packs, the largest (L. monodi), the medium-sized (M. gryllotalpa) and the smallest taxon (N. caudata) being characterised by the highest, intermediate and lowest fractal dimension D of the respective abundance patterns. This body size-related pattern emerges independently from the influence of both the density-independent (abiotic features of the benthic environment) and density-dependent (population abundances) controlling factors characterising the two study sites. This is consistent with recent hypotheses on the influence of body size in the structuring of stream invertebrate communities (Schmid et al. 2000, 2002) and in general with expectations from metabolic theory (Brown et al. 2004). According to the latter, movement velocity (Marquet et al. 2005), home range size (Peters 1983), patch encounter rates (Ritchie 1998; Haskell et al. 2002), resource depletion in the patch (Basset 1995; Ritchie 1998) and its perception (indirectly expressed by the resource giving-up density: Basset 1995 and literature cited therein) all scale with individual body size due to the allometry of metabolic activity (Peters 1983). Accordingly, in our study the large-sized L. monodi might have perceived the depletion and consequent renewal of the detrital patch earlier than the smaller M. gryllotalpa; similarly, the amphipod might have responded to patch depletion earlier than the smaller N. caudata. Aggregating on or abandoning the leaf packs at a higher rate necessarily increased the complexity of abundance patterns in time, highlighted by the higher fractal dimension D. Thus, our observation of a positive relationship between the mean body size of invertebrates and the fractal dimension of their patch use in the time domain is fully consistent with the negative relationship between body size of taxa and the path fractal dimension commonly observed spatially (e.g. insects: Wiens and Milne 1989; With 1994a, 1994b; Wiens et al. 1995; aquatic invertebrates: Schmid 2000 and literature cited therein; vertebrates: Bascompte and Vilà 1997; McDonald and St. Clair 2004; Nams 2005), since in both cases the observed patterns are strictly dependent on the size-related use of space.
In conclusion, our results suggest that body size of vagile macroinvertebrates can be a strong enough forcing constraint to overcome both density-independent and density-dependent controls on their patterns of abundance in ephemeral detrital patches. These findings imply that the metabolism of individuals, of which body size is an ultimate determinant, might represent a key factor regulating the dynamics of vagile consumer–resource interactions. Further experimental efforts are needed to provide additional support for our results and to test their generality. Specifically, highly spatially or temporally resolved field studies will provide a useful framework in which to understand the influence of body size-dependent mechanisms of resource use on species coexistence in natural communities.
References
Bärlocher F (1992) Community organization. In: Bärlocher F (ed) The ecology of aquatic hyphomycetes. Springer, Berlin Heidelberg New York, pp 38–76
Bascompte J, Vilà C (1997) Fractals and search paths in mammals. Landscape Ecol 12:213–221
Basset A (1995) Body size-related coexistence: an approach through allometric constraints on home-range use. Ecology 76:1027–1035
Basset A, Rossi L (1990) Competitive trophic niche modifications in three populations of detritivores. Funct Ecol 4:685–694
Bengtsson G, Hedlund K, Rundgren S (1993) Patchiness and compensatory growth in a fungus—Collembola system. Oecologia 93:296–302
Brown JH, Bowers MA (1984) Patterns and processes in three guilds of terrestrial vertebrates. In: Strong DR Jr, Simberloff D, Abele LG, Thistle AB (eds) Ecological communities: conceptual issues and the evidence. Princeton University Press, Princeton, NJ, pp 282–296
Brown JH, Gillooly JF, Allen AP, Savage VM, West GB (2004) Toward a metabolic theory of ecology. Ecology 85:1771–1789
Burrough PA (1981) Fractal dimensions of landscapes and other environmental data. Nature 294:240–242
Cebrian J, Lartigue J (2004) Patterns of herbivory and decomposition in aquatic and terrestrial ecosystems. Ecol Monogr 74:237–259
Charnov E (1976) Optimal foraging theory: the marginal value theorem. Theor Popul Biol 9:129–136
Fierer N, Kotler BP (2000) Evidence for micropatch partitioning and effects of boundaries on patch use in two species of gerbils. Funct Ecol 14:176–182
Graça MAS (2001) The role of invertebrates on leaf litter decomposition in streams—a review. Int Rev Gesamten Hydrobiol 86:383–393
Graça MAS, Newell SY, Kneib RT (2000) Grazing rates of organic matter and living fungal biomass of decaying Spartina alterniflora by three species of saltmarsh invertebrates. Mar Biol 136:281–289
Haskell JP, Ritchie ME, Olff H (2002) Fractal geometry predicts varying body size scaling relationships for mammal and bird home ranges. Nature 418:527–530
Heard SB, Buchanan CK (2004) Grazer-collector facilitation hypothesis supported by laboratory but not field experiments. Can J Fish Aquat Sci 61:887–897
Hedlund K, Augustsson A (1995) Effects of enchytraeid grazing on fungal growth and respiration. Soil Biol Biochem 27:905–909
Herben T, During HJ, Law R (2000) Spatio-temporal patterns in grassland communities. In: Dieckmann U, Law R, Metz JAJ (eds) The geometry of ecological interactions: simplifying spatial complexity. Cambridge University Press, Cambridge, pp 48–64
Hildrew AG, Townsend CR (1982) Predators and prey in a patchy environment: a freshwater study. J Anim Ecol 51:797–815
Hutchinson GE (1959) Homage to Santa Rosalia, or why are there so many kinds of animals? Am Nat 93:145–159
Legendre P, Legendre L (1998) Numerical ecology. Elsevier, Amsterdam
Levinton JS (1982) The body size-prey size hypothesis: the adequacy of body size as a vehicle for character displacement. Ecology 63:869–872
Levinton JS, Kelaher B (2004) Opposing organizing forces of deposit-feeding marine communities. J Exp Mar Biol Ecol 300:65–82
MacArthur RH (1972) Geographic ecology. Harper & Row, New York
Malmqvist B (1993) A comparison of activity and giving up time in two species of Rhyacophila (Trichoptera). In: Otto C (ed) Proceedings of the 7th International Symposium on Trichoptera, 1992. Buckhuhs, Leiden, pp 257–260
Mancinelli G, Sabetta L, Basset A (2005) Short-term patch dynamics of macroinvertebrate colonization on decaying reed detritus in a Mediterranean lagoon (Lake Alimini Grande, Apulia, SE Italy). Mar Biol 148:271–283
Manly BF (1991) Randomization and Monte Carlo methods in biology. Chapman & Hall, New York
Maraun M, Martens H, Migge S, Theenhaus A, Scheu S (2003) Adding to the enigma of soil animal diversity: fungal feeders and saprophagous soil invertebrates prefer similar food substrates. Eur J Soil Biol 39:85–95
Marquet PA, Quinones RA, Abades S, Labra F, Tognelli M, Arim M, Rivadeneira M (2005) Scaling and power-laws in ecological systems. J Exp Biol 208:1749–1769
McDonald WR, St. Clair CC (2004) The effects of artificial and natural barriers on the movement of small mammals in Banff National Park, Canada. Oikos 105:397–407
Mille-Lindblom C, Tranvik LJ (2003) Antagonism between bacteria and fungi on decomposing aquatic plant litter. Microb Ecol 45:173–182
Moore JC, Berlow EL, Coleman DC, de Ruiter PC, Dong Q, Hastings A, Johnson NC, McCann KS, Melville K, Morin PJ, Nadelhoffer K, Rosemond AD, Post DM, Sabo JL, Scow KM, Vanni MJ, Wall DH (2004) Detritus, trophic dynamics and biodiversity. Ecol Lett 7:584–600
Morrison SJ, White DC (1980) Effects of grazing by estuarine gammaridean amphipods on the microbiota of allochthonous detritus. Appl Environ Microbiol 40:659–671
Nams VO (2005) Using animal movement paths to measure response to spatial scale. Oecologia 143:179–188
Nikolcheva LG, Bärlocher F (2005) Seasonal and substrate preferences of fungi colonizing leaves in streams: traditional versus molecular evidence. Environ Microbiol 7:270–280
Peters RH (1983) The ecological implications of body size. Cambridge studies in ecology. Cambridge University Press, Cambridge
Pfister CA (1995) Estimating competition coefficients from census data: a test with field manipulations of tidepool fishes. Am Nat 146:271–291
Ritchie ME (1998) Scale-dependent foraging and patch choice in fractal environments. Evol Ecol 12:309–330
Rossi L (1985) Interactions between invertebrates and microfungi in freshwater ecosystems. Oikos 44:175–184
Rossi L, Fano AE, Basset A, Fanelli C, Fabri AA (1983) An experimental study of a microfungal community on plant detritus in a Mediterranean woodland stream. Mycologia 75:887–896
Sangiorgio F (2004) Ruolo della mole corporea individuale nella organizzazione di comunità a base detrito. PhD Thesis, University of Lecce, Italy
Schmid PE (2000) Fractal properties of habitat and patch structure in benthic ecosystems. Adv Ecol Res 30:339–401
Schmid PE, Tokeshi M, Schmid-Araya JM (2000) Relationship between population density and body size in stream communities. Science 289:1157–1160
Schmid PE, Tokeshi M, Schmid-Araya JM (2002) Scaling in stream communities. Proc R Soc Lond B 269:2587–2594
Suberkropp K (1992) Interactions with invertebrates. In: Bärlocher F (ed) The ecology of aquatic hyphomycetes. Springer, Berlin Heidelberg New York, pp 118–134
Taniguchi H, Tokeshi M (2004) Effects of habitat complexity on benthic assemblages in a variable environment. Freshw Biol 49:1164–1178
Tilman D, Kareiva P (1997) Spatial ecology: the role of space in population dynamics and interspecific interactions. Princeton University Press, Princeton, NJ
Tokeshi M (1999) Species coexistence: ecological and evolutionary perspectives. Blackwell, Oxford
von Ende CN (1993) Repeated-measures analysis: growth and other time-dependent measures. In: Schneider SM, Gurevitch J (eds) Design and analysis of ecological experiments. Chapman & Hall, New York, pp 113–137
Wiens JA, Crist TO, With KA, Milne BT (1995) Fractal patterns of insect movement in microlandscape mosaics. Ecology 76:663–666
Wiens JA, Milne BT (1989) Scaling of landscapes in landscape ecology, or, landscape ecology from a beetle’s perspective. Landscape Ecol 3:87–96
With KA (1994a) Ontogenic shifts in how grasshoppers interact with landscape structure—an analysis of movement patterns. Funct Ecol 8:477–485
With KA (1994b) Using fractal analysis to assess how species perceive landscape structure. Landscape Ecol 9:25–36
Wooster D, Sih A (1995) A review of the drift and activity responses of stream prey to predator presence. Oikos 73:3–8
Yu DW, Wilson HB, Frederickson ME, Palomino W, De la Colina R, Edwards DP, Balareso AA (2004) Experimental demonstration of species coexistence enabled by dispersal limitation. J Anim Ecol 73:1102–1114
Acknowledgments
We wish to thank Piervincenzo Pintozzi and Luca Tafuro for their help in field data collection, and Franca Sangiorgio for providing information on the Lake Alimini system. We are in debt to Donald DeAngelis for his comments on an advanced version of the manuscript, which was further improved by constructive suggestions provided by Peter Schmid and an anonymous reviewer. Funding from EEC project n ENV-CT97-0584, INTERREG IIIB CADSES (2000–2006), and FIRB 2003–2005 are acknowledged. All of the experiments described in the present paper comply with current Italian laws.
Author information
Authors and Affiliations
Corresponding author
Additional information
Communicated by Martin Attrill.
Rights and permissions
About this article
Cite this article
Mancinelli, G., Sabetta, L. & Basset, A. Colonization of ephemeral detrital patches by vagile macroinvertebrates in a brackish lake: a body size-related process?. Oecologia 151, 292–302 (2007). https://doi.org/10.1007/s00442-006-0586-x
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00442-006-0586-x