
Abstract
Many small passerine birds worldwide lay white eggs speckled with red, brown and black protoporphyrin pigment spots (maculation). Unlike some patterns of avian eggshell pigmentation which clearly serve a crypsis or signalling function, the ubiquity of maculation among passerines suggests that its origins lie in another function, not specific to any particular ecological or behavioural group. Elsewhere, we have presented evidence that protoporphyrin pigments serve a structural function related to eggshell thickness and calcium availability: eggshell maculation in the great tit Parus major increases with decreasing soil calcium levels, pigments demarcate thinner areas of shell, and both the pigment intensity and distribution are related to shell thickness. Here we show that maculation also affects the rate of water loss from the egg during incubation (≈ Mass Loss per Day or MLD, which is critical to egg viability), but not that of unincubated eggs. We also demonstrate, both by observation and experiment, that the effect of female incubation behaviour on MLD compensates in some way for variation in egg characteristics, and that differences between females in the degree of such compensation are related to differences in clutch maculation. Our results suggest that, while a principal function of maculation in this species may be to strengthen the eggshell, it may also reduce eggshell permeability when large amounts of pigment are used, and that this necessitates a behavioural adjustment from the female during incubation. We discuss these findings and make further testable predictions from our model.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Many small passerines lay white eggs speckled with reddish spots (the maculated egg), typically forming a ‘corona’ ring around the shoulder (Davies and Brooke 1989a, b; Lack 1968; Gosler et al. 2005). Since passerines represent c. 60% of all extant bird species (Sibley and Monroe 1990), this pattern represents one of the most common definable forms of eggshell pigmentation in birds. Eggshell pigmentation has fascinated naturalists for many decades (see, e.g., Seebohm 1896; Newton 1896), and explanations of other forms of pigmentation pattern are clearly found in terms of their providing crypsis (Bakken et al. 1978; Blanco and Bertellotti 2002; Sanchez et al. 2004), in signalling female condition (Moreno and Osorno 2003; Moreno et al. 2004; Soler et al. 2005), or in relation to brood-parasitism (Davies and Brooke 1989a, b). However, the ubiquity of the maculated egg amongst passerines varying widely in geographical range, ecology and behaviour (see Gosler et al. 2005) demands a more general hypothesis to explain its origins and function.
Elsewhere, we have presented evidence for a new paradigm to explain the origin of maculation—one based on structural function rather than visible pattern (Gosler et al. 2005). Essentially, we have suggested that pigmentation is used to compensate for localized shell-thinning (e.g. caused by calcium deficiency) that may reduce shell strength. In support of this, we have shown in a wild population of the great tit Parus major that pigment spots (maculae) mark thin areas of shell, and that maculation correlates negatively with soil calcium levels near the nest; both the pigment ‘darkness’ and ‘spread’ (reflecting intensity and distribution of pigment) are linked to the variability or patchiness in shell thickness. Apart from its effect on eggshell strength (Ar et al. 1979; Tyler 1969), eggshell thickness also critically, and inversely, affects the rate of water conductivity through the eggshell by lengthening the diffusion pathway for water vapour (Tazawa and Whittow 2000), thus affecting the rate of mass loss from the egg during incubation (Ar et al. 1974). The rate of water loss, which is typically c. 18% throughout incubation, is critical for normal embryonic development (Rahn and Ar 1974). Eggshell thickness has already been shown to co-vary with the rate of water loss, specifically in wild great tit eggs (Weimar and Schmidt 1998). However, the eggshells of small passerines may be even more permeable than might be predicted from their thickness and pore density alone. This is because, unlike the eggshells of most birds in which the ‘true eggshell’ (sensu Mikhailov 1997) consists of a number of distinct crystalline layers, that of small passerines is almost entirely ‘occupied by the continuous layer, consisting of “flaky” and well-vesiculated squamatic material’ (quote from Mikhailov 1997 p.70). Mikhailov (1997) further described this as a ‘simplified’ eggshell structure. Thus, in this kind of eggshell, the vesicles or interstices between the crystals of the calcite lattice may promote water loss by increasing the connectivity of spaces between crystals and between shell pores. This may be particularly important in thinner eggshell. The close association of pigment with thinner, and therefore more permeable, areas of shell of great tit eggs (Gosler et al. 2005) presents the possibility that pigmentation may compensate for localised shell-thinning by reducing permeability (by physically blocking the interstices between calcite crystals). Here we focus on these relationships by investigating the rates of mass loss with respect to maculation in wild great tit eggs.
The great tit is a small passerine [c. 18 g] whose eggs [c. 1.7 g], maculated with red-brown spots that often form a corona, are typical of such species worldwide. We have argued (Gosler et al. 2005) that the great tit’s hole-nesting habit, together with the facts that the female covers the clutch with nest-lining and appears unresponsive to brood parasitism (see Gosler 1993), suggest that crypsis or signalling are unlikely to explain eggshell maculation in this species. This led us to investigate possible structural functions for pigmentation (first hypothesized by Solomon 1987). Calcium availability during egg formation is potentially a critical limiting resource for eggshell formation amongst all birds. However, as this is especially so for small species, which must find calcium daily for eggshell formation (Graveland and Berends 1997; Graveland and Drent 1997; Graveland et al. 1994; Perrins 1996), our studies have focused on linking pigment maculation and structural properties of the shell—such as thickness and water conductance—with local calcium availability.
Protoporphyrins, which are the primary constituents of eggshell maculae (Kennedy and Vevers 1976), have two physical properties that could have structural consequences for maculated eggs. First, their molecular structure suggests that they might strengthen the shell by providing a solid-state lubricant that increases shell strength by acting as an intercrystalline shock-absorber (Solomon 1987, 1997). Secondly, although we offer a different mechanism by which maculation may reduce rate of water loss (see above), we note that eggshell protoporphyrins reflect strongly in the infra-red (Bakken et al. 1978). This raises the intriguing possibility that they might produce cold spots under incubation, so potentially reducing water loss. However, it is unclear whether this could have a significant effect (see Discussion) and we do not test this hypothesis here.
Although eggshell strength is largely determined by one factor—eggshell thickness (Tyler 1969; Ar et al. 1979), the rate of water loss (≈ mass loss, Ar et al. 1974) from the egg during incubation is determined by two factors: (1) The water vapour conductance of the shell and shell membranes, which is largely a function of both the thickness of the shell, and the size and number of shell pores (Tazawa and Whittow 2000), and (2) the difference in water-vapour pressure between the contents of the egg and the microenvironment in which the egg is situated (Rahn and Ar 1974; Ar and Rahn 1978; Tazawa and Whittow 2000). Thus, while the mass loss per day (MLD) from the whole egg is determined chiefly by the egg’s size (MLD increasing with surface area) and eggshell thickness (MLD decreasing with thickness), the water-vapour pressure difference is determined by a combination of microenvironmental factors including the nature of the nest and incubation ‘tightness’, and the temperature and ventilation frequency of the incubating parent (believed always to be the female in Paridae). The water-vapour pressure difference is therefore under some parental control (Ar and Rahn 1978; Tazawa and Whittow 2000). Thus, the total rate of water loss (MLD) is determined by both intrinsic properties of the egg and the external influence of the incubating female. We term the rate of mass loss due only to intrinsic factors (e.g. surface area, shell thickness) the ‘passive’ MLD, whilst that due also to female-specific factors experienced during incubation is termed the ‘active’ MLD.
If eggshell pigmentation compensates perfectly for the increased water loss that results from shell thinning, we should expect to see no correlation between MLD and any measure of eggshell pigmentation. Thus, in such an optimized system it should not be possible to measure the efficacy of pigmentation directly; rather, its effects might be inferred through its interaction with other factors that affect MLD. Although we have looked for effects of both pigment darkness and spread, we concentrate on the former because we have shown that pigment darkness is itself a measure of the extent of localized shell-thinning (Gosler et al. 2005), which should increase MLD. By measuring passive MLD in eggs collected prior to incubation to determine which factors should explain water loss from passive eggs, by then measuring active MLD in incubated eggs, and by the means of a cross-fostering experiment, we show that maculation does affect eggshell permeability, that the nest microenvironment produced by the female (e.g. through incubation behaviour) also has a profound effect on MLD, and that this too is integrated with eggshell maculation. None of these relationships would be expected from any maculation hypothesis based on signalling or crypsis.
As in Gosler et al. (2005), we studied the eggs of great tits using nestboxes in Wytham Woods near Oxford, UK. Maculation varies greatly between great tit clutches in this study area, but a common feature is a strong intra-clutch gradient from lighter to darker pigmentation through the laying sequence (see Fig. 5 in Gosler et al. 2005). As previously, nestboxes were associated with their nearest soil-calcium values, and each egg was assessed visually for pigment ‘darkness’ and ‘spread’ (see 'Materials and methods' section and Gosler et al. 2000, 2005). Pigment ‘darkness’ is known to be heritable on the female line (Gosler et al. 2000), but pigment ‘spread’ appears not to be heritable (Gosler et al. 2005).
Materials and methods
Study site
Wytham Woods (360 ha, 51°47′N, 1°19′W) lie on a hill rising from the Oxford Clay underlying the Thames floodplain (60 m above sea level) to the north, up through sandstone to coral rag limestone at 164 m above sea level. A soil survey of Wytham Woods was undertaken by the Commonwealth Forestry Institute in 1974 (CFI, Dawkins and Field 1978). This survey sampled soils on a chess-board pattern using the 1-ha grid of the national Ordnance Survey, and showed that Wytham’s varied geology is associated with a range in soil calcium that spans several orders of magnitude (63–23,000 mg/100 g−1soil) (Dawkins and Field 1978; Farmer 1995). The Wytham great tit population, which breeds largely in nestboxes, has been studied since the 1940s (Perrins 1979; Gosler 1993). The present study was undertaken on the north slope of Wytham Hill, an area of just 1 km2 containing 291 tit nestboxes out of about 1,000 in the whole Wytham estate. We used the mean calcium values for each 1-ha square of the 1974 CFI soil survey. Nestboxes were associated with their nearest soil-calcium values by taking the maximum calcium value of the four or five (as available) 1-ha squares nearest to the nestbox square. Soil-calcium (SC) values were normalized by Log10 transformation, and are referred to as ‘soil-calcium’ (see Gosler et al. 2005 for further detail).
Active mass loss during incubation
In 2002, 22 clutches were visited daily (by JPH) during the laying period to determine the clutch laying sequence (LS): the day’s egg was lightly numbered with a felt-tip pen. The clutches were selected at random within three main soil-calcium levels corresponding with clay (low calcium: < 1,000 mg/100 g soil), sand (medium calcium: 1,000–5,000 mg/100 g soil) and limestone (high calcium: > 5,000 mg/100 g soil) soils. Each egg in these nests was scored (by AGG) for intensity (I: 0–5), distribution (D: 0–5) and spot-size (S: 0–3) using the methods described by Gosler et al. (2000, 2005). The principal components pc1 and pc2 were calculated from the correlation matrix of I, D and S, and taken to represent the ‘darkness’ and ‘spread’ of pigmentation respectively (Gosler et al. 2005). Each egg was weighed in the field when fresh, then again during the 1st, and again during the 2nd week of incubation to an accuracy of 0.002 g on a portable Tanita 1210–100 digital scale. The difference in mass between these two weighings was used to calculate mass loss per day (MLD), and scaled for egg-size by including fresh (newly layed, unincubated) mass in the analyses as necessary (see 'Results' section). MLD was uncorrelated with the day of incubation on which the eggs were weighed. Since MLD is considered critical to egg viability, we also recorded, for each egg, whether or not it hatched (scored 1 or 0 respectively), and analysed these data separately.
Statistical analysis used multivariate modelling to determine the factors that predicted rates of MLD. As clutch size (CS) and lay date (LD: 1 April = 1) are important to great tit reproductive ecology (Perrins 1996; Gosler 1993), these were initially incorporated in each model. Also, as eggs from the same clutch are not statistically independent, data were analysed using Generalized Linear Mixed Models using Restricted Maximum Likelihood (REML in Genstat Release 8) with ‘Clutch ID’ as a random effect. For fixed effects, df of the reported Wald Statistic W is 1 unless stated otherwise, and effect direction is indicated + or −. Minimum adequate models (MAM) are reported, resulting from step-down model simplification. All bivariate interaction terms were incorporated initially in models, but those listed here are the ones subsequently found to be significant. REML Full Models used were (abbreviations as in methods above): (1) Model 1, which tests the general effects of factors on MLD during incubation. Response: MLD; random effects: ClutchID; fixed effects: LS, LD, CS, PD, PS, SC, EV, LS × PD, EV × PD; and (2) Model 2, which tests the effect of egg viability on MLD. Response: MLD; random effects: ClutchID; fixed effects: LS, LD, CS, EV, PD, PD × EV. Most of the abbreviations of factors are given above: those which are not are (a) EV—egg viability (binary factor: hatched or not hatched), (b) PD—pigment darkness, and (c) PS—pigment spread.
Passive mass loss
In order to confirm the physical determinants of passive MLD in fresh unincubated eggs (passive water loss), 15 clutches were selected in 2003 from nests on a range of soil-calcium levels (five low, five medium, and five high soil calcium, as above). These nests were visited daily during egg-laying (by AGG) and the day’s egg lightly numbered and pigment scored (I, D, S) as in 2002. Once the clutch was complete, 45 unincubated eggs were collected, representing the first, middle (3rd–5th depending on clutch size) and last eggs (in which every egg was pigment-scored for I, D and S). This was termed egg rank (ER: 1–3). The eggs were weighed to an accuracy of 0.00001 g on a Sartorius R160 D electronic balance, and measured (length, maximum breadth to 0.1 mm), and the breadth/length ratio (B/L), volume and surface-area (ESA) were calculated following Hoyt (1979). The time of weighing was recorded (h: min) and the eggs were refrigerated at 4.0°C until 16 May, when they were reweighed and moved to storage at 22.5°C (range determined by thermocouple: 22.0–23.0°C) until 24 May, when they were reweighed and returned to the refrigerator until 10 June, when they were again reweighed (see Table 1 for details). Exact weighing times were recorded, and MLD calculated from the mass change between the two weighings. These eggs were subsequently emptied and their shells reduced to ash in a Carbolite furnace at 800°C for 17 h, so vaporizing any organic matter (e.g. membrane) adhering to the shells; the ash was expressed as g/mm2 ash/surface area (EAM). See also Gosler et al. (2005) for further information concerning these eggs. When analysed together with ESA, variation in EAM largely reflects variation in eggshell thickness. Evidence for this assertion is presented below. MLD at the two temperatures (ET) was investigated by fitting Generalized Linear Mixed Models using Restricted Maximum Likelihood (REML in Genstat Release 8) with ‘Clutch ID’ as a random effect as described above. The REML Full Model used was; (abbreviations as above): Model 3, which tests for effects of factors determining passive MLD. Response: MLD; random effects: ClutchID; fixed effects: ER, LD, CS, PD, PS, SC, ESA, B/L, ET.
Cross-fostering experiment
In 2002 and 2003, we undertook experimental manipulations to determine whether egg mass loss during incubation was independent of female incubation behaviour (i.e. solely due to intrinsic eggshell properties), or whether a difference in incubation regimes between females affected the rate of mass loss (i.e. an external control). In each year, ten nests were visited daily to determine the laying sequence, and to score all eggs for maculation (as above). These were again chosen to cover a range of soil calcium conditions, but also so as to provide ten pairs (overall) of nests for experimental cross-fostering, paired for clutch-size (± 1 egg) and first-egg date (± 2 days) and offering a range of pigmentation. About 5 days into incubation, half of the eggs in each nest were swapped with half the eggs from its paired nest (in each year, odd-numbered eggs were swapped from five nests, and even-numbered eggs from the other five nests). Eggs were weighed in the field to 0.002 g (as described above) prior to being swapped. The eggs were re-weighed the following week, after which all the swapped eggs were returned to their natal nests so as to be reared by their natural mothers. Eggs were returned at least 2 days before the earliest estimated hatching date. Eggs left within their natal nests throughout the experiment acted as controls for those swapped. Although all clutches in the experiment hatched successfully in 2002, in 2003 significant egg mortality in one clutch, and desertion of one other reduced the sample to three pairs (six nests), making 16 nests in all over the 2 years.
The MLD of each egg was calculated from the two weighings, and within clutches the mean MLD of swapped eggs compared with the mean for the control eggs. Across clutches, the difference in mean mass of these two groups (swapped and control) was compared by linear regression with the difference in mean eggshell pigmentation between the donor and host clutches. Any significant correlation between the differences in mass loss between swapped and control eggs in a clutch, and the differences in pigmentation between host and donor clutches, must be due to some correlated difference in the incubation regime (temperature, behaviour etc.) of the two females.
Results
Passive mass-loss from unincubated eggs
The mass of great tit eggs collected prior to incubation was determined chiefly by their size (as Van Noordwijk et al. 1981), although some mass loss had already occurred from the earlier-laid eggs by the time that the last eggs were laid and the clutches collected (See Fig. 1). Based on a prediction made from the mass/surface area relationship for last-laid eggs, this amounted to 4.79±1.925% for first eggs, and 3.42±2.336% for middle eggs.

Egg mass and surface area correlate positively and linearly. Some mass loss appears to have occurred prior to collection, with greater mass loss in earlier eggs. In an ANCOVA, mass at collection is strongly dependent upon both surface area (F 1,86 = 980.7, P < 0.001) and egg rank (F 2,86 = 20.5, P < 0.001)
In storage, these eggs subsequently lost mass about 3.75 times more rapidly at 22.5°C than at 4.0°C; this difference was highly significant (t paired = 10.21, P < 0.001, n = 45). However, the rate of mass loss from the eggs when kept initially at 4.0°C did not differ from their rate of mass loss when returned to this temperature after having been kept at 22.5°C (t paired = 0.84, P = 0.406, n = 45). This indicates that for these unincubated eggs, the rate of mass loss at a given temperature was unaffected by the amount of water already lost, which, by the time the eggs were returned to 4.0°C, amounted to some 12.5%, on average, of predicted fresh mass (Table 1).
The Generalized Linear Mixed Model conducted on data from these unincubated eggs (Model 3) found three significant physical determinants of passive MLD. These were the egg’s surface area, (positive effect on MLD, W 1 = + 13.82, P < 0.001), ambient temperature, (strong positive effect on MLD, W 1 = + 205.81, P < 0.001), and eggshell ash mass, which is a surrogate for eggshell thickness when included in the model with surface area, (negative effect on MLD, W 1 = − 6.07, P = 0.014). The effects of egg surface area and ambient temperature on MLD of these eggs are shown in Fig. 2. No other predictor was a significant determinant of MLD.

The rate of passive mass loss (MLD) and egg surface area correlate positively and linearly with a large (3.75 times) increase in mass loss caused by an increase in temperature from 4.0–22.5°C. Eggs kept at 4.0°C maintained the same mass loss at 4.0°C, even when returned to this temperature after having been kept at 22.5°C. As passive results, these represent the intrinsic structural and physiological relationships between these parameters
Active mass loss during incubation
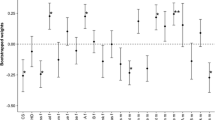
Unlike during passive water loss, when eggshell pigmentation and laying sequence had no affect on MLD, they exerted strong direct, or interactive, effects on active MLD. Of the seven main fixed effects included in the GLMM (Model 1) to predict active MLD, only the egg number (LS: W 1 = − 9.15, P = 0.002) and its squared term (LS2: W 1 = + 3.76, P = 0.052) were significant (marginally so in the latter case). However, a strong interaction was found between pigment darkness and egg viability (PD × EV: −22.91, P < 0.001) and a weaker one was found between pigment darkness and egg number (PD × LS: −4.45, P = 0.035). These relationships are depicted in Fig. 3a, which shows that the relationship between pigment ‘darkness’ and MLD changes significantly and markedly through the clutch. Earlier eggs in the laying sequence showed a positive relationship between pigment darkness and MLD, but this relationship became increasingly negative through the clutch. The between-clutch tendency for darker ‘early’ (within the clutch) eggs to lose more water was significantly greater in eggs that subsequently died (REML Model 2, Fig. 3a), suggesting that eggshell pigmentation might be critical for egg viability through its interaction with water loss (Fig. 3a). We found no evidence for an effect of pigment ‘spread’ on MLD. A number of eggs, which failed to hatch, lost substantially more mass than the average, suggesting that these might have had defective shells (Fig. 4).

The relationship between pigmentation and mass loss per day (MLD) during incubation differs with egg number across clutches. a the relationship between DML and eggshell pigment ‘darkness’ switches from positive to negative through the clutch. This was significant for viable eggs (filled symbols and solid line: Y = 0.0015511 [±0.0007831 s.e.] − 0.0000530 [± .0002573]X − 0.0000178 [± .00001788]X 2 r 2 = 72.9% F 2,10 = 13.4788 P = 0.001), but yet more striking when failed eggs were included (open symbols and dashed line: Y = 0.0045771 [±0.0006858] − 0.0005211 [±0.00008640]X, r 2 = 76.8% [polynomial: 76.9%], F 1,11 = 36.38, P < 0.001). Across the first eight eggs, the interaction of slope between ‘darkness’ and egg-viability was significant (REML Model 2, W = 5.69, P = 0.017), indicating that egg mortality was associated with excessive water-loss for the degree of pigmentation. Clutch size (W = 6.87, P = 0.009) and egg viability (W = −43.06, P < 0.001) were also strong predictors of MLD. Note that here each point represents each egg number across all clutches. b The within-clutch relationship between ‘darkness’ and MLD varies from negative to positive with increasing soil calcium (Y = −0.0116651 [±0.003549] + 0.0029732 [±0.0009089]X, r 2=36.0%, F 1,19 = 10.70 P = 0.004). Note that here each point represents a clutch. The X-axis is a Log10 scale, such that ‘3’ represents a soil calcium value of 1,000 mg/100 g, and ‘4.5’ approximately 31,600 mg/100 g

The rate of active mass loss (MLD) is uncorrelated with egg fresh mass (which is almost synonymous with surface area—see text and Fig. 1), showing that the relationships demonstrated in Fig. 2 are no longer valid under incubation. This result shows that the nest environment provided for the female specifically compensates for intrinsic differences between the eggs. The combined trend Y = 0.2043 [±0.2370 s.e.]X − 4.2152 [± 0.4047] is non-significant, r 2 = 0.0061
Although not indicated by the general model reported above, we also found evidence that the within-clutch slope of the relationship between MLD and ‘darkness’ interacted strongly with soil calcium (Fig. 3b). Thus in clutches on low-calcium soils, darker (later) eggs lost water more slowly than their paler sibs, whilst in clutches on high-calcium soils darker (later) eggs lost water more rapidly than their paler sibs. Whilst this result is difficult to comprehend, we suggest an explanation in the discussion below.
These complex relationships between rates of mass loss during incubation and egg characteristics such as pigmentation, which differ strikingly from the simple physical effects observed in eggs allowed to lose water passively, indicate a strong interaction between the clutch and the microenvironment produced by the female. Further evidence of this effect is presented in the results of the cross-fostering experiment.
Cross-fostering experiment: incubation effect on MLD
Across nests, we found a significant correlation between the difference in rate of mass loss between experimental and control eggs within a nest pair, and the difference in mean pigment darkness (pc1 diff.) between the two clutches (r 14 =0.609, P=0.011, Fig. 5). This relationship did not differ significantly between years (year × pc1 diff. F 1,12 =0.00, P=0.99, pc1 diff. F 1,12 =7.10, P=0.021), and was not influenced by the experimental pair (exp. pair F 7,7 = 1.20, P=0.406, pc1 diff. F 1,7 =9.6, P=0.017).

Results of a cross-fostering experiment to determine whether variation in female-specific incubation regime (temperature, time on: time off etc.) affects the rate of water loss (represented by mass loss per day—MLD) in relation to clutch pigmentation. Egg number in the laying sequence, pigmentation scores and egg mass (to 0.002 g) were determined for each egg in 16 clutches over 2 years. Half clutches (odd or even egg numbers) were then swapped between paired clutches for at least 4 days during incubation. They were then reweighed and returned to their natal nests to hatch. For each clutch, the rate of MLD in swapped eggs was compared with that of control eggs from the same clutch. The difference in MLD for a given clutch is therefore independent of that in any other nest (neither year nor experimental pair influenced the relationship significantly—see text). The figure shows the relationship between this difference and the difference in pigment darkness (pc1) between the two nests in each pair. The significant regression (Y = 0.000104 [±0.0008469 s.e.] + 0.00184 [±0.0006522]X, r 2 = 31.7%, F 1,14 = 7.96, P=0.014) indicates that variations in the incubation environment (e.g. female incubation regime) influence the relationship between MLD and pigmentation. The difference in MLD was not correlated significantly with the difference in local soil calcium level (r 14 = 0.338, P=0.200)
Thus in both years, eggs transferred to females with a more pigmented clutch than the donor’s lost more water than control eggs left with the donor (Fig. 5). Conversely, eggs transferred to a female with a less-pigmented clutch than the donor lost less water than eggs remaining with the donor. The intercept of the regression of mass-loss difference on pigment difference did not differ significantly from zero (intercept: 0.0001654±0.0008258 s.e., T =0.20, P=0.844), implying that eggs transferred between identically-pigmented clutches should show near-identical mass-loss.
This experiment demonstrates for the first time that female incubation varies with maculation, and that females with more pigmented clutches appear to have increased incubation effort compared with females laying less-pigmented clutches. As in the overall relationship between pigmentation and MLD, the difference in pigment spread (pc2) between nests was not a significant predictor of the difference in mass loss between swapped and control eggs (r 14 = −0.458, P=0.074). It appears, therefore, that rates of water loss are affected specifically by the darkness (which indicates the depth of ‘pitting’ in the shell: Fig. 3 in Gosler et al. (2005), and perhaps also the quantity of pigment present) rather than the spread of pigment.
Discussion
For present purposes, the bird’s egg can be considered as a water-filled bag encased within a semi-permeable calcium carbonate shell. Therefore the rate at which water is conducted through the shell, and that mass is lost from the egg, should be determined by the egg’s size (specifically surface area), shell thickness and the ambient temperature (and humidity). We have shown this to be so here for great tit eggs by studying passive MLD, whilst also demonstrating that the eggshell’s pigmentation does not affect passive MLD. However, we then showed that, in eggs incubated by females (active MLD), maculation and female incubation behaviour combined to affect rates of water loss. Once incubation and embryonic development within the egg commence, the relationship between eggshell thickness and water loss becomes more dynamic than that seen in ‘passive’ eggs because, for example, the shell thickness declines as calcium is withdrawn from it by the embryo, and the female interacts behaviourally with the clutch. In this paper, we have demonstrated unexpected relationships between eggshell thickness, maculation, and female incubation behaviour and active MLD.
The ‘structural–function hypothesis’ (Gosler et al. 2005) for the function of eggshell maculation predicts relationships between pigmentation, calcium availability and mass-loss not expected from any signalling function. Here, we have provided further support for the hypothesis by showing links between maculation and egg mass loss during incubation, and by demonstrating a relationship between female incubation behaviour and clutch maculation. Thus, we have provided detailed evidence of structural and physiological effects of eggshell maculae. As we have argued before (Gosler et al. 2005), since egg speckles convey information about shell structure, this does give them potential as intraspecific signals of shell quality, but signalling functions for this type of eggshell maculae are likely to be epiphenomena.
The intra-clutch relationships shown between mass loss and pigment demonstrate that maculation is a significant factor affecting the intrinsic property of eggshell permeability in great tit eggs. Furthermore, the results of the cross-fostering experiment indicate that females can exert an external control over this, suggesting that they may have the behavioural flexibility to match the incubation environment appropriately to the structural properties of the clutch that they have laid. The incubating female, together with her nest and clutch of eggs, would, therefore, constitute an integrated unit, in which clutch characteristics are not independent of the female’s incubation regime.
The mechanism by which protoporphyrins could affect water loss through the shell is unclear. The simplified eggshell structure of small passerines (Mikhailov 1997) described above, in which calcite crystals are separated by interstices forming a lattice structure, means that, in thinner shell, water loss may be enhanced by the increased connectivity between vesicles and between shell pores. If protoporphyrin pigments were to fill these interstices where applied, they might physically reduce this effect in thinner shells. Although, as noted earlier, it may be important that protoporphyrins reflect heat, detailed analysis under laboratory conditions would be required to test whether this might have any effect at the very fine scale required to make this relevant to small eggs such as those of the great tit.
The changing intra-clutch relationships between mass loss and maculation in relation to soil calcium are not straightforward, and can be difficult to understand. Why, for example, should increased maculation apparently increase eggshell permeability on high-calcium soils but decrease it on low-calcium soils? To understand this, it must be remembered that the presence of pigment indicates localized eggshell thinning, and that darker pigment implies ‘deeper’ thinning (Gosler et al. 2005). Our interpretation, then, is that protoporphyrins are incorporated in the shell principally to strengthen it, and that although pigment can counteract the effect of eggshell thinning on permeability, it is not a perfect adaptive solution to this problem. Thus, on high-calcium soils, where eggshells are generally thicker and less pigmented overall, a small amount of extra pigment is added through the clutch to maintain eggshell strength in later eggs (with slightly darker and more pitted shells), but this is inadequate to prevent an increase in water loss caused by the eggshell thinning. On low-calcium soils, where shells are generally thinner and more heavily pigmented, the greater amounts of pigment used to maintain shell strength cause an overall water-loss reduction, which is even greater in the later eggs that are heavily pigmented. Thus, while pigment could affect both shell strength and permeability, the relationship between pigment and each of the two variables may not be parallel, and is not necessarily linear in either. Indeed, we suggest that it would be very remarkable if protoporphyrin offered a perfect solution (in terms of relative effect per unit pigment) to both of the problems arising from reduced eggshell thickness, namely reduced strength and increased permeability. Thus, we argue that the presence of relationships similar to those depicted in Fig. 3a and b are likely consequences (i.e. they should be expected) of the system described here and in Gosler et al. (2005).
This explanation is consistent with, and indeed predicts, the results obtained from the cross-fostering experiments (Fig. 5), in which females that laid darker clutches showed more intense incubation effort than did females that laid paler clutches, since they would need to increase the conductivity of these darker eggshells. Although females can move eggs within the nest, and may thereby change the amount of heat received by specific eggs, we assume that a female’s incubation behaviour will in general provide a response to the average conductance properties of the clutch, and may therefore represent a compromise between the ideal requirements of different eggs. Thus, while we view the incubating female and her clutch as an integrated unit, nevertheless the extrinsic component (the incubation regime of the female and insulating properties of her nest) must have the flexibility to adapt to variation within the intrinsic components (the effects of intra-clutch variation in egg-size, eggshell thickness, permeability, pigmentation and hence conductivity). This compromise may have the effect that eggs towards the extremes of the distribution (perhaps earlier or later eggs in a clutch) might receive too much or too little heat, and so we might expect intra-clutch egg mortality to be non-random with respect to egg number. This is supported by the increased relationship between MLD and pigment darkness in eggs that subsequently died, and the fact that this was especially seen in earlier eggs in the laying sequence (Fig. 3a). Our study might also shed light on the reasons for hatch failure sometimes encountered in cross-fostering experiments conducted, for example, in studies of population genetics.
A variety of mechanisms could stimulate the behavioural adjustment of incubation in accordance with clutch conductance. Physiological cues related to calcium itself might predispose females towards certain incubation regimes. Alternatively, females could use proximate cues in the nest to optimize incubation behaviour. We consider the latter case to be much more likely, and that females use cues such as nest humidity to assess current rates of water loss, and so to adjust their behaviour appropriately. These two types of hypothesis (physiological versus proximate indicators) might be distinguished by testing the hypothesis in species in which both males and females incubate. If males adjusted their incubation behaviour according to clutch maculation, this would be a strong indicator that proximate nest cues were used. The birds’ use of nest humidity as a stimulus could also be tested experimentally by using temperature data loggers to record female incubation behaviour, whilst manipulating nest humidity with water-absorbing agents such as silica gel.
Further investigation of the structural function hypothesis is likely to require several different approaches. Experimental calcium enrichment is known to affect eggshell thickness and egg volume (Mand et al. 2000; Tilgar et al. 1999), and is likely to provide a key test of hypotheses presented here and in Gosler et al. (2005). Investigation of potential mechanisms by which pigment may inhibit water loss could include high-resolution SEM analysis of the shell structure (particularly shell pores) in relation to pigment. Finally, two provisos should be noted. First, some of this study’s results are correlative, and it remains possible that a third factor, one that co-varies with maculation, could be involved in determining the water-loss relationships observed here. Secondly, no conclusions on the generality or specificity of this work can be derived until comparative studies have assessed intra-clutch patterns of maculation, and correlates with eggshell properties, across a range of taxa in a range of environments.
References
Ar A, Rahn H (1978) Interdependence of gas conductance, incubation length and weight of the avian egg. In: Piiper J (ed) Respiratory function in birds, adult and embryonic. Springer, Berlin Heidelberg New York, pp 227–236
Ar A, Paganelli CV, Reeves RB, Greene DG, Rahn H (1974) The avian egg: water vapour conductance, shell thickness, and functional pore area. Condor 76:153–158
Ar A, Rahn H, Paganelli CV (1979) The avian egg: mass and strength. Condor 81:321–327
Bakken GS, Vanderbilt VC, Buttemer WA, Dawson WR (1978) Avian eggs: thermoregulatory value of very high near-infrared reflectance. Science 200:321–323
Blanco G, Bertellotti M (2002) Differential predation by mammals and birds: implications for egg-colour polymorphism in a nomadic breeding seabird. Biol J Linn Soc 75:137–146
Davies NB, Brooke MD (1989a) An experimental-study of co-evolution between the Cuckoo, Cuculus-Canorus, and its hosts. 1. Host egg discrimination. J Anim Ecol 58:207–224
Davies NB, Brooke MD (1989b) An experimental-study of co-evolution between the Cuckoo, Cuculus-Canorus, and its hosts. 2. Host egg markings, chick discrimination and general discussion. J Anim Ecol 58:225–236
Dawkins HC, Field DRB (1978) C.F.I. Occasional Paper No. 1. A Long-Term Surveillance System for British Woodland Vegetation. Commonwealth Forestry Institute, University of Oxford, Oxford
Farmer AM (1995) Soil chemistry change in a lowland English deciduous woodland. Water Air Soil pollut 85:677–682
Gosler AG (1993). The Great Tit. Hamlyn species guides. Paul Hamlyn, London
Gosler AG, Barnett PR, Reynolds SJ (2000) Inheritance and variation in eggshell patterning in the great tit Parus major. Proc R Soc Lond B Biol Sci 267:2469–2473
Gosler AG, Higham JP, Reynolds SJ (2005) Why are birds’ eggs speckled? Ecol Lett 8:1105–1113
Graveland J, Berends AE (1997) Timing of the calcium intake and effect of calcium deficiency on behaviour and egg laying in captive great tits, Parus major. Physiol Zool 70:74–84
Graveland J, Drent RH (1997) Calcium availability limits breeding success of passerines on poor soils. J Anim Ecol 66:279–288
Graveland J, Vanderwal R, Van Balen JH, Van Noordwijk AJ (1994) Poor reproduction in forest passerines from decline of snail abundance on acidified soils. Nature 368:446–448
Hoyt DF (1979) Practical methods of estimating volume and fresh weight of birds eggs. Auk 96:73–77
Kennedy GY, Vevers HG (1976) A survey of avian eggshell pigments. Compu Biochem Physiol 55B:117–123
Lack D (1968) Ecological adaptations for breeding in birds. Methuen, London
Mand R, Tilgar V, Leivits A (2000) Reproductive response of Great Tits, Parus major, in a naturally base-poor forest habitat to calcium supplementation. Can J Zool-Rev Can Zool 78:689–695
Mikhailov KE (1997) Avian eggshells: an atlas of scanning electron micrographs., BOC occasional publications: 3 British Ornithologists’ Club
Moreno J, Osorno JL (2003) Avian egg colour and sexual selection: does eggshell pigmentation reflect female condition and genetic quality? Ecol Lett 6:803–806
Moreno J, Osorno JL, Morales J, Merino S, Tomás G (2004) Egg colouration and male parental effort in the pied flycatcher Ficedula hypoleuca. J Avian Biol 35:300–304
Newton AV (1896) A dictionary of birds. A. C. Black, London
Perrins CM (1979) British Tits. Collins, London
Perrins CM (1996) Eggs, egg formation and the timing of breeding. Ibis 138:2–15
Rahn H, Ar A (1974) The avian egg: incubation time and water loss. Condor 76:147–152
Sanchez JM, Corbacho C, del Viejo AM, Parejo D (2004) Colony-site tenacity and egg color crypsis in the Gull-billed Tern. Waterbirds 27:21–30
Seebohm H 1896 Eggs of British birds. Pawson & Brailsford, Sheffield
Sibley CG, Monroe BL Jr (1990) Distribution and taxonomy of the birds of the world. Yale University Press, New Haven
Soler JJ, Moreno J, Aviles JM, Møller AP (2005) Blue and green egg-color intensity is associated with parental effort and mating system in passerines: support for the sexual selection hypothesis. Evolution 59:636–644
Solomon SE (1987) Egg shell pigmentation. In: Wells RG, Belyarin CG (eds) Egg quality—current problems and recent advances. Butterworths, London, pp 147–157
Solomon SE (1997) Egg and eggshell quality. Iowa State University Press, Ames
Tazawa H, Whittow GC (2000) Incubation physiology. In: Sturkie PD, Whittow GC (eds) Avian physiology. Academic Press, London
Tilgar V, Mand R, Leivits A (1999) Effect of calcium availability and habitat quality on reproduction in Pied Flycatcher Ficedula hypoleuca and Great Tit Paras major. J Avian Biol 30:383–391
Tyler C (1969) Avian egg shells: their structure and chararcteristics. In: Felts WJL, Harrison RJ (eds) International review of general and experimental zoology. Academic Press, New York
Van Noordwijk AJ, Keizer LCP, Van Balen JH, Scharloo W (1981) Genetic variation in egg dimensions in natural populations of the Great Tit. Genetica 55:221–232
Weimar V, Schmidt K-H (1998) Untersuchungen zur Eiqualitat bei der Kohlmeise (Parus major) in Abhangigkeit von der Bodenbeschaffenheit. Journal fur Ornithologie 139:3–9
Acknowledgments
This work was carried out under English Nature licences to AGG—2002: 20020828 and 20012058, and 2003: 20030589, 20030184 and 20031149. Robin McCleery made the CFI calcium data available to us. We are grateful to Ben Sheldon, Dany Garant, Tom Pizzari and three anonymous referees for their constructive criticism of earlier drafts, and Stuart Semple, Markku Orell and one anonymous referee for helpful comments on the present one. JPH was supported by a BBSRC studentship, AGG by the Edward Grey Institute, University of Oxford.
Author information
Authors and Affiliations
Corresponding author
Additional information
Communicated by Markku Orell.
Rights and permissions
About this article
Cite this article
Higham, J.P., Gosler, A.G. Speckled eggs: water-loss and incubation behaviour in the great tit Parus major . Oecologia 149, 561–570 (2006). https://doi.org/10.1007/s00442-006-0484-2
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00442-006-0484-2