
Abstract
Knowledge about plant–plant interactions for pollinator service at the plant community level is still scarce, although such interactions may be important to seed production and hence the population dynamics of individual plant species and the species compositions of communities. An important step towards a better understanding of pollination interactions at the community level is to assess if the variation in floral traits among plant species explain the variation in flower visitation frequency among those species. We investigated the relative importance of various floral traits for the visitation frequency of all insects, and bumblebees and flies separately, to plant species by measuring the visitation frequency to all insect-pollinated species in a community during an entire flowering season. Visitation frequency was identified to be strongly positive related to the visual display area and the date of peak flowering of plant species. Categorical variables, such as flower form and symmetry, were important to the visitation frequency of flies only. We constructed floral similarity measures based on the species’ floral traits and found that the floral similarity for all species’ traits combined and the continuous traits separately were positively related to individual visitation frequency. On the other hand, plant species with similar categorical floral traits did not have similar visitation frequencies. In conclusion, our results show that continuous traits, such as flower size and/or density, are more important for the variation in visitation frequency among plant species than thought earlier. Furthermore, differences in visitation frequency among pollinator groups give a poor support to the expectations derived from the classical pollination syndromes.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
The interactions among plant species and pollinators at the plant community level are complex and still poorly understood, although an increasing number of pollination studies are concentrating on interactions at the community level (e.g. Olesen and Jordano 2002). The community approach is necessary to draw general conclusions about pollination interactions (Jordano 1987; Memmott 1999; Waser et al. 1996) and studies of plant–plant interactions, for pollination are an important part of the community approach. The two most common plant–plant interactions are pollinator attraction and heterospecific pollination (e.g. Campbell and Motten 1985; Waser and Fugate 1986; Feinsinger 1987; Caruso 1999). While heterospecific pollination is a purely competitive interaction, pollinator attraction can also be facilitative. As a consequence, attractive and generalist plant species that have many interactions with animals in visitation webs have the largest potential to affect other plant species through both types of plant–plant interactions.
Other studies describing plant–pollinator interactions at the community level (e.g. Herrera 1988; Petanidou and Vokou 1993; Bosch et al. 1997; Memmott 1999; Dicks et al. 2002; Dupont el al 2003) have not focused on why plant species differ in their attractiveness to potential pollinators, although there is a large variation in floral traits among species within a community. Simultaneously, there are large variations in the attractiveness, in terms of flower visitation frequency, among plant species of the same community; some are highly attractive while others seem never to receive visits (e.g. Schemske et al. 1978; Pleasants 1980; Herrera 1988; Inouye and Pyke 1988; McCall and Primack 1992; Totland 1993; Elberling and Olesen 1999; Memmott 1999). Surprisingly, very few have studied if this large variation in interspecific pollinator attractiveness is related to the variation in floral traits among species (but see McCall and Primack 1992). Several relevant ecological questions can be addressed by investigating this issue. First, can commonly observed patterns within species, such as increased pollinator attraction to larger flowers (Conner and Rush 1996; Goulson et al. 1998; Ohashi and Yahara 1998; Pickering 2001) also be established in among-species relationships? Second, do species with a similar combination of floral traits have similar attractiveness on pollinators, and can such information help us predict how species interact (i.e. facilitate or compete) for pollinator visitation, for example if any species within a plant community act as ‘magnets’ (e.g. Laverty 1992; Memmott 1999)? Third, can classical pollination syndromes (e.g. Fægri and van der Pijl 1979), that mainly focus on categorical floral traits, explain why pollinator groups (e.g. bumblebees and flies) tend towards preferring some plant species above others, or are continuous floral traits, such as flower size and floral density, more important in explaining the variation in visitation frequencies of such pollinator groups? We address these questions by examining how species’ floral traits (number of visual displays, visual display area, number of flowers per visual display, length of flowering period, date of peak flowering, flower colour, flower shape, flower symmetry, and abundance of insect-pollinated plant species within a community) relate to visitation frequency of plant species within a community.
Materials and methods
Study community
Our study site is a species-rich grassland/meadow of the intermediate dry, medium base-rich lowland grassland-type (Fremstad 1997), situated on the border between the boreo-nemoral and south-boreal zone in the inner region of the Sognefjord, west Norway. The grassland is located in a steep slope about 10–20 m above sea level and 50 m away from the shore. The climate is relatively warm and dry; the closest climate station (Lærdal) had a mean July temperature of 17.3°C and temperatures ranging from 7.7°C to 30.5°C during the study season (DNMI 2003).
We delimited an area of approximately 50×50 m by an electric fence during the field season to avoid disturbance by sheep and red deer that have grazed in the area since the fields were abandoned in the early 1960s. The surrounding area is dominated by deciduous forest and contains several patches of similar floral composition as the study site. A total of 38 insect-pollinated flowering species, with different floral traits, occurred inside the study site. Twenty-seven of these species flowered simultaneously at peak flowering in mid-July (see Table 1 for the plant species included in the analysis). The nomenclature follows Lid and Lid (1994).
Bumblebees (Bombus ssp.) and flies (Diptera) dominated the visitor pool and accounted for 61% and 26 % of the total 3,634 recorded visits, respectively. The other pollinator groups recorded were different bees (Apidae, except bumblebees, 5%), beetles (Coleoptera, 5%), butterflies (Lepidoptera, 2%) and ants (Formicidae, 1%).
Data collection
We sampled data between 28 May 2003 and 18 August 2003, with the highest sampling intensity in July. This period covered the entire flowering season of most plant species in the community, and thus the main period for pollination. To determine the attractiveness of each plant species in the community to potential pollinators, we conducted a visitation survey inside 20 permanent plots of 1.5×1.5 m, randomly positioned at the start of the fieldwork and used throughout the entire season. Within the plots, we monitored the visitation frequency by counting the number of visits, including identification of the main visitor group (Diptera, Bombus spp., non-Bombus Apidae, Lepidoptera, Coleoptera, Formicidae), to visual displays for every insect-pollinated plant species during 10-min observation periods. Most periods were obtained between 10 a.m. and 4 p.m. In total we conducted 201 (i.e. 2,010 min) observation periods during the flowering season. We reduced the amount of non-pollinating insects in our data set by only counting the visits, where the visitor actually landed on the visual display and stayed for more than 1 s. To measure floral density, we counted the number of visual displays of each plant species inside the plot after every period. Depending on the species, a visual display consisted of a single flower, a flowering head/inflorescence or a group of flowers occurring together in a recognisable visual unit (see Table 1). We measured the area of visual displays, as a measure of flower size, of 10–20 randomly selected plants of each species and calculated the mean visual display area of each species according to the shape of the flowers. The area of visual displays with a circular outline (e.g. Leucanthemum vulgare) was calculated using the formula: πr2, and for other flat visual displays (e.g. Achillea millefolium) we used the formula: length×width. When visual displays also had a depth dimension (e.g. Trifolium pratense), the formula: 2πr2d+πr2 were used (see Table 1 for formulas used for each species). On the same random collection of plants, we also counted the number of flowers per visual display and calculated the mean per species. We determined the length of the flowering period and the date of peak flowering for each species by counting the number of open visual displays inside all the 20 plots, every 3–6 days during the season. For those species that we could not determine the actual start or end of flowering because it extended the study period (e.g. Potentilla erecta), we extrapolated their flowering period curves by a maximum of 15 days in any direction to obtain a date of flowering start and/or stop. Unfortunately, we were unable to obtain comparable data on nectar and pollen amounts of each species as a consequence of the large amount of species and their temporal variation in floral reward production (see also Lack 1982).
Data analysis
To examine the relationship between single floral trait variables and visitation frequency of species, we performed simple linear regressions for continuous variables and one-way analysis of variance (ANOVA) for categorical variables (Table 2). We used stepwise analysis of co-variance (ANCOVA) to determine the relative importance of floral traits (both continuous and categorical) in explaining variation in pollinator visitation frequency among species on the community level. In the stepwise ANCOVA, we used a backward selection of variables because some of the categorical variables had more than two levels (Zar 1999). We checked all the data and the residuals from the ANCOVA with normal probability plots and tested the standardised residuals with a Kolmogorov-Smirnov test for normality, using the Lilliefors option in SYSTAT (Wilkinson 2003). Variables with a non-normal distribution were log-transformed, until normality of residuals was ensured. All models presented passed these assumption tests. We also inspected the residuals to detect outliers (leverage >0.2), but since none of the potential outliers affected the direction or statistical significance of the results in our analyses we chose not to remove them.
We tested the homogeneity of slopes-assumption of ANCOVA according to the methods described by Glantz and Slinker (2001). No interactions between categorical and continuous variables occurred, showing that none of the continuous variables had heterogeneous slopes with the response variable.
The visitation frequency of each species (response variable) was defined as the mean number of visits per period, and thus a measure of the species’ attractiveness to potential pollinators. Our predictors included the continuous variables: (1) number of visual displays (mean per species calculated from the mean per plot), (2) number of flowers per visual display (mean per species), (3) visual display area (mean per species), (4) length of flowering period (number of days per species), (5) date of peak flowering (per species), and the categorical variables: (6) flower shape (open or tubular flowers), (7) flower symmetry (actinomorphic or zygomorphic flowers), (8) flower colour (white, yellow, blue-violet or pink-red flowers; as seen by human eyes), (9) abundance in the community (rare: flowering in 1–6 plots; intermediate: flowering in 7–13 plots; and abundant: flowering in 14–20 of the 20 plots at the peak flowering of each species). Hereafter we commonly term these predictors as floral traits.
Those plant species that were observed in less than 11 periods throughout the study period were not included in the final analysis, yielding a sample size of 27 plant species that were visited by all the insects (all insects). Analyses using visitation frequency of bumblebees (Bombus ssp.; 18 plant species) and flies (Diptera; 25 plant species) were performed separately. Other groups of pollinating insects (e.g. beetles and butterflies) did not perform enough visits to a sufficient amount of plant species to be included as separate visitor groups in the statistical analysis. However, their visits are included in the visitation frequency of all the insects group.
We used the principal component analysis (PCA) to ordinate the plant species according to their floral similarity. Species with similar scores along the first axis have a similar set of traits (i.e. higher floral similarity), whereas species that are far from each other along the first axis have dissimilar floral traits. The PCAs were run using all floral traits combined and the continuous and categorical predictors separately. We assessed if the degree of floral similarity among species was related to the visitation frequency by performing a simple linear regression between the scores of the first axis and the visitation frequency of all the insects. All statistical analysis was done with the statistical program SYSTAT 10.0 (Wilkinson 2003).
Results
Single predictor models
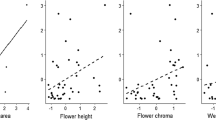
Visitation frequency of all insects combined increased with visual display area (N=27, P=0.005; Fig. 1a) and date of peak flowering (N=27, P= 0.001; Fig. 1b), but was not significantly affected by the number of visual displays per plot; number of flowers per visual display and length of flowering period (N=27, all P>0.35). None of the categorical variable explained differences in all insects visitation frequencies (P>0.57; Table 1).

a–d Simple linear regressions on the relationship between mean visitation frequency of all insects and a visual display area and b date of peak flowering; and between mean visitation frequency of bumblebees and c visual display area and d date of peak flowering among insect-pollinated plant species in the temperate grassland Rudsviki by the Sognefjord in west Norway. Only the significant relationships are presented. See text for P values of non-significant relationships. Each point represents a plant species and is based on mean values from all observations on that plant species. The multiple coefficients of determination (R2), probability values (P) and numbers of observations (N) are given for each relationship. The full species names are given in Table 1
Bumblebee visitation frequency also increased with visual display area (N=18, P=0.016; Fig. 1c) and date of peak flowering (N=18, P=0.004; Fig. 1d), but was not significantly affected by the number of visual displays per plot; number of flowers per visual displays and length of flowering period (N=18, P>0.29). None of the categorical variable explained differences in bumblebee visitation frequencies (P>0.37; Table 1).
Visitation frequency of flies showed no significant linear relationships with any of the continuous predictors: visual display area, date of peak flowering, number of flowers per visual displays and length of flowering period (N=25, P>0.15 for all variables), but were marginally positive related to the number of visual displays per plot (N=25, P=0.09). On the other hand, flies visited plant species with open flowers more frequently than species with tubular flowers (F 1,23=6.82, P=0.016,) and species with actinomorphic flowers more frequently than species with zygomorphic flowers (F 1,23=4.72, P=0.04). None of the other categorical variables explained differences in the visitation frequency of flies (flower colour: F 3, 21=0.75, P=0.53; abundance: F 2, 22=0.06, P=0.95) among plant species for any of the visitor groups when analysed separately.
Multiple predictor models
All stepwise ANCOVA models explained a substantial amount of the interspecific variation in visitation frequency for each visitor group (R 2≥0.45; Table 3). Both visitation frequency of all insects and bumblebees increased with visual display area and date of peak flowering, while there were no differences in visitation frequency for categorical variables (Table 3). The final model for flies included six significant or close to significant predictors (Table 3). Visitation frequency of flies increased with the length of flowering period, visual display area and date of peak flowering, while it decreased with the number of flowers per visual display. Visitation frequency of flies also differed significantly between flower symmetries and among flower colours.
The PCA on all floral traits of species resulted in a first axis, explaining 27.4% of the total variation in floral traits among species. Species scores along the first PCA-axis showed a significant relationship (N=27, P=0.006) with the visitation frequency of all insects, indicating that plant species with similar combinations of floral traits had more equal visitation frequency than species with dissimilar multivariate expression of floral traits (Fig. 2). A PCA with only continuous floral traits yielded a first axis explaining 35.6% of the total variation in floral traits. There was a significant relationship between species scores along the first axis and visitation frequency of species (P=0.003, R 2=0.30). A PCA on the categorical predictors explained 52% of the total variation in the components, but the species scores along the first axis showed no significant relationship with the visitation frequency (P=0.14, R 2 =0.09).

Simple regression on the relationship between mean visitation frequency of all insects and species scores of floral traits along the first PCA-axis for each insect-pollinated plant species in the temperate grassland Rudsviki by the Sognefjord in west Norway. The PCA is based on all floral traits measured (see Materials and methods). The multiple coefficients of determination (R 2), probability values (P) and numbers of observations (N) are given. The full species names are given in Table 1
Discussion
To our knowledge, no other studies have examined the relative importance of floral traits of species for attraction of potential pollinators at the interspecific level within a community, even if such data often are available. In our study, the area of visual display appears to have a particularly important, and positive, effect on a species ability to attract potential pollinators. Since there is a positive correlation between the visual display area of a species and its floral density in patches (i.e. the product of a species’ mean visual display area and mean number of visual displays within the plots; r=0.801, P<0.001), we cannot determine the relative importance of display area and floral density in patches for the interspecific variation in visitation frequency. Both species with large displays, like Filipendula ulmaria and Centaurea jacea, or species that often occur with high floral density in patches, like T. pratense and Prunella vulgaris, are among the most attractive to visiting insects. Interestingly, our results at the interspecific level are in accordance with patterns found on the intraspecific level, demonstrating that individuals with many flowers or that occur in dense conspecific patches often receive most visits (e.g. Goulson et al. 1998; Ohashi and Yahara 1998; Totland and Matthews 1998; Pickering 2001). Potential pollinators may prefer plant species with larger displays and/or those growing in dense patches to minimise costs of searching for rewards. Visual cues in themselves, such as the area of the floral display, can also influence the decisions of which flowers or inflorescences within a plant population an individual insect chooses to visit (Medel et al. 2003). Moreover, within a species there is often a correlation between the area of the visual display and the amount of nectar (Stanton and Preston 1988; Cresswell and Galen 1991; Duffield et al. 1993; Inoue et al. 1995) or pollen (Stanton and Preston 1988) produced. If correlations between flower size and reward quantity are also established in interspecific relationships, that may explain why visual display area is important in explaining the variation in pollinator attraction among plant species. Bumblebees, that dominated the visitor pool in our study, may learn quickly and adapt to flower species with the highest reward (Goulson 2003 and references therein). Therefore a correlation between floral display area and floral rewards may increase the visitation to plant species with such visual cues and indicate that visual cues can be important for insects’ choice of flowering species in a species-rich plant community.
Pollinator species do not necessarily base their choice of flowering species on single floral traits. Indeed, our correlation between floral trait similarity among plant species and visitation frequency showed that plant species with similar combinations of traits have comparable attractiveness to potential pollinators. This indicates that several floral traits affect the flower choice of insects. Given that there was no significant relationship between PCA-axis 1 based on categorical variables and visitation frequency (only a trend; P=0.14), whereas a similar regression based on continuous floral traits was highly significant (P=0.003), our results also suggest that the role of continuous floral traits in explaining variation in among-species attraction of potential pollinators is larger than thought earlier (see for example Fægri and van der Pijl 1979).
Late-flowering species had higher visitation frequencies than early-flowering species, a result consistent with earlier studies in similar habitats (e.g. Thomson 1982; McCall and Primack 1992). An interpretation of this pattern has at least two components. First, the number of potential pollinator species and their abundance will normally increase throughout the season in northern lowland ecosystems, and hence many plant species that flower relatively late have the highest visitation. In our study area, especially bumblebees increase in abundance during the flowering season (Hegland and Totland, unpublished). Second, the number of insect-pollinated species in bloom increased during the flowering season at our study site (Hegland and Totland, unpublished). As a consequence, there is no competition for pollinator attraction among plant species, thus there is higher visitation frequency to species flowering simultaneously late in the season. Feinsinger (1987) argued that facilitation for pollinator visitation among plants by co-flowering could be at least as important as competition resulting in divergent flowering. However, only a handful of studies have reported indications for facilitative interactions to attract pollinators among co-flowering plant species (Thomson 1978; 1981; Campbell and Motten 1985; Laverty 1992).
The term “magnet”-species has been used for highly attractive plant species that facilitate visitation frequency of less attractive plant species (Laverty 1992). Based on the ANCOVA models for all insects’ and bumblebees’ visitation frequency, such a potential “magnet”-species in our study community has large visual displays, alternatively high floral density in patch, and late flowering, for example Filipendula ulmaria, C. jacea and T. pratense (see Fig. 2). Identifying potential “magnet”-species among the fly-pollinated species is more complicated since there are many more traits included in the ANCOVA-model, but P. erecta is one of the species that closely fit the predictions made by that model, such as long flowering period and actinomorphic flowers (see Table 3 for the other significant traits). In an analysis of a grassland community, Memmott (1999) found that on plant species, Daucus carota attracted many more visitors (42% of total) and visitor species (61% of total) than the other species in the community. As suggested by Memott (1999), an interesting experiment would be to manipulate the densities of such attractive species to investigate the effects on several, not just one, other species. Such an experiment could reveal which facilitation or competition is the most important interaction for pollinator attraction of plant species that co-flower at the plant community level. Amongst others, such an experiment may have implications for predictions of community-level effects of extinction or introduction of highly attractive plant species.
Bumblebees are regarded to be of large importance for pollination of both wild and cultivated crops in temperate ecosystems, and they are often more efficient than, for example, honeybees (Goulson 2003). We do not know of any study that has compared the relative importance of different pollinator groups for different plant species’ reproduction on the community level. However, results from studies of single species may suggest that bumblebees are the most important pollinator for many plant species (e.g. Meynie and Bernard 1997, Aizen 2001, Kandori 2002). Moreover, our results show that bumblebees perform a large fraction of visits to those species that are visited by more pollinator groups (see Table 1). Furthermore, the results for all insects resemble those for bumblebees, indicating that species traits important to bumblebees contribute substantially to determining the visitation frequency of all insects.
The plant traits related to bumblebee- and fly-visitation frequencies in our community have a poor fit to patterns predicted by classical pollination syndromes. Only flies had a visitation frequency that significantly depended on categorical floral traits, preferring species with actinomorphic and open flowers, a pattern consistent with the fly-pollination syndrome (Fægri and van der Pijl 1979, see also Pellmyr 2002 and references therein). The most frequently visited plant species by flies; P. erecta, F. ulmaria and Galium verum, all have actinomorphic and open flowers. Bumblebee visitation frequencies, on the other hand, did not differ between levels of categorical floral traits. Consequently, our results suggest that the classical pollination syndromes do not necessarily reflect the visitation pattern of a whole group of pollinators. For example, among bumblebee species there is considerable variation in important morphological traits, such as tongue length, that determine if individuals of a species choose to visit a plant (Goulson 2003). The species most frequently visited by bumblebees in our study community F. ulmaria, T. pratense, and P. vulgaris, are species that have differences in categorical floral traits, for example flower colour. McCall and Primack (1992) found that bees in a woodland-meadow community preferred plant species with purple flowers, while flies preferred species with open flowers, thus partly confirming visitation patterns predicted by pollination syndromes. However, in tundra and fynbo, other floral traits appeared to be important determinants of floral preference by bees and flies (McCall and Primack 1992). Our results suggest that classical pollination syndromes represent a too simple picture of the interactions that occur between pollinators and plant species at the community level. An alternative to a taxonomical division of pollinators is a separation of pollinators into functional groups (e.g. Fenster et al. 2004), implying that for example short- and long-tongued bumblebees are partitioned into separate groups. Furthermore, even if categorical traits (here: flower symmetry and shape) may be important for visitation frequency, our results indicate that their importance has been exaggerated relative to continuous traits (here: display area and/or floral density) in understanding the variation in visitation frequency among plant species within a community.
References
Aizen MA (2001) Flower sex ratio, pollinator abundance, and the seasonal pollination dynamics of a protandrous plant. Ecology 82:127–144
Bosch J, Retana J, Cerda X (1997) Flowering phenology, floral traits and pollinator composition in a herbaceous Mediterranean plant community. Oecologia 109:583–591
Campbell DR, Motten AF (1985) The mechanism of competition for pollination between two forest herbs. Ecology 66:554–563
Caruso CM (1999) Pollination of Ipomopsis aggregata (Polemoniaceae): Effects of intra- vs. interspecific competition. Am J Bot 86:663–668
Conner JK, Rush S (1996) Effects of flower size and number on pollinator visitation to wild radish, Raphanus raphanistrum. Oecologia 105:509–516
Cresswell JE, Galen C (1991). Frequency dependent selection and adaptive surfaces for floral character combinations: the pollination of Polemonium viscosum. Am Nat 138:1342–1353
Dicks LV, Corbet SA, Pywell RF (2002) Compartmentalization in plant-insect flower visitor webs. J Anim Ecol 71:32–43
DNMI (2003) Meteorologisk institutt, Oslo. http://met.no/observasjoner/sogn_og_fjordane/2003/juli/index.html?month. Cited 3 Jan 2005
Duffield GE, Gibson RC, Gilhooly PM, Hesse AJ, Inkley CR, Gilbert FS, Barnard CJ (1993) Choice of flowers by foraging honey-bees (Apis mellifera): possible morphological cues. Ecol Entomol 18:191–197
Dupont YL, Hansen DM, Olesen JM (2003) Structure of a plant-flower-visitor network in the high-altitude sub-alpine desert of Tenerife, Canary Islands. Ecography 26:301–310
Elberling H, Olesen JM (1999) The structure of a high latitude plant-flower visitor system: the dominance of flies. Ecography 22:314–323
Faegri K, van der Pijl L (1979) The principles of pollination ecology, 3rd edn. Pergamon, Oxford
Feinsinger P (1987) Effects of plant-species on each others pollination: is community structure influenced. Trends Ecol Evol 2:123–126
Fenster CB, Armbruster WS, Wilson P, Dudash MR, Thomson JD (2004) Pollination syndromes and floral specialisation. Annu Rev Ecol Evol Syst 35:375–403
Fremstad E (1997) Vegetasjonstyper i Norge. NINA, Trondheim
Glantz SA, Slinker BK (2001) Primer of applied regression and analysis of variance. McGraw Hill, New York
Goulson D (2003) Bumblebees: behaviour and ecology. Oxford University Press, Oxford
Goulson D, Stout JC, Hawson SA, Allen JA (1998) Floral display size in comfrey, Symphytum officinale L. (Boraginaceae): relationships with visitation by three bumblebee species and subsequent seed set. Oecologia 113:502–508
Herrera J (1988) Pollination relationships in southern Spanish Mediterranean shrublands. J Ecol 76:274–287
Inoue K, Maki M, Masuda M (1995) Different responses of pollinating bees to size variation and sexual phases in flowers of Campanula. Ecol Res 10:267–273
Inouye DW, Pyke GH (1988) Pollination biology in the Snowy Mountains of Australia: comparisons with montane Colorado, USA. Aust J Ecol 13:191–210
Jordano P (1987) Patterns of mutualistic interactions in pollination and seed dispersal: connectance, dependence asymmetries, and coevolution. Am Nat 129:657–677
Kandori I (2002) Diverse visitors with various pollinator importance and temporal change in the important pollinators of Geranium thunbergii (Geraniaceae). Ecol Res 17:283–294
Lack AJ (1982) The ecology of flowers of chalk grassland and their insect pollinators. J Ecol 70:773–790
Laverty TM (1992) Plant interactions for pollinator visits: a test of the magnet species effect. Oecologia 89:502–508
Lid J, Lid DT (1994) Norsk flora. Samlaget, Oslo
McCall C, Primack RB (1992) Influence of flower characteristics, weather, time of day, and season on insect visitation rates in three plant communities. Am J Bot 79:434–442
Medel R, Botto-Mahan C, Kalin-Arroyo M (2003) Pollinator-mediated selection on the nectar guide phenotype in the Andean Monkey flower, Mimulus luteus. Ecology 84:1721–1732
Memmott J (1999) The structure of a plant–pollinator food web. Ecol Lett 2:276–280
Meynie S, Bernard R (1997) Pollinator efficiency of some insects in relation to wild species populations of Helianthus L. Agronomie 17:43–51
Ohashi K, Yahara T (1998) Effects of variation in flower number on pollinator visits in Cirsium purpuratum (Asteraceae). Am J Bot 85:219–224
Olesen JM, Jordano P (2002) Geographic patterns in plant–pollinator mutualistic networks. Ecology 83:2416–2424
Pellmyr O (2002) Pollination by animals. In: Herrera CM, Pellmyr O (eds) Plant-animal interactions: an evolutionary approach. Blackwell, Oxford, pp 157–184
Petanidou T, Vokou D (1993) Pollination ecology of Labiatae in a phryganic (east Mediterranean) ecosystem. Am J Bot 80:892–899
Pickering CM (2001) Size and sex of floral displays affect insect visitation rates in the dioecious Australian alpine herb, Aciphylla glacialis. Nord J Bot 21:401–409
Pleasants JM (1980) Competition for bumblebee pollinators in Rocky Mountain plant communities. Ecology 61:1446–1459
Schemske DW, Willson MF, Melampy MN, Miller LJ, Verner L, Schemske KM, Best LB (1978) Flowering ecology of some spring woodland herbs. Ecology 59:351–366
Stanton ML, Preston RE (1988) Ecological consequences and phenotypic correlates of petal size variation in wild radish, Raphanus sativus (Brassicaceae). Am J Bot 75:528–539
Thomson JD (1978) Effects of stand composition on insect visitation in two-species mixtures of Hieracium. Am Midl Nat 100:431–440
Thomson JD (1981) Spatial and temporal components of resource assessment by flower-feeding insects. J Anim Ecol 50:49–59
Thomson JD (1982) Patterns of visitation by animal pollinators. Oikos 39:241–250
Totland O (1993) Pollination in alpine Norway: flowering phenology, insect visitors, and visitation rates in two plant-communities. Can J Bot 71:1072–1079
Totland O, Matthews I (1998) Determinants of pollinator activity and flower preference in the early spring blooming Crocus vernus. Acta Oecol 19:155–165
Waser NM, Fugate ML (1986) Pollen precedence and stigma closure: a mechanism of competition for pollination between Delphinium nelsonii and Ipomopsis aggregata. Oecologia 70:573–577
Waser NM, Chittka L, Price MV, Williams NM, Ollerton J (1996) Generalization in pollination systems, and why it matters. Ecology 77:1043–1060
Wilkinson L (2003) SYSTAT 10.0. SPSS Inc, Chigaco
Zar JH (1999) Biostatistical analysis. Prentice Hall, New Jersey
Acknowledgements
We thank Sondre Eikås, Vegard Eldholm, Peter Greve, Maria Collett Knagenhjelm, Torkjell Solbraa, and Knut Fredrik φ i for their invaluable field assistance, Luc Boeke for statistical advices, and Anne-Line Bjerknes, Andreas Jürgens, Anders Nielsen and two anonymous reviewers for very constructive comments on the manuscript. We also thank Christen Knagenhjelm for letting us use the meadow in Rudsviki. The study is financed by the Norwegian Research Council through the Biodiversity-program (project number 154185/720).
Author information
Authors and Affiliations
Corresponding author
Additional information
Communicated by Christian Koerner
Rights and permissions
About this article
Cite this article
Hegland, S.J., Totland, Ø. Relationships between species’ floral traits and pollinator visitation in a temperate grassland. Oecologia 145, 586–594 (2005). https://doi.org/10.1007/s00442-005-0165-6
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00442-005-0165-6