
Abstract
In the past years, a number of studies have used experimental plant communities to test if biodiversity influences ecosystem functioning such as productivity. It has been argued, however, that the results achieved in experimental studies may have little predictive value for species loss in natural ecosystems. Studies in natural ecosystems have been equivocal, mainly because in natural ecosystems differences in diversity are often confounded with differences in land use history or abiotic parameters. In this study, we investigated the effect of plant diversity on ecosystem functioning in semi-natural grasslands. In an area of 10×20 km, we selected 78 sites and tested the effects of various measures of diversity and plant community composition on productivity. We separated the effects of plant diversity on ecosystem functioning from potentially confounding effects of community composition, management or environmental parameters, using multivariate statistical analyses. In the investigated grasslands, simple measures of biodiversity were insignificant predictors of productivity. However, plant community composition explained productivity very well (R2=0.31) and was a better predictor than environmental variables (soil and site characteristics) or management regime. Thus, complex measures such as community composition and structure are important drivers for ecosystem functions in semi-natural grasslands. Furthermore, our data show that it is difficult to extrapolate results from experimental studies to semi-natural ecosystems, although there is a need to investigate natural ecosystems to fully understand the relationship of biodiversity and ecosystem functioning.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
The consequences of the observed worldwide loss in biodiversity for ecosystem functioning are a hotly debated topic in ecological research (Aarssen 1997; Huston 1997; Grime 1998; Hector et al. 2000; Troumbis 2001; Wardle 2001; Loreau et al. 2002). In most of the recent laboratory and field experiments, the effect of biodiversity on productivity was tested in artificial plant communities where different diversity levels were established by drawing plant species from a random species pool. Generally, these experiments have shown an asymptotic increase in productivity with increasing number of species (Naeem et al. 1994, 1996; Tilman et al. 1996, 1997; Hooper and Vitousek 1997; Hector et al. 1999). The observed patterns have largely been attributed to niche complementarity (Tilman et al. 1996; Hector 1998; Loreau 1998; Loreau and Hector 2001). Niche complementarity predicts that an increase in species richness will lead to a more efficient use of available resources and thus increased productivity. Several authors have questioned the applicability of the findings of these studies to natural ecosystems, where biodiversity will be insignificant compared to the overwhelming influences of environmental and anthropogenic factors on ecosystem functioning (Grime 1997; Wardle et al. 1997; Huston and McBride 2002). It has also been argued that the results achieved in experimental studies may have little predictive value for species loss in natural ecosystems where species extinction is not random but directed (Grime 2002; Diaz et al. 2003). In recent reviews, Chapin (2000) and Loreau (2001) have therefore suggested that the scope of the study of biodiversity and ecosystem functioning should be expanded to natural ecosystems.
In studies of natural ecosystems, biodiversity has traditionally been viewed as a response rather than a predictive variable. Diversity has been hypothesized to peak at intermediate levels of productivity and decrease at high or low productivity, and a number of data sets support this theory (Al-Mufti et al. 1977; Rosenzweig and Abramsky 1993; Grace 1999; Waide et al. 1999; Mittelbach et al. 2001). Although productivity per se has historically been considered as the main variable driving the productivity–diversity relationship, recent studies have demonstrated that this pattern can arise from covariation of productivity with other abiotic or management factors, illustrating the complexity of environmental regulation of species diversity in natural communities (Gough et al. 1994; Chase and Leibold 2002; Schaffers 2002; Fukami and Morin 2003; Rajaniemi 2003).
This traditional view of biodiversity, where high productivity results in low diversity, seems inconsistent with the experimental results, where high diversity results in increased productivity. Both approaches are, however, complementary rather than contradictory (Loreau 2000; Huston and McBride 2002; Schmid 2002). While the traditional approach attempts to identify the spatial variation of diversity across environmental gradients, the experimental approach tries to determine the consequences of species loss in a given system where all environmental factors remain constant. Testing the effect of biodiversity on productivity in complex natural communities would therefore require the control of environmental gradients, which is difficult because of the large number of variables that influence diversity (Wardle 2001). An alternative is to apply multivariate models that permit control of environmental variables statistically and detect direct and indirect effects of diversity and environmental variables on ecosystem functions.
While most experimental studies and the traditional analyses of the productivity–diversity relationship have focused on species richness as a measure of plant biodiversity, different aspects of diversity should be considered when the effects on productivity are investigated. Several studies have, for example, shown that the number of plant functional groups, functional diversity or evenness influenced productivity more strongly than species richness (Tilman et al. 1997; Wilsey and Potvin 2000; Diaz and Cabido 2001; Spehn et al. 2002; Petchey et al. 2004). Also, it has been shown that the presence of one or a few dominant species with strong ecosystem effects is likely to mask simple relationships of species richness and productivity (Chapin et al. 1997; Huston 1997; Hooper and Vitousek 1998; Aarssen 2001; Huston and McBride 2002). Consequently, the effect of specific species and community composition need to be considered if diversity effects on ecosystem functions are tested in a semi-natural ecosystem.
In this study, we aimed to determine the effect of different aspects of plant diversity on productivity in semi-natural managed grasslands. Specifically, we tested: (1) the direct effects of plant diversity (using different indices) and community composition on productivity as well as, (2) the direct and indirect effects of environmental parameters such as soil properties, management and site characteristics on both diversity and productivity. In a final step, we applied a path analysis to distinguish between direct and indirect effects of environmental variables on plant diversity and productivity.
Materials and methods
Study area and study sites
The study was conducted in the Thüringer Schiefergebirge/Frankenwald, a plateau-like mountain range at the Thuringian/Bavarian border in central Germany, which reaches a maximum height of 870 m. The bedrock material in the investigated area produces a carbonate-free, nutrient-poor soil. Average annual precipitation is above 1,000 mm with a slight summer maximum. Annual average temperature is 5°C. Before human settlement in the middle ages, montane spruce–fir–beech forests formed the natural vegetation in the area. Thereafter, much of the forest was converted into an agricultural landscape, with a high proportion of different montane hay meadow and pasture grasslands (Geranio–Trisetetum, Knapp ex Oberd. 1957) (Hundt 1964).
In 2001, seventy-eight managed grasslands were studied. All sites were located between 500 and 840 m altitude on high elevation plateaus of the mountain range, so that orographic and edaphic factors were relatively comparable among sites. To qualify for the study, grasslands had to be free of woody plants as a sign of recent management, and had to be uncut or ungrazed by the time of the study. The minimum size of a site had to be 1 ha. In each site, a 2×2 m plot was established at a distance of about 50 m from other habitats or roads. In each plot, plant species richness, percent cover, aboveground biomass (following used as surrogate for productivity), soil pH, soil moisture, soil C:N and plant available soil nutrients were determined.
Sampling of environmental, productivity and vegetation data
During a field campaign (28 May–9 June 2001), two soil cores (4.5×10 cm) were taken within each plot. Soil of each core was sieved to 2 mm. One part of the soil was extracted with 1 M KCl on the same day of sampling. KCl extracts were frozen at −20°C and later analysed using a continuous flow analyser (SAN Plus; Skalar, Erkelenz, Germany) for NH +4 and NO −3 and an ICP-AES (Optima 3300 DV; Perkin-Elmer, Norwalk, Conn.) for Ca2+. The remaining soil was dried at 35°C and extracted using a 1 M calcium–acetate–lactate (CAL) solution. CAL extracts were analysed with ICP-AES (Optima 3300 DV) for P, K+, Mg2+ and SO 2−4 . Soil pH was measured in a water extract. For C:N, total N (Ntot) and total C (Ctot) determinations, dry soil was ground and analysed with an Element Analyser (Vario EL II; Elementar, Hanau, Germany). Soil moisture (vol%) was measured in the field using time domain reflectometry (ThetaProbe; Delta-T Devices, Cambridge) at four different locations within each plot.
All plant species within the 2×2 m plot were identified to the species level and the percent cover of each species was estimated visually. We used aboveground standing biomass as a surrogate for productivity in this study. Biomass was harvested at peak standing biomass in two 25×50 cm rectangles 2 cm above ground in each plot. The plant material was dried at 60°C for 48 h and the dry weight determined thereafter.
For an orographic characterization of the sites, geographic position (GPS coordinates), altitude, exposition, inclination as well as the distance to the next woodland habitat was recorded. Based on the mean exposition and inclination, the mean potential direct solar insolation, was calculated for each site [Homann, Schumacher and Perner, unpublished software program based on an algorithm by Volz (1959)].
Information about the management regimes of the investigated sites was collected using a standardized questionnaire. Each farmer was interviewed in December 2001 or January 2002 about the following topics: (1) approximate age of grassland, (2) mowing intensity (number and timing of cuts, separately for 2001 and the past 10 years), (3) grazing intensity (animal species, grazing density, timing, duration, separately for 2001 and the past 10 years), (4) fertilization (type of fertilizer, amount of fertilizer applied, timing of fertilization, separately for 2001 and for the past 10 years). For statistical analysis, management data were converted into numeric values. For the cutting regime, dates of cutting were weighted to reflect that early cuts have a stronger effect on plant diversity than later cuts (Klapp 1971): a value of 3 was assigned to cuts in early summer (prior to 15 June), a value of 2 for cuts between 15 June and 1 September, and a value of 1 for cuts after 1 September. In case grasslands were cut more than once during the season, the numbers assigned to each cut were summed to yield a single value for the statistical analysis. For grazing, values between 1 and 4 were assigned. For each value, grazing time, intensity and frequency as well as type of grazing animal (cattle, cow, horse and sheep) were considered. High “grazing values” reflect a high grazing impact on the grassland (Klapp 1971). For fertilization, values between 1 and 9 were assigned, with low values for “light-impact fertilization” and high values for “high-impact fertilization”. Values between 1 and 4 were assigned to sites that received dung from grazing animals, with higher stocking densities resulting in larger values. Values of 5, 6 and 7 were assigned to sites with applications of <50, 50–100 kg ha−1 year−1 and >100 kg ha−1 year−1 of industrial NPK fertilizer, respectively. Sites treated with solid manure were assigned a value of 8, while liquid manure application was assigned a value of 9.
Statistical analysis
To improve normality of variances and avoid distortions plant species cover data were square-root transformed before analyses. In a first step, the number of edaphic site characteristics and management variables was reduced using principal component analysis [PCA, CANOCO (ter Braak and Smilauer 2002)]. This procedure summarizes the information of the variables as four major axes of a standardized PCA. Since PCA axes are by definition orthogonal and independent of each other, this procedure creates composite independent variables and avoids the danger of spurious correlations (i.e. multicollinearity). PCAs were performed separately for edaphic, management and orographic parameters. From each PCA, the axes explaining most of the variance (but no more than the first four) were extracted resulting in new PCA-derived variables. These PCA-derived variables were used in all consecutive analyses as independent parameters. Very little information was lost by this procedure since the extracted axes explained most of the total variance contained in the original parameter groups.
From the plant cover data, we calculated plant species richness, effective diversity (heterogeneity or exponential Shannon–Wiener), and Camargo’s evenness (calculation algorithms see Krebs 1999). To analyse the compositional differences among the plant communities of the 78 studied grassland sites, non-metric multidimensional scaling (NMDS) ordination techniques were applied using the program PC-ORD (McCune and Mefford 1997). NMDS is an iterative search for a ranking and placement of n entities (samples) in k dimensions (ordination axes) that minimizes the stress of the k-dimensional configuration. The “stress” value is a measure of departure from monotonicity in the relationship between the dissimilarity (distance) in the original p-dimensional space and in the reduced k-dimensional ordination space (Clarke 1993). As a distance measure, the Bray-Curtis coefficient was used (also known as Sørensen or Czekanowski coefficient), which is one of the most robust measures for this purpose (Faith et al. 1987). NMDS ordination was based on square-root-transformed cover data. To analyse which of the species are mainly responsible for the compositional changes within the investigated plant communities (along the extracted NMDS axes), we performed linear regressions of species cover versus the scores of the NMDS axes.
We used least squares linear regressions to analyse the effects of plant diversity and community composition (species richness, effective diversity, Camargo’s evenness, NMDS axes) on productivity. Thereafter, we performed multiple regression analyses to test the effect of the same diversity measures on productivity in combination with different sets of environmental parameters. The multiple regression analyses were performed separately for each diversity measure and one of the PCA constructed parameter groups, edaphic variables, site characteristics and management variables.
In a next step, we used multiple stepwise regressions to test whether the PCA-derived variables were significant predictors for plant diversity measures that were significantly correlated with productivity. In addition, we tested the predictive value of the PCA-derived variables for productivity itself. For each dependent variable (diversity measures and productivity), separate regression models were calculated for each parameter group, edaphic variables, site characteristics and management variables, respectively. Regression analyses were performed using SPSS version 11 (SPSS 2001).
In a final path analysis, we used structural equation modelling [AMOS version 4.0 (Arbuckle and Wothke 1995–1999)] to test the hypothesis that environmental and management parameters influence productivity both directly or indirectly by affecting plant species composition in the sites. Starting from the most complex model that included all significant variables from the multiple regression analyses, model simplification was based on the significance of the regression weights. The competing models were compared by bootstrapping each model 1,000 times and using the Akaike information criterion (AIC), the Browne-Cudeck criterion, and the consistent Akaike information criterion (Arbuckle and Wothke 1995–1999). Based on Schmid (2002), the model assumed that productivity is a response variable only, having no effect on environmental variables or species composition.
Results
Plant diversity and environmental parameters
Plant species richness in the 78 sites varied between 8 and 33 species 4 m−2, effective diversity between 4.3 and 19.4, and Camargo’s evenness between 0.28 and 0.54. Mean aboveground plant biomass was 359 g dry weight (dw) m−2 (range 125–610 gdw m−2) (Table 1). For range and descriptive statistics of plant available soil nutrients and site characteristics see Table 1.
The majority of the investigated grasslands were cut for haymaking (58%). Fewer sites were grazed (12%) or cut and grazed (27%). One site was not managed in the year of the investigation. In most cases, cutting occurred once a year (n=54), although some sites were cut twice (n=18) or even 3 times (n=3) per season. Grazing occurred mainly with cows or cattle (n=17), with only some sites grazed by horses (n=8) or sheep (n=4). Only 22% of all sites were fertilized. Applied fertilizer included industrial NPK, dung or liquid manure. Recent management (2001) was closely correlated to the management in the last 10 years (P<0.001).
Aggregation of environmental data
Four axes were extracted as independent variables from the PCA of the edaphic parameters and labelled soil1–soil4 (Table 2). These four axes explained 77.2% of the total variance of all edaphic parameters. Table 2 illustrates the species loadings of the individual axes. Soil1 was mainly related to pH, SO 2+4 , Ca2+, Mg2+, Nmin and soil moisture. Except for the last two variables, all of these parameters are in some manner related to soil acidity. Soil2 was correlated with total soil C and N as well as C:N. Soil3 was correlated mainly with K+, while soil4 was strongly correlated with extractable P (Table 2).
The PCA on the site characteristics extracted four axes (site1–site4), explaining 100% of the total variation. Site1 was mainly correlated with inclination and potential insolation, site2 with altitude and potential insolation, site3 with altitude and distance to next habitat and site4 with inclination (Table 2).
For the PCA on management variables, the first two axes explained 86.9% of the total variation (manage1 and manage2); Reflecting the close correlations between current and past management, Manage1 was mainly correlated with present and past grazing and fertilization regimes while manage2 was mainly correlated with present and past cutting as well as fertilization regimes. Axes 3 and 4 were ignored in further analyses for their low explanatory value (Table 2).
Plant community composition
Based on plant species composition, the 78 investigated grasslands can be separated into two overlapping groups, a more productive Geranio–Trisetetum alopecuretosum type and a less productive Geranio–Trisetetum nardetosum type (Fig. 1). Both grassland types are closely related and belong to the same phytosociological association [Geranio (Sylvatici)–Trisetetum, Knapp ex Oberd. 1957] (Hundt 1964). Plant species richness was not correlated with species composition. NMDS showed that a two-dimensional solution was sufficient to achieve low stress values (first axis/dimension=23.3, second axis/dimension=16.1, R2=0.73) to explain plant composition (Fig. 1). For further analyses, we therefore used the scores of the first two axes as parameters for plant community composition (NMDS1 and NMDS2). Several dominant plant species showed a strong positive or negative relation with the NMDS axes (Table 3).

Non-metric multidimensional scaling (NMDS) ordination of the 78 montane grassland sites (minimum stress values first axis/dimension=23.3, second axis/dimension=16.1, R2=0.73). Increasing symbol size Increasing plant species richness (range 8–33), empty circles more productive Geranio-Trisetetum alopecuretosum sites, filled circles Geranio-Trisetetum nardetosum sites
The relationship between diversity and productivity
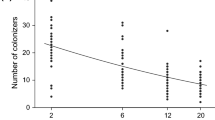
The diversity measures, plant species richness, effective diversity and Camargo’s evenness, had no significant effect on productivity (aboveground standing biomass) when tested in linear regressions (Fig. 2). However, community composition represented as NMDS1 and NMDS2 was significantly related to productivity. While NMDS1 was negatively but only weakly related to productivity (R2=0.05, P=0.050), NMDS2 showed a positive and highly significant effect on productivity (R2=0.31, P<0.001) (Fig. 2). The explanatory value of the five diversity measures increased only marginally when environmental variables were included in the analysis using multiple regressions (data not shown).

Relationships of different plant diversity measures and community composition to productivity in semi-natural grasslands
Environmental parameters, plant community composition and productivity
When tested in multiple stepwise regression models, 75.0% of the total variability in NMDS2 was explained by the edaphic parameters soil1, soil4 and soil2 (Table 4). In the regressions with either site characteristics or management parameters as independent variables, the PCA-derived parameters site3, and manage1 and manage2 explained only 6.5 and 38.8% of the variation in NMDS2 scores, respectively (Table 4). For productivity, the PCA-derived variables had little explanatory power in multiple regression analyses. Only 12% of variation in productivity was explained by soil1 and only 18% by manage2 (Table 4). None of the site characteristics emerged as a significant predictor variable (Table 4).
Structural equation modelling (path analysis)
For structural equation modelling, we only considered the variable NMDS2 as diversity or composition measure because this was the only variable which was correlated with productivity. The initial model tested in AMOS consisted of all PCA-derived edaphic, management and site parameters that were significantly correlated with either productivity or NMDS2 in the multiple regression analyses (Table 4). Thus, the initial model included the environmental variables soil1, soil2, soil4, site3, manage1 and manage2 (Fig. 3a). Productivity was assumed to be dependent on NMDS2 so the model included only a path from NMDS2 to productivity and not vice versa (testing a model in which paths were drawn in both directions resulted in non-significant regression weights for both paths). This initial model was simplified by removing variables and paths according to the measures of fit (Table 5). All of the tested models were significant. The model excluding the variables site3 and manage1 but including the regressions of the variables soil1 and manage2 on productivity resulted in the best AIC value and the highest explained variability for the variables productivity and NMDS2 (Table 5, Fig. 3b). Thus, environmental variables influenced productivity, both directly as well as indirectly via an effect on plant species composition.

Structural equation modelling (cf. Table 5). a Initial model. Single-headed arrows indicate paths. Double-headed arrows show covariances that were included in the model based on modifications proposed by AMOS (procedure modification indices). The exogenous unobserved variables err1 and err2 account for the unexplained error in the estimation of NMDS2 and productivity, respectively. Their regression weights were a priori set to unity. b Standardized regression weights (along paths), correlations (along double-headed arrows) and squared multiple correlations (beside the productivity and NMDS2 boxes) for the best-fitting model C (Table 5).
Discussion
The data presented in this study illustrate that simple measures of biodiversity such as species richness are weak predictors for productivity in semi-natural grasslands. Community composition, however, explained productivity very well and was a better predictor for productivity than environmental variables and management parameters together. Nevertheless, some of the edaphic, management and site parameters showed a direct effect on productivity, although their influence was not as strong as maybe expected. More importantly, these variables influenced productivity indirectly via their influence on community composition. Our results show that complex measures such as community composition are important predictors of ecosystem functioning in semi-natural ecosystems.
The grasslands selected for this study cover a wide range of species diversity, productivity and environmental parameters, representing a good sub-sample of montane semi-natural grasslands found in central Europe (Hundt 1964). Aboveground biomass was comparable to those given in other studies where the effect of plant diversity on productivity was studied in grasslands of Europe or North America (Hector et al. 1999; Tilman et al. 2002). Although the investigated sites cover a wide range in plant species richness (from eight to 33 species, Table 1), average plant species richness of 20 species was low when compared to other semi-natural European grasslands (Baur et al. 1996). Montane grasslands such as investigated in this study are, however, generally lower in species diversity than comparable calcareous grasslands (Hundt 1964). In addition, we tested the quality of our sampling method by nested subset sampling using modified Whittaker plots in adjacent grasslands and found that the error of underestimating species richness was small (A. Kahmen, unpublished data). Consequently, the number of plant species in 2×2 m plots presents a good relative estimate of plant diversity of the investigated sites.
The relationship between productivity and plant species richness has been described to peak at intermediate levels of productivity in numerous different grassland ecosystems (Al-Mufti et al. 1977; Rosenzweig and Abramsky 1993; Grace 1999; Waide et al. 1999; Mittelbach et al. 2001). The data of our study are consistent with this finding (Fig. 2a, reversed axes). However, the hump-shaped relationship found in our study results from a line enveloping the outer-most data points rather than from a line of fitted average values as suggested by Al-Mufti et al. (1977). Our results are better explained by the theoretical model introduced by Schmid (2002) that combines the hump-shaped relationship from observational biodiversity studies with results from experimental studies where high diversity results in increased productivity. In his model, Schmid introduces site fertility as a third parameter in addition to productivity and biodiversity. The model assumes that productivity as well as biodiversity is ultimately driven by site fertility and that, given an intact species pool, species richness shows a hump-shaped relationship with productivity as suggested by Al-Mufti et al. (1977). If the species pool at a given fertility level is, however, reduced as a result of extinction or experimental manipulation, diversity and eventually productivity will drop below the ideal hump-shaped line.
We tested if declining diversity in the observed grasslands had an effect on productivity but found that simple diversity measures such as species richness, effective diversity or Camargo’s evenness showed no significant relationship with productivity (Fig. 2). The explanatory value of the simple diversity measures also did not increase when potentially confounding environmental parameters such as soil variables or management parameters were included in the model using multiple regression analyses. Interestingly, our results are not consistent with experimental studies, where an asymptotic increase in biomass with increasing plant diversity or evenness was found (Naeem et al. 1996; Hooper and Vitousek 1997; Tilman et al. 1997; Hector et al. 1999; Wilsey and Potvin 2000; Polley et al. 2003; Symstad et al. 2003). For these experimental studies, it was argued that the observed positive effects of biodiversity on productivity in experimental studies are largely due to niche complementary (Tilman et al. 1996, 2002; Hector 1998; Loreau 1998; Loreau and Hector 2001). The niche complementary effect suggests that an increasing number of species results in a more efficient resource exploitation and thus enhanced ecosystem functioning. The observed diversity effects on productivity in the experimental studies are, however, driven by very low species levels, which are not representative for natural grasslands. Tilman (2002) for example states that in his study about five species might account for the observed biodiversity effects. In contrast, the lowest diversity level in our study contained eight plant species. We therefore suggest that biodiversity effects based on niche complementary are strongest in ecosystems where diversity has dropped below a critical level such as in experimental grasslands.
In contrast to plant diversity, effective diversity or evenness, which are non-significant, it was community composition (NMDS1 and NMDS2) that had a significant effect on the productivity of the investigated grasslands (Fig. 2). Community composition (NMDS2) is correlated with several highly productive plant species (Table 3), suggesting that species with specific traits such as high competitive ability or high nutrient use efficiency may be the important drivers in the relationship of community composition and productivity. This would be analogous to results found in several experimental biodiversity studies where species composition or functional traits of specific species were a better predictor for ecosystem functioning than species richness (Hooper and Vitousek 1998; Symstad et al. 1998; Diaz and Cabido 2001; Petchey et al. 2004). The influence of species-specific traits does not, however, dismiss the so called “diversity effects” from the relationship of community composition and productivity. Tilman (2002) for example suggested that communities with complementary functional composition should be more productive than communities with equal species numbers but redundant functional composition. Also, several studies have shown that legumes facilitated increased productivity by transferring fixed nitrogen (N) to other plant species in the community (Mulder et al. 2002; Spehn et al. 2002; Scherer-Lorenzen et al. 2003). Testing the effects of functional diversity or the transfer of symbiotically fixed N was however, not, in the scope of this study. We therefore cannot exclude such diversity effects from our interpretation of species composition.
It has long been acknowledged that ecosystem functioning is influenced by biotic factors such as species-specific traits and species interactions as well as abiotic parameters such as climate, soil and disturbance. The role of biodiversity in the creation, maintenance and functioning of ecosystems has, however, only recently been addressed (Schulze and Mooney 1993; Lawton 1994; Chapin et al. 1997; Naeem 2002) and much of the recent debate about biodiversity and ecosystem functioning has focused on the relative contributions of any of these factors to the observed ecosystem processes. While in experimental studies, environmental variables have been controlled, several authors have questioned their applicability for semi-natural ecosystems in the face of overwhelming influences of extrinsic factors (Grime 1997; Wardle et al. 1997; Huston and McBride 2002). In our study, we therefore tested the individual influences of species composition (NMDS2) and environmental variables on productivity in a single structural equation model (Fig. 3a). The model reveals that community composition is the most important parameter that is directly driving productivity in the investigated grasslands. While environmental parameters and management are highly important basic factors for ecosystem functions, their influence on productivity is indirect via driving the community composition (Fig. 3b). Our model stresses that community composition of an ecosystem with its species-specific functional traits as well as its species interactions needs to be taken into account when ecosystem functioning is to be understood.
References
Aarssen LW (1997) High productivity in grassland ecosystems: effected by species diversity or productive species? Oikos 80:183–184
Aarssen LW (2001) On correlations and causations between productivity and species richness in vegetation: predictions from habitat attributes. Basic Appl Ecol 2:105–114
Al-Mufti MM, Sydes CL, Furness SB, Grime JP, Band SR (1977) Quantitative analysis of shoot phenology and dominance in herbaceous vegetation. J Ecol 65:759–791
Arbuckle JL, Wothke W (1995–1999) AMOS 4.0 user’s guide. Small Waters Cooperation, Chicaco, Ill.
Baur B et al (1996) Variation in species richness of plants and diverse groups of invertebrates in three calcareous grasslands of the Swiss Jura mountains. Rev Suisse Zool 103:801–833
ter Braak CJF, Smilauer P (2002) CANOCO reference manual and CanoDraw for Windows user’s guide: software for canonical community ordination (version 4.5). Microcomputer Power, Ithaca, N.Y.
Chapin FS et al (1997) Biotic control over the functioning of ecosystems. Science 277:500–504
Chapin FS et al (2000) Consequences of changing biodiversity. Nature 405:234–242
Chase JM, Leibold MA (2002) Spatial scale dictates the productivity–biodiversity relationship. Nature 416:427–430
Clarke KR (1993) Nonparametric multivariate analyses of changes in community structure. Aust J Ecol 18:117–143
Diaz S, Cabido M (2001) Vive la difference: plant functional diversity matters to ecosystem processes. Trends Ecol Evol 16:646–655
Diaz S, Symstad AJ, Chapin FS, Wardle DA, Huenneke LF (2003) Functional diversity revealed by removal experiments. Trends Ecol Evol 18:140–146
Faith DP, Minchin PR, Belbin L (1987) Compositional dissimilarity as a robust measure of ecological distance. Vegetatio 69:57–68
Fukami T, Morin PJ (2003) Productivity–biodiversity relationships depend on the history of community assembly. Nature 424:423–426
Gough L, Grace JB, Taylor KL (1994) The relationship between species richness and community biomass—the importance of environmental variables. Oikos 70:271–279
Grace JB (1999) The factors controlling species density in herbaceous plant communities: an assessment. Perspect Plant Ecol Evol Syst 2:1–28
Grime JP (1997) Biodiversity and ecosystem function: the debate deepens. Science 277:1260–1261
Grime JP (1998) Benefits of plant diversity to ecosystems: immediate, filter and founder effects. J Ecol 86:902–910
Grime JP (2002) Declining plant diversity: empty niches or functional shifts? J Veg Sci 13:457–460
Hector A (1998) The effect of diversity on productivity: detecting the role of species complementarity. Oikos 82:597–599
Hector A et al (1999) Plant diversity and productivity experiments in European grasslands. Science 286:1123–1127
Hector A, et al. (2000) No consistent effect of plant diversity on productivity: response. Science 289:1255a
Hooper DU, Vitousek PM (1997) The effects of plant composition and diversity on ecosystem processes. Science 277:1302–1305
Hooper DU, Vitousek PM (1998) Effects of plant composition and diversity on nutrient cycling. Ecol Monogr 68:121–149
Hundt R (1964) Die Bergwiesen des Harzes, Thüringer Waldes und Erzgebirges. Fischer, Jena
Huston MA (1997) Hidden treatments in ecological experiments: re-evaluating the ecosystem function of biodiversity. Oecologia 110:449–460
Huston MA, McBride AC (2002) Evaluating the relative strengths of biotic versus abiotic controls on ecosystem processes. In: Loreau M, Naeem S, Inchausti P (eds) Biodiversity and ecosystem functioning. Oxford University Press, New York, pp 47–60
Klapp E (1971) Wiesen und Weiden - Eine Grünlandlehre. Parey, Berlin
Krebs CJ (1999) Ecological methodology. Addison–Welsey, Menlo Park
Lawton JH (1994) What do species do in ecosystems. Oikos 71:367–374
Loreau M (1998) Separating sampling and other effects in biodiversity experiments. Oikos 82:600–602
Loreau M (2000) Biodiversity and ecosystem functioning: recent theoretical advances. Oikos 91:3–17
Loreau M et al (2001) Biodiversity and ecosystem functioning: current knowledge and future challenges. Science 294:804–808
Loreau M, Hector A (2001) Partitioning selection and complementarity in biodiversity experiments. Nature 412:72–76
Loreau M, Naeem S, Inchausti P (2002) Biodiversity and ecosystem functioning: synthesis and perspectives. Oxford University Press, Oxford
McCune B, Mefford MJ (1997) PC-ORD. Multivariate analysis of ecological data. Version 3.0. MjM Software Design, Glenden Beach
Mittelbach GG et al (2001) What is the observed relationship between species richness and productivity? Ecology 82:2381–2396
Mulder CPH, Jumpponen A, Hogberg P, Huss-Danell K (2002) How plant diversity and legumes affect nitrogen dynamics in experimental grassland communities. Oecologia 133:412–421
Naeem S (2002) Ecosystem consequences of biodiversity loss: the evolution of a paradigm. Ecology 83:1537–1552
Naeem S, Thompson LJ, Lawler SP, Lawton JH, Woodfin RM (1994) Declining biodiversity can alter the performance of ecosystems. Nature 368:734–737
Naeem S, Hakansson K, Lawton JH, Crawley MJ, Thompson LJ (1996) Biodiversity and plant productivity in a model assemblage of plant species. Oikos 76:259–264
Petchey OL, Hector A, Gaston KJ (2004) How do different measures of functional diversity perform? Ecology 85:847–857
Polley HW, Wilsey BJ, Derner JD (2003) Do species evenness and plant density influence the magnitude of selection and complementarity effects in annual plant species mixtures? Ecol Lett 6:248–256
Rajaniemi TK (2003) Explaining productivity-diversity relationships in plants. Oikos 101:449–457
Rosenzweig ML, Abramsky Z (1993) How are diversity and productivity related? In: Ricklefs RE, Schluter D (eds) Species diversity in ecological communities. University of Chicago Press, Chicago, Ill., pp 52–65
Schaffers AP (2002) Soil, biomass, and management of semi-natural vegetation—Part II. Factors controlling species diversity. Plant Ecol 158:247–268
Scherer-Lorenzen M, Palmborg C, Prinz A, Schulze ED (2003) The role of plant diversity and composition for nitrate leaching in grasslands. Ecology 84:1539–1552
Schmid B (2002) The species richness-productivity controversy. Trends Ecol Evol 17:113–114
Schulze E-D, Mooney HA (1993) Biodiversity and ecosystem function. Springer, Berlin Heidelberg New York
Spehn EM et al (2002) The role of legumes as a component of biodiversity in a cross-European study of grassland biomass nitrogen. Oikos 98:205–218
SPSS Inc. (2001) SPSS Base 11.0 user’s guide. SPSS, Chicago, Ill.
Symstad AJ et al (2003) Long-term and large-scale perspectives on the relationship between biodiversity and ecosystem functioning. Bioscience 53:89–98
Symstad AJ, Tilman D, Willson J, Knops JMH (1998) Species loss and ecosystem functioning: effects of species identity and community composition. Oikos 81:389–397
Tilman D, Wedin D, Knops J (1996) Productivity and sustainability influenced by biodiversity in grassland ecosystems. Nature 379:718–720
Tilman D, Knops J, Wedin D, Reich P, Ritchie M, Siemann E (1997) The influence of functional diversity and composition on ecosystem processes. Science 277:1300–1302
Tilman D, Knops J, Wendin D, Reich P (2002) Plant diversity and composition: effects on productivity and nutrient dynamics of experimental grasslands. In: Loreau M, Naeem S, Inchausti P (eds) Biodiversity and ecosystem functioning. Oxford University Press, Oxford, pp 21–35
Troumbis AY (2001) No observational evidence for diversity-enhancing productivity in Mediterranean shrublands? A reply to Wardle. Oecologia 129:622–623
Volz F (1959) Globalstrahlung auf geneigten Hängen. Meteorol Rundsch 11:132–135
Waide RB et al (1999) The relationship between productivity and species richness. Annu Rev Ecol Syst 30:257–300
Wardle DA (2001) No observational evidence for diversity enhancing productivity in Mediterranean shrublands. Oecologia 129:620–621
Wardle DA, Zackrisson O, Hornberg G, Gallet C (1997) The influence of island area on ecosystem properties. Science 277:1296–1299
Wilsey BJ, Potvin C (2000) Biodiversity and ecosystem functioning: Importance of species evenness in an old field. Ecology 81:887–892
Acknowledgements
This study was supported by grant LC0013 in the BIOLOG program of the German Federal Ministry for Education and Research (BMBF). We are grateful to Carla Wytrykush and Nidal Odat for help in the field as well as Elke Panschitz for assistance in the laboratory. We thank in particular Michael Scherer-Lorenzen for valuable comments on the manuscript.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Kahmen, A., Perner, J., Audorff, V. et al. Effects of plant diversity, community composition and environmental parameters on productivity in montane European grasslands. Oecologia 142, 606–615 (2005). https://doi.org/10.1007/s00442-004-1749-2
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00442-004-1749-2