
Abstract
Stem cells in mammary tissue have been well characterised by using the mammary stem cell marker, cytokeratin (CK) 5 and the mature epithelial markers CK14, CK18 and CK19. As these markers have never been reported in cells from breastmilk, the aim of this study has been to determine whether mammary stem cells are present in expressed human breastmilk. Cultured cells from human breastmilk were studied by using immunofluorescent labelling and reverse transcription/polymerase chain reaction (RT-PCR). We found a heterogeneous population of cells with differential expression of CK5, CK14, CK18 and CK19. Further, by using the multipotent stem cell marker, nestin, we identified cells in culture that were positive only for nestin or double-positive for CK5/nestin, whereas no co-staining was observed for CK14, CK18 and CK19 with nestin. When cells isolated from breastmilk were analysed by using RT-PCR prior to culture, only nestin and CK18 were detected, thereby indicating that breastmilk contained differentiated epithelial and putative stem cells. Furthermore, fluorescence-activated cell-sorting analysis demonstrated, in breastmilk, a small side-population of cells that excluded Hoechst 33342 (a key property of multipotent stem cells). When stained for nestin, the cells in the side-population were positive, whereas those not in the side-population were negative. The presence of nestin-positive putative mammary stem cells suggests that human breastmilk is a readily available and non-invasive source of putative mammary stem cells that may be useful for research into both mammary gland biology and more general stem cell biology.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
The human mammary gland is unique in its ability to undergo proliferation, secretion and involution cycles during the course of a woman’s life. The mammary gland proliferates from a ductal structure prior to pregnancy, to a complex secretory organ during lactation when it is characterised by a high metabolic rate, equating to ~25% of the energy expenditure of the body. As the infant weans from the breast and milk production declines, involution-induced apoptosis of the mammary parenchyma leads to regression and to a ductal structure, until subsequent pregnancies once again stimulate proliferation (Neville and Daniel 1986). This ability for massive proliferation and self-renewal of the mammary gland during pregnancy suggests the presence of stem cells in the mammary tissue.
Early transplantation work demonstrated that reconstitution of mammary glands occurred when isolated mammary epithelium from donor mice were transplanted into cleared mammary fat pads (DeOme et al. 1959). More recent studies in mice have shown that small, randomly chosen fragments of mammary tissue, transfected with mouse mammary tumour virus have the capacity to repopulate a cleared mammary fat pad through the generation of clonal dominant populations as demonstrated by the unique site of viral integration (Kordon and Smith 1998). Further, following pregnancy, these glands are able to lactate and, when fragments of them are transplanted into second and third generation mammary glands, they again form functional lactating mammary glands (Kordon and Smith 1998). Studies of stem cells isolated from the human mammary gland have been restricted either to the use of biopsy material or tissue isolated from breast reduction mammoplasty (Bocker et al. 1992a, b; Petersen et al. 2003; Dontu et al. 2003; Bocker et al. 2002). These studies have utilised cytokeratin (CK) markers to identify mammary stem cells during differentiation. CKs are intermediate filaments differentially expressed in mammary epithelia dependent upon the differentiation status of the cell (Berdichevsky and Taylor-Papadimitriou 1991); CK5 is indicative of mammary stem cells (Bocker et al. 1992a, b; Petersen et al. 2003; Boecker and Buerger 2003) and CK14, CK18 and CK19, of more mature mammary epithelial cells (Dontu et al. 2003; Gudjonsson et al. 2002; Sting et al. 1998, 2001). Consistent amongst these findings is that when cultured, a heterogeneous population of mammary stem and differentiated cells, as defined by the CK markers, is observed (Sting et al. 1998, 2001).
Human breastmilk is known to have a cellular component (14,000 cells/ml) comprising various types of immune and epithelial cells, with those of epithelial lineage comprising up to 90% of the total cell population (Ho et al. 1979). Many lactocytes (mammary secretory epithelial cells) are believed to occur in milk after exfoliation from the basement membrane, either because of the turnover of the secretory tissue or as a consequence of the pressures associated with the continued filling and emptying cycle associated with breastmilk synthesis and breastfeeding. We hypothesise that, since mammary stem cells have been isolated and characterised from non-lactating mammary tissue (a period of quiescence for the mammary gland), use of identical CK markers to those previously employed should identify these same cells in breastmilk during a period of continued lactocyte renewal and proliferation-lactation.
Materials and methods
Breastmilk sample collection
Twenty breastfeeding women (stage of lactation-1 to 18 months) were recruited through the Australian Breastfeeding Association to participate in the study by donating milk as needed. Breastmilk samples were typically collected in the morning from volunteers by using a Medela Symphony Electric Breastpump (Medela AG, Switzerland) and transported on ice in polypropylene vials to the laboratory. The study was approved by the Human Research Ethics Committee of The University of Western Australia and each mother supplied written informed consent.
Cell isolation and culture
Cells were isolated from breastmilk through centrifugation at 500g for 10 min and washed three times in RPMI (Gibco). Breastmilk cells were grown in RPMI supplemented with 20% heat-inactivated fetal bovine serum (FBS; Gibco), 2 mM L-glutamine, 5 μg/ml insulin (Sigma-Aldrich), 0.5 μg/ml hydrocortisone (Sigma-Aldrich), 10 ng/ml epidermal growth factor (BD Bioscience), 60 ng/ml cholera toxin (List Biological Laboratories), penicillin (200 U/ml), streptomycin (200 μg/ml) and fungizone (0.5 μg/ml; Biowhittaker) in 24-well tissue-culture plates (Sarstedt) with 13-mm-diameter circular coverslips at 37°C under an atmosphere of 5% CO2 and 95% relative humidity until confluence. Because of the variability between women in the total number of cells in breastmilk, the time taken to confluence ranged between 7 and 50 days.
Reverse transcription/polymerase chain reaction
All materials for reverse transcription/polymerase chain reaction (RT-PCR) were purchased from Fisher Biotech unless otherwise stated. RNA from the cultured cells were extracted using Tri-reagent (Invitrogen) and reverse-transcribed with oligo-dT primers (Proligo) for 25°C for 15 min, 42°C for 60 min and 72°C for 15 min. Negative controls, without reverse transcriptase, were performed for each RNA sample to ensure the absence of DNA contamination. For PCR amplification, the conditions used were 94°C for 2 min, 30–40 cycles of 94°C for 15 s, 60°C for 15 s and 72°C for 60 s and a final extension at 72°C for 4 min. All PCR products were sequenced to confirm the correct amplification of the target product.
The primers used for PCR amplification were as follows:
-
D-glyceraldehyde-3-phosphate dehydrogenase: 5′AAGGACTCATGACCACAGTCCATG-3′ and 5′TTGATGGTACATGACAAGGTGCGG-3′;
-
nestin: 5′-GGTCAGTTCCTGAAGTTCACTCAG-3′ and 5′-CCTAGTACTATCGGGATTCAGCTG-3′;
-
CK5: 5′-CGACAAGGTGCGGTTCCTG-3′ and 5′GCAGATTGGCGCACTG-3′;
-
CK14: 5′-GATGACTTCCGCACCAAGTATGAG-3′ and 5′-TCAATCTCCAGGTTCTGCATGGTG-3′;
-
CK18: 5′-AGAAATCTGAAGGCCAGCTTGGAG-3′ and 5′-TACCCTGCTTCTGCTGGCTTAATG-3′;
Immunofluorescent labelling
Cells isolated from breast milk were grown on glass coverslips in 24-well plates and fixed with methanol/acetone (1:1 v/v). Non-specific binding was blocked with 5% rabbit serum (Jackson ImmunoResearch) and 3% bovine serum albumin (Sigma-Aldrich) in TRIS-buffered saline for 30 min at room temperature. Cells were then incubated with primary antibody (1:100) overnight at 4°C and fluorescently labelled secondary antibody (1:400) for 30 min at room temperature in the dark. The slides were then washed three times with phosphate-buffered saline (PBS) and rinsed with deionised water. Primary antibodies used were: mouse anti-CK5 (Chemicon), rabbit anti-CK14 (Chemicon), mouse anti-CK18 (Chemicon), mouse anti-CK19 (Dakocytomation) and rabbit anti-nestin (Invitrogen). Secondary antibodies used were: goat anti-rabbit IgG AlexaFluor 546 (Molecular Probes) and goat anti-mouse IgG AlexaFluor 488 (Molecular Probes). Following staining, mounting media (ProLong Gold Antifade Reagent with 4,6-diamidino-2-phenylindole; Invitrogen) was added and the cells were visualised and photographed with a Zeiss Axioplan 2 fluorescent microscope (Carl Zeiss, Australia).
Hoechst 33342 staining and fluorescence-activated cell-sorting analysis
Hoechst staining was performed as previously described (Storms et al. 2000). Briefly, cells were washed from breastmilk with warmed RPMI (Gibco), resuspended in culture medium with Hoechst 33342 (Sigma) at a final concentration of 2.5 μg/ml and incubated for 1 h at 37°C. In control samples, Verapamil (Sigma) was added (final concentration: 50 μg/ml) and, after incubation, samples were washed (BioWhittaker) and resuspended in HBSS (Gibco) supplemented with 5% FBS. For fluorescence-activated cell-sorting (FACS) anaylsis, cells were stained with 1 μg/ml propidium iodide (Sigma) for viability and then analyzed and sorted by flow cytometry by using FACStarPLUS (Becton Dickinson). The side-population and non-side-population were then cytospun onto coverslips, fixed and immunofluorescently labelled for nestin by using rabbit anti-nestin (Invitrogen; 1:100) overnight at 4°C as the primary antibody and goat anti-rabbit IgG AlexaFluor 546 (Molecular Probes; 1:400) for 30 min at room temperature in the dark as the secondary antibody. The slides were then washed three times with PBS and rinsed with deionised water.
Results and discussion
Breastmilk contains a heterogeneous population of mammary epithelial and putative mammary stem cells
To confirm that primary cultures of cells isolated from human breastmilk were heterogeneous (in that there were undifferentiated and differentiated populations present) and were thus consistent with prior studies of digested mammary tissue (Bocker et al. 1992a, b; Petersen et al. 2003; Boecker and Buerger 2003; Dontu et al. 2003; Gudjonsson et al. 2002; Sting et al. 1998, 2001), we performed immunofluorescent labelling and RT-PCR analysis on such cultures. Cells were isolated from expressed human breastmilk by centrifugation, the cell pellet was resuspended and the cells were cultured. At confluence, the cultures were then stained for the mature mammary epithelial cell markers, CK14, CK18 and CK19, and for the mammary stem cell marker, CK5.
From dual staining of CK14 and CK18, we noted two morphologically mature mammary epithelial cells. Morphologically, CK14+ cells were often elongated and large, whereas CK18+ cells were rounder and large (Fig. 1a,b). This correlated with the morphological description of the two mammary epithelial cell types of luminal/ductal and myoepithelial lineage as previously reported in cultures derived from mammary tissue (Gudjonsson et al. 2002; Sting et al. 1998, 2001; Pechoux et al. 1999). We also observed cells expressing only CK5 (Fig. 1c,d) or CK19 (Fig. 1e,f). Although occasional co-staining of CK19 and CK14 was observed, co-staining of the mature epithelial marker CK14 with the stem cell marker, CK5, was not observed, thereby contradicting reports of co-expression between these markers (Boecker and Buerger 2003). Together, these data demonstrated that the cultures derived from expressed human breastmilk were similar to those established from mammary tissue in that a heterogeneous population of mature cells and putative mammary stem cells existed.

Expression of cytokeratin (CK) markers (a–f) and light micrographs (g, h) of cells obtained from human breastmilk and subsequently cultured. Immunofluorescent labelling staining for CK14 (red in a–f) and CK18 (green in a, b), CK5 (green in c, d) or CK19 (green in e, f). Bars 50 μm
Because of the large amount of cellular debris and lipids present in expressed human breastmilk (Brooker 1980), immunofluorescent labelling of the cells isolated directly from breastmilk was problematic. Therefore, RT-PCR for the markers CK5, CK14 and CK18 was performed to determine whether this population of cells also existed in breastmilk. We observed that CK5 expression was either non-existent or below the detection threshold of RT-PCR, whereas CK18 was readily detected (Fig. 2). The lack of detection of CK5 in breastmilk suggested that CK5+ putative mammary stem cells arose in primary culture from a small population of cells in breastmilk.

Expression of CK5, CK14, CK18, nestin and D-glyceraldehyde-3-phosphate dehydrogenase (GAPDH) in cells isolated from human breastmilk (Milk) and subsequently cultured (Culture); detection by RT-PCR
Breastmilk contains a side-population of cells that are nestin-positive
The detection of the terminally differentiated markers CK14 and CK18, in combination with a lack of detection of the mammary specific stem cell marker CK5 in breastmilk, despite its presence in culture, led us to consider the possibility that CK5+ putative mammary stem cells in culture may derive from a progenitor cell in breastmilk. Nestin is a well-characterised marker of multipotent stem cells and, in adult tissues, is closely associated with stem cell populations (Lendahl et al. 1990; Dahlstrand et al. 1992; Zulewski et al. 2001; Toma et al. 2001; Kabos et al. 2002). To this end, the expression of nestin in breastmilk and our primary cultures was concomitantly investigated to determine whether the CK5+ cells observed could have originated from a multipotent progenitor present in breastmilk. Using immunofluorescent labelling, we observed cells in culture that stained exclusively positive for nestin, CK5, CK14, CK18 or CK19 (Fig. 3). Further, although heterogeneous cultures staining for nestin, CK14, CK18 and CK19 were common, the only regular patterns of co-staining observed was for nestin and CK5. As no CK5+ cells could be detected in breastmilk by using RT-PCR (Fig. 2), the presence of nestin+/CK5−, nestin+/CK5+ and nestin−/CK5+ cell types suggested that either the level of CK5 expression in breastmilk was below detection threshold or that the nestin+ cells found in breastmilk had differentiate in culture into CK5+ cells.

Co-staining of the multipotent marker, nestin, with CKs in cells obtained from human breastmilk and subsequently cultured. Immunofluorescent labelling staining for nestin (red in a–h) and CK5 (green in a, b), CK14 (green in c, d), CK18 (green in e, f) or CK19 (green in g, h). Bars 50 μm
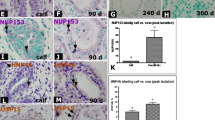
However, it was necessary to demonstrate that those nestin+ cells observed in culture and detected in breastmilk were comparable with the putative stem cells observed in other models. To this end, we investigated another characteristic of such cells, viz. the increase in membrane transporter activity as demonstrated by Hoechst 33342 dye exclusion (Zhou et al. 2001), a technique used widely to identify primitive subsets of stem/progenitor cells from a variety of tissues, including the mammary gland (Dontu et al. 2003; Alvi et al. 2003; Clarke et al. 2005; Welm et al. 2002). In cells isolated directly from breastmilk, we observed a side-population that excluded Hoechst 33342 dye (Fig. 4a) and that was not evident in control experiments in which verapamil was used to block the membrane transporters (Fig. 4b). Further, when the cells in the side-population where stained for nestin by using immunofluorescent labelling, cells positive for nestin were observed (Fig. 4c), whereas none of the cells of the non-side-population stained positively for nestin (Fig. 4d).

Hoechst 33342 exclusion and nestin expression of cells isolated directly from human breastmilk. A population excluding Hoechst 33342 (side-population) could be identified in the cells isolated from human breastmilk (gated R3), whereas the majority of cells (ungated) stained intensely with Hoechst (a). As a control, verapamil was used to block the exclusion of the Hoechst dye, resulting in the loss of the side-population (b). When the side-population and non-side-population population observed in a were fluorescently stained for nestin, most of the side-population cells, although small in number, were observed to be nestin-positive (green in c), whereas all of the non-side-population cells were nestin-negative (d). Bars 50 μm
Nestin is a marker for multipotent stem cells in bone marrow (Kabos et al. 2002) and in neural (Lendahl et al. 1990; Dahlstrand et al. 1992), pancreatic (Zulewski et al. 2001) and epithelial (Toma et al. 2001) tissues. Based on the results presented in this study, we hypothesise that human breastmilk is a source of nestin+ putative mammary stem cells. As previous studies have shown that nestin+ adult stem cells can differentiate into hepatic and exocrine phenotypes from pancreatic islet cells (Zulewski et al. 2001), into neuronal, glia, keratinocytes, melanocytes and smooth muscle cells from hair follicle stem cells (Amoh et al. 2005) and into neural cells from bone marrow stem cells (Kabos et al. 2002), the nestin+ cells in human breastmilk might have the same differentiation potential. Further, recent findings in mice in which a single transplanted mammary stem cell can reconstitute a functional mammary gland in vivo (Shackleton et al. 2006) suggest a potential for the nestin+ stem cells that we have identified in breastmilk to be utilised in the regeneration of human mammary glands.
Concluding remarks
The data described here show that expressed human breastmilk is a novel source of nestin+ putative mammary stem cells. These can be obtained non-invasively from lactating women, thereby creating a plentiful and ethical source of putative mammary stem cells for use in a wide-range of applications with regard to mammary gland and stem cell biology.
References
Alvi AJ, Clayton H, Joshi C, Enver T, Ashworth A, Vivanco MM, Dale TC, Smalley MJ (2003) Functional and molecular characterisation of mammary side population cells. Breast Cancer Res 5:R1–R8
Amoh Y, Li L, Katsuoka K, Penman S, Hoffman RM (2005) Multipotent nestin-positive, keratin-negative hair-follicle bulge stem cells can form neurons. Proc Natl Acad Sci USA 102:5530–5534
Berdichevsky F, Taylor-Papadimitriou J (1991) Morphological differentiation of hybrids of human mammary epithelial cell lines is dominant and correlates with the pattern of expression of intermediate filaments. Exp Cell Res 194:267–274
Bocker W, Bier B, Freytag G, Brommelkamp B, Jarasch ED, Edel G, Dockhorn-Dworniczak B, Schmid KW (1992a) An immunohistochemical study of the breast using antibodies to basal and luminal keratins, alpha-smooth muscle actin, vimentin, collagen IV and laminin. I. Normal breast and benign proliferative lesions. Virchows Arch A Pathol Anat Histopathol 421:315–322
Bocker W, Bier B, Freytag G, Brommelkamp B, Jarasch ED, Edel G, Dockhorn-Dworniczak B, Schmid KW (1992b) An immunohistochemical study of the breast using antibodies to basal and luminal keratins, alpha-smooth muscle actin, vimentin, collagen IV and laminin. II. Epitheliosis and ductal carcinoma in situ. Virchows Arch A Pathol Anat Histopathol 421:323–330
Bocker W, Moll R, Poremba C, Holland R, Van Diest PJ, Dervan P, Burger H, Wai D, Ina Diallo R, Brandt B, Herbst H, Schmidt A, Lerch MM, Buchwallow IB (2002) Common adult stem cells in the human breast give rise to glandular and myoepithelial cell lineages: a new cell biological concept. Lab Invest 8:737–746
Boecker W, Buerger H (2003) Evidence of progenitor cells of glandular and myoepithelial cell lineages in the human adult female breast epithelium: a new progenitor (adult stem) cell concept. Cell Prolif 36:73–84
Brooker BE (1980) The epithelial cells and cell fragments in human milk. Cell Tissue Res 210:321–332
Clarke RB, Spence K, Anderson E, Howell A, Okano H, Potten CS (2005) A putative human breast cell population is enriched for steroid receptor-positive cells. Dev Biol 277:443–456
Dahlstrand J, Zimmerman LB, McKay RD, Lendahl U (1992) Characterization of the human nestin gene reveals a close revolutionary relationship to neurofilaments. J Cell Sci 103:589–597
DeOme KB, Faulkin LJ, Bern HA, Blair PB (1959) Development of mammary tumours from hyperplastic alveolar nodules transplanted into gland free mammary fat pads of female C3H mice. Cancer Res 19:515–520
Dontu G, Abdallah WM, Foley JM, Jackson KW, Clarke MF, Kawamura MJ, Wicha MS (2003) In vitro propagation and transcriptional profiling of human mammary stem/progenitor cells. Genes Dev 17:1253–1270
Gudjonsson T, Villadsen R, Nielsen HL, Ronnov-Jessen L, Bissell MJ, Petersen OW (2002) Isolation, immortalization and characterization of a human breast epithelial cell line with stem cells properties. Genes Dev 16:693–706
Ho FCS, Wong RLC, Lawton JWN (1979) Human colostral and breast milk cells. Acta Paediatr Scand 68:389–396
Kabos P, Ehtesham M, Kabosova A, Black KL, Yu JS (2002) Generation of neural progenitor cells from whole adult bone marrow. Exp Neurol 178:288–293
Kordon EC, Smith GH (1998) An entire functional mammary gland may comprise the progeny from a single cell. Development 125:1921–1930
Lendahl U, Zimmerman LB, McKay RD (1990) CNS stem cells express a new class of intermediate filament protein. Cell 60:585–595
Neville MC, Daniel CW (1986) The mammary gland: development regulation and function. Plenum, New York
Pechoux C, Gudjonsson T, Ronnov-Jessen L, Bissell MJ, Petersen OW (1999) Human mammary luminal epithelial cells contain progenitors to myoepithelial cells. Dev Biol 206:88–99
Petersen OW, Gudjonsson T, Villadsen R, Bissell MJ, Ronnov-Jessen L (2003) Epithelial progenitor cells lines as models of normal breast morphogenesis and neoplasia. Cell Prolif 36:33–44
Shackleton M, Vaillant F, Simpson KJ, Stingl J, Smyth GK, Asselin-Labat ML, Wu L, Lindeman GJ, Visvader JE (2006) Generation of a functional mammary gland from a single stem cell. Nature 439:84–88
Sting L, Eaves CJ, Kuusk U (1998) Phenotypic and functional characterization in vitro of a multipotent epithelial cell present in the normal adult human breast. Differentiation 63:201–213
Sting L, Eaves CJ, Zandieh I, Emerman JT (2001) Characterization of bipotent mammary epithelial progenitor cells in normal adult human breast tissue. Breast Cancer Res Treat 67:93–109
Storms RW, Goodell MA, Fisher A, Mulligan RD, Smith C (2000) Hoechst dye efflux reveals a novel CD71/CD342 lymphoid progenitor in human umbilical cord blood. Blood 96:2125–2133
Toma JG, Akhavan M, Fernandes KJ, Barnabe-Heider F, Sadikot A, Kaplan DR, Miller FD (2001) Isolation of multipotent adult stem cells from the dermis of mammalian skin. Nat Cell Biol 3:778–784
Welm BE, Tepera SB, Venezia T, Graubert TA, Rosen JM, Goodell MA (2002) Sca-1 (pos) cells in the mouse mammary gland represent an enriched progenitor cell population. Dev Biol 245:42–56
Zhou S, Schuetz JD, Bunting KD, Colapietro AM, Sampath J, Morris JJ, Lagutina I, Grosveld GC, Osawa M, Nakauchi H, Sorrentino BP (2001) The ABC transporter Bcrp1/ABCG2 is expressed in a wide variety of stem cells and is a molecular determinant of the side-population phenotype. Nat Med 7:1028–1034
Zulewski H, Abraham EJ, Gerlach MJ, Daniel PB, Moritz W, Muller B, Vallejo M, Thomas MK, Habener JF (2001) Multipotential nestin-positive stem cells isolated from adult pancreatic islets differentiate ex vivo into pancreatic endocrine, exocrine, and hepatic phenotypes. Diabetes 50:521–533
Acknowledgements
The authors are grateful to all the breastfeeding mothers who participated in this research and to the Australian Breastfeeding Association for assistance in their recruitment.
Author information
Authors and Affiliations
Corresponding author
Additional information
Yiping Fan and Amber Appelbee contributed equally to this work.
This project was funded by Medela, Switzerland.
Rights and permissions
About this article
Cite this article
Cregan, M.D., Fan, Y., Appelbee, A. et al. Identification of nestin-positive putative mammary stem cells in human breastmilk. Cell Tissue Res 329, 129–136 (2007). https://doi.org/10.1007/s00441-007-0390-x
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00441-007-0390-x