
Abstract
Epimorphic regeneration is the “holy grail” of regenerative medicine. Research aimed at investigating the various models of epimorphic regeneration is essential if a fundamental understanding of the factors underpinning this process are to be established. Deer antlers are the only mammalian appendages that are subject to an annual cycle of epimorphic regeneration. In our previous studies, we have reported that histogenesis of antler regeneration relies on cells resident within the pedicle periosteum (PP). The present study elaborates this finding by means of functional studies involving the deletion of PP. Four yearling and four 2-year-old stags were selected for total PP deletion or partial PP deletion experiments. Of the animals in the total PP deletion group, one showed no signs of antler regeneration throughout the antler growth season. Two showed substantial and one showed marginal delays in antler regeneration (at 34, 20 and 7 days, respectively) compared with the corresponding sham-operated sides. Histological investigation revealed that the delayed antlers were derived from regenerated PP. Unexpectedly, the regenerative capacity of the antler from the total periosteum-deleted pedicles depended on antler length at surgery. Of the four deer that had partial PP deletion, two regenerated antlers exclusively from the left-over PP on the pedicle shafts in the absence of participation from the pedicle bone proper. The combined results from the PP deletion experiments convincingly demonstrate that the cells of the PP are responsible for antler regeneration.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Regenerative medicine is a newly emerging field that holds exciting possibilities for the replacement of damaged or missing body parts in human beings. However, the realization of these possibilities requires a fundamental understanding of the mechanism underlying the various regenerative systems (Stocum 2004). All animals have the ability to regenerate structures to a certain extent. However, true epimorphic regeneration, a phenomenon of de novo development of appendages distal to the level of amputation, is mainly found in lower vertebrates, such as amphibians (Wallace 1981). As has been well documented, epimorphic regeneration in urodele amphibians (newts and salamanders) takes place through the initial formation of a cone-shaped cell mass that is derived from the dedifferentiation of cells from diverse origins at the amputation plane of an appendage stump and that is referred to as a blastema (Mescher 1996). Interestingly, Gargioli and Slack (2004) recently reported that epimorphic tail regeneration in Xenopus tadpoles (an anuran amphibian) does not involve the process of dedifferentiation/metaplasia but is instead achieved through the proliferation and differentiation of the reserve stem cells resident in the tissue. As Xenopus tail regeneration seems more akin to the normal tissue renewal processes in mammals than to those in urodeles, Gargioli and Slack (2004) predict that the processes involved in mammalian regeneration might be closer to that of anurans (reserve stem cell-based) than to the urodeles (dedifferentiation-based).
Deer antlers are the only mammalian appendages that are cast and then fully regenerate each year (Li 2003). In our previous studies, we have reported that regenerating antler buds are morphologically and histologically not comparable to the blastemas that form over the regenerative limb stumps of urodeles (Li et al. 2004, 2005b). Kierdorf et al. (2003) have suggested that the pedicle periosteum (PP) provides the main cell source for antler regeneration, as the distal PP becomes thickened prior to antler regeneration. We have also described that the growth centres for regenerating antlers are formed directly through the proliferation and differentiation of the distal PP cells (reserve stem cells) and that no dedifferentiation process is detectable by using histological and immunohistochemical techniques (Li et al. 2005a,b). Consequently, antler regeneration may be achieved through a similar pathway to that which prevails in Xenopus tail regeneration. Our results therefore support the predictions of Gargioli and Slack (2004). However, our findings in relation to antler regeneration have to date been based solely on descriptive techniques and, therefore, studies designed to investigate the functional responses of regenerating tissue in vivo are now required.
Logically, the most conclusive way to determine whether the stem cells for antler regeneration exclusively reside within PP would be (1) to delete PP from pedicle bone prior to antler regeneration in order to ascertain whether antler regeneration can still take place and (2) to transplant PP elsewhere on the deer to observe whether ectopic antlers grow. However, significant difficulties are inherent within this approach. The prevention of antler regeneration through total pedicle removal, which would certainly include the PP, has failed because of the widespread distribution of antlerogenic tissues in the vicinity of the pedicle base (Jaczewski 1955; Bubenik and Pavlansky 1956). The induction of ectopic antler formation via PP transplantation has been equally unsuccessful (our pilot trial, see below). Thus, significant technical challenges can be expected with respect to the successful undertaking of tissue deletion and transplantation experiments designed to identify the key tissue type for antler regeneration.
The aim of the present study has been to determine whether PP in a pedicle stump is the sole tissue type responsible for antler regeneration by using the following approaches. (1) We have totally deleted the PP in order to ascertain whether antler regeneration can be substantially delayed compared with that in sham-operated control or even completely inhibited. An assumption underpinning this approach is that complete regeneration of the deleted PP will take time, even if the remaining marginal periosteum around the base of a pedicle has significant proliferation potential. If the histogenesis of antler renewal relies on PP, then antler regeneration from the PP-deleted-pedicles will inevitably be delayed. Moreover, if PP deletion is conducted at an optimal time of antler growth, antler regeneration may be completely abrogated in that growing season. (2) We have partially deleted the PP, i.e. resected 20 mm PP from the distal end of each pedicle, with the aim of observing whether a PP-derived antler can be induced to grow if the pedicle bone proper is effectively excluded from participation in antler regeneration, so that antler regeneration can only take place at an “ectopic” site markedly distant from the original position (apex of a pedicle stump) for antler regeneration.
Materials and methods
Animals
Four yearling red deer stags were selected for experiment 1 (total PP deletion) and four 2-year-old red deer stags for experiment 2 (partial PP deletion). Experiment 1 was conducted in October when the yearling stags had growing antlers of variable length (Table 1). Experiment 2 was carried out in September, about a month prior to the initiation of antler regeneration in 2-year-old stags in the southern hemisphere. All these experiments were conducted under general anaesthesia and were approved by the Invermay Animal Ethics Committee. One pedicle from each animal was randomly chosen for partial or total PP deletion and the counterlateral side was used as the sham-operated control.
Surgery
Experiment 1, total PP deletion
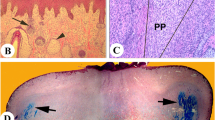
Prior to surgery the surrounding scalp skin and pedicles of each deer were thoroughly shaved and then sterilized with 70% alcohol and 1% iodine tincture (Fig. 1a). The skin around the distal end of the pedicle was incised with a scalpel 10 mm below the pedicle/antler junction and the bone was then cut with a sterile saw. The pedicle skin was retracted, a further 5 mm of bone was cut off and then a minimal amount of bone wax (Ethicon, UK) was applied to cover the central cancellous bone of the sawn surface on each pedicle stump by forcing it into the exposed bone surface and then scraping off the excess. The purpose of using bone wax was two-fold: (1) for haemostatic benefits and (2) to form a barrier so that the bone cells in the central region of the pedicle stump were blocked from participation in antler regeneration. Once applied, bone wax can persist at the bony site for years functioning as an immunologically inert foreign body (Johnson and Fromm 1981). Two vertical skin incisions were made by using a scalpel on each pedicle stump skin starting from the distal end and terminating at the boundary between the pedicle base and frontal bone. The first incision was located along the line at which the anterior and medial surfaces met, and the second was positioned along the line at which the posterior and lateral surfaces met. Pedicle skin together with major blood vessels and nerves (on the lateral side) were separated from the pedicle bone by blunt dissection and reflected outwards to expose the PP. To remove the PP from a pedicle completely, a scalpel was used to make a circumferential incision on the deer skull 5 mm away from the junction between the pedicle base and frontal bone. The PP above the circumferential incision was then cut into 5 mm wide vertical strips (Fig. 1b). Each strip was peeled off the pedicle bone by using a pair of rat-toothed forceps (Fig. 1c). The surface of the exposed pedicle bone was thoroughly scraped to ensure any cells adhering to the surface of the bone were removed. Thereafter, the reflected skin was replaced and the vertical incisions were sutured with surgical silk. A purse string suture was then applied to the skin at the distal end of the pedicle (Fig. 1d) to cover the top of the cut surface and to stop it retracting towards the pedicle base. The sham operation was conducted by using the same procedure as for the total PP deletion, except that the PP was left intact.

Total pedicle periosteum (PP) deletion on yearling red deer stags. a Preparation for surgery involved thorough shaving and sterilization of the pedicles and surrounding scalp region (pedicle height: approximately 55 mm). b PP on an artificially created pedicle stump was cut into 5 mm wide vertical strips (arrow) by using a scalpel. c PP strips were peeled off the pedicle bone with a pair of rat-toothed forceps (arrow). d A purse string was applied to the distal pedicle skin of the pedicle stump
Experiment 2, partial PP deletion
The surgical procedure was essentially the same as for Experiment 1 except that less periosteum was removed. The pedicle stumps were created by sawing off the distal end of a pedicle together with its attached hard button 10 mm beneath the pedicle/button junction (Fig. 2a). A circumferential incision was made on the PP at the site 20 mm distal to the sawn surface of each pedicle stump. Periosteal strips were peeled off with a pair of rat-toothed forceps after being divided into 5-mm wide strips by using scalpel (Fig. 2b,c). The lateral skin incision was then sutured with silk, but no purse string was applied to the distal pedicle skin (Fig. 2d).

Partial PP deletion in 2-year-old stag. a Thoroughly shaved and sterilized pedicle (pedicle height: approximately 50 mm). b PP above circumferential periosteum incision (20 mm distal to sawn plane) on an artificially created pedicle stump was cut into 5 mm wide vertical strips by using a scalpel and peeled off the pedicle bone by rat-toothed forceps (arrow). c Pedicle stump with bone lacking distal periosteum (arrow cut edge of remaining PP). d Skin incisions were sutured with silk (arrow)
Observation of stumps
For the first week after surgery, deer were monitored daily and thereafter were observed weekly and photographed fortnightly to record the date of antler regeneration and the antler length from both PP-deleted and sham-operated pedicle stumps. Observations were terminated in Experiment 1 when convincing evidence was obtained of antler regeneration from the PP deleted pedicle stumps or that growth of the regenerating antlers from the sham-operated pedicles had visibly stopped. Observations were terminated in Experiment 2 when antlers had convincingly regenerated from partial-PP-deleted pedicles.
Tissue sampling
At the termination of Experiment 1, deer were killed at a slaughterhouse. The head of each deer was recovered and carefully dissected to determine the extent to which the PP had regenerated from the marginal periosteum surrounding the base of each PP-deleted pedicle. At the same time, tissue samples were collected from both PP-deleted and sham-operated pedicles for histological examination. No tissue samples were collected from Experiment 2.
Histology
Detailed procedures for histology have been reported elsewhere (Li and Suttie 1994). Briefly, tissue samples were fixed in 10% neutral buffered formalin, decalcified, embedded in paraffin wax and sectioned at 5 μm. Haematoxylin and eosin were employed for the staining of the sections.
Results
Experiment 1, total PP deletion
Surgery was successfully undertaken on all four animals and the completion of wound healing occurred within a short time period (17–20 days). The application of the purse string suture around the distal pedicle skin successfully prevented the pedicle skin from retracting on PP-deleted pedicles. Three out of four PP-deleted pedicles regenerated antlers; however, the remaining deer showed no antler regrowth within the observed growth season. Unexpectedly, the extent of antler regeneration from the PP-deleted pedicle stumps was directly correlated with antler length at the time of surgery (Table 1). Antler regeneration from the PP-deleted pedicle stump with a 15-mm long antler at surgery was delayed by 7 days compared with the sham-operated pedicle (Fig. 3a) and antler length was 10 mm shorter at slaughter (Fig. 3b). Differences for the pedicles with 60-mm long antlers at the time of surgery widened to 20 days and 220 mm, respectively (Fig. 3c,d). These parameters for the pedicles with antlers of 150 mm increased to 34 days (Fig. 3e) and 230 mm (Fig. 3f), respectively. No antler regeneration occurred from the PP-deleted pedicle that had antlers of 175 mm in length at the time of surgery, whereas the sham-operated pedicle gave rise to an antler with three branches (Fig. 3g). The skin over the apex of the PP-deleted pedicle stump of this deer remained scalp-like throughout the experimental period (Fig. 3h).

Antler regeneration status from total-PP-deleted and sham-operated pedicles. a, b Pedicles with 15-mm long antlers at the time of PP deletion. Antler regeneration from the PP-deleted side (arrow) occurred 7 days later than on the sham-operated side (arrowhead in a shiny skin on the apex of the sham-operated pedicle) and antler length was 10 mm shorter at the end of the experiment (b). c, d Pedicles with 60-mm long antlers at the time of PP deletion. Antler regeneration from the PP-deleted side (arrow) occurred 20 days later than on the sham-operated side (arrowhead), and antler length was 220 mm shorter than that on the sham-operated side (arrowhead) at the end of the experiment (d). e, f Pedicles with 150-mm long antlers at the time of PP deletion. Antler regeneration from the PP-deleted side (arrows) occurred 34 days later than the sham-operated side (arrowhead) and antler length was 230 mm shorter at the end of the experiment (f). Note that the sham-operated pedicle formed a 3-branched antler (black arrowhead). g, h Pedicles with 175-mm antlers at the time of PP deletion. No antler regeneration occurred on the PP-deleted pedicle (arrow), although the sham-operated pedicle formed a 3-branched antler (arrowhead in g). Typical scalp skin (asterisk) remained on the apex of the PP-deleted pedicle till end of the antler growth season (h)
Dissection of these PP-deleted pedicles after slaughter clearly showed that new PP had regenerated from the cut end of the frontal periosteum in the vicinity of the pedicle base and that the regenerated PP also formed a layer of fresh trabecular bone apposed to the existing bone. The freshly formed trabecular bone tissue in the regenerating antler was continuous with that formed by the regenerated PP in the pedicle (Fig. 4a,b) indicating that the regenerating antler tissue was derived from the regenerated PP. White-coloured marks (Fig. 4a,b), reminiscent of the abscission line prior to antler casting, were clearly discernable as a narrow line up to a 1-mm wide gap. These were a result of the application of bone wax to the pedicle stump at the time of surgery. The regenerated PP at the time of slaughter was thinner and more fragile (Fig. 4c) than that from sham-operated pedicles (Fig. 4d).

Dissection and histology of total-PP deletion experiment. a, b Sagittally cut surface of PP-deleted pedicles that had given rise to antlers. Note that PP had regenerated well (arrowhead) from the remaining marginal periosteum (a) and that the regenerated PP was continuous (arrowhead) with that of the regenerating antler (at white marks in b). The white marks (arrows) were caused by the application of bone wax at surgery; this had effectively inhibited bone tissue in the central region from participating in antler regeneration. c, d PP strips (arrows) peeled off by a pair of rat-toothed froceps. Note that PP regenerated from the PP-deleted pedicles (c) was thinner and more fragile than that from the sham-operated pedicles (d). e–h PP (asterisks) with associated subperiosteal bone (SB) and partial enveloping dermis (DE). No histological difference between PP on the PP-deleted pedicles (e, f) and that on the sham-operated pedicles (g, h) was detected, either at the distal region (e, g) or at the proximal region (f, h). Note that the PP from both the PP-deleted and sham-operated pedicles at the proximal region was thinner and less active and had more mature subperiosteal bone than that at the distal region. Haematoxylin and eosin (HE) staining. Bar 0.63 mm. i Tip of an antler regenerated from a PP-deleted pedicle. Note that the regenerated antler was composed of tissue layers typical of normal antlers (distoproximally: RM reserve mesenchyme, PC precartilage, TZ transitional zone, CA cartilage). HE staining. Bar 1.6 mm
Histological examination of the sampled tissues showed that the PP-deleted pedicle stumps contained periosteum that had regenerated from the cut edge of the pedicle base/frontal bone periosteum. The regenerated PP clearly consisted of a fibrous and a cellular layer and sub-periosteal cancellous bone (Fig. 4e,f) and was similar in structure to that in the sham-operated PP (Fig. 4g,h). The tissue histology evident in the regenerated antler tips from the PP-deleted pedicles was essentially the same as that in a naturally regenerated antler (Li et al. 2002) and consisted of reserve mesenchyme, precartilage and cartilage layers (Fig. 4i).
Experiment 2, partial PP deletion
Wound healing took place 2–3 days after surgery and, by day 10, the distal pedicle skin was tightly bound to the pedicle bone. The cut edge of the pedicle dermis was sealed by the overgrowth of epidermal tissue (Fig. 5a), a phenomenon reminiscent of a pedicle carrying a hard antler. No visible difference in healing rate was detected between the sham-operated and partial-PP-deleted pedicles for each deer at this stage. Shortly thereafter, the distal pedicle skin from both sides started to retract proximally. Around 20 day post-surgery, the skin retraction ceased, leaving a distal protrusion of bare pedicle bone (Fig. 5b). Interestingly, the retraction distance of the skin on the PP-deleted pedicles (7.0–15.0 mm) was much greater than that (1.0–2.0 mm) on the sham-operated sides. Thereafter, the distal skin of each pedicle gradually became shiny and more akin to velvet and, at the same time, the exposed bony protrusions became progressively desiccated. Observation on day 33 showed that two out of four partial-PP-deleted pedicles had formed antler buds on the pedicle shafts at the site (10 and 15 mm distal to the pedicle apices, respectively) at which the cut ends of the PP and pedicle skin met (Fig. 5c). These antler buds subsequently developed into incipient antlers and the dead distal bony protrusions were ablated (on days 39 and 40, respectively) at the point at which live bone/pedicle skin and dead bone met (Fig. 5d). By day 46, 3-branched-antlers had formed (Fig. 5e). These incipient antlers formed on the shafts of the partial-PP-deleted pedicles indicating that the PP itself had successfully established interactions with pedicle skin and initiated antler regeneration. In the remaining two animals from the partial-PP-deletion trial, the distal pedicle skin became shiny and bulged out significantly around the exposed bony protrusion. The protruding distal portion of the exposed periosteum-less bone became desiccated and was eventually sequestered on days 40 and 42 post-surgery, respectively (Fig. 5f). Subsequently, these shiny and greatly bulged distal ends transformed into incipient antlers.

Antler regeneration status from partial-PP-deleted and sham-operated pedicles. a Pedicle stumps 10 days after partial PP deletion. Note that skin wounds had healed, with no visible difference in wound healing between PP-deleted (PD) and sham-operated (SO) pedicles. The dermis of the distal end of the pedicle skin was sealed by the overgrowing of its epidermis (arrow). b Pedicle stumps 20 days after surgery. Note that skin from each pedicle had retracted proximately leaving each pedicle with a protruding bone tip. The retraction distance of the pedicle skin on the PP-deleted side (PD: 7.0–15.0 mm) was greater than that (SO: 1.0–2.0 mm) on the sham-operated side. The pedicle bone lacking distal periosteum had become desiccated (asterisk). c PP-deleted pedicle 33 days after surgery. Note that an antler bud (arrow) formed on the pedicle shaft at the site at which the regenerating PP and retracting pedicle skin met, but markedly distant from the pedicle apex (asterisk). d Incipient antler (arrow) formed from the antler bud in c, but at 40 days after surgery, when the dead bony protrusion was ablated from the live and dead junction (asterisk). e Three-branched antler (arrow) 46 days after surgery, from the same antler as that in d. f Pedicle stumps 42 days after surgery. Note that the distal ends of pedicles from both sides became shiny and bulged out (black arrows), although the PP-deleted pedicle (PD) was much shorter than sham-operated pedicle (SO), as the former had lost the distal bony protrusion
Discussion
The tissue types responsible for the formation or regeneration of an organ/appendage have traditionally been defined by experiments employing the classical approach of tissue deletion. This approach (Goss 1983) together with tissue transplantation (Hartwig and Schrudde 1974) have been successfully used to discover the stem tissue (antlerogenic periosteum) for the formation of pedicles and first antlers. Experiments (Jaczewski 1955; Bubenik and Pavlansky 1956; Goss 1961) designed to pinpoint the cellular source of the annual regenerating antlers have employed similar approaches but have failed to yield clear results. These experiments have, however, convincingly demonstrated that antlerogenic potential is held in the region well beyond the pedicle base, as antler regeneration has been shown to occur after total pedicle removal. Goss (1995) has predicted that the identification of the stem cells for antler regeneration may require more sophisticated techniques than those currently available. In contrast to this current opinion, of the four total-PP-deleted pedicles in the present study, one pedicle showed no sign of antler regeneration at the end of the antler growth season and two substantially delayed their regeneration of antlers (34 and 20 days compared with the sham-operated sides). In the delayed cases, histological evidence strongly suggests that the antlers that finally grew from the total-PP deleted pedicles were derived from the regenerated PP (see Results). Of four partial-PP-deleted pedicles, two successfully regenerated ectopic antlers that were solely derived from the distal end of the left-over PP. Therefore, the results of the total and partial PP deletion experiments successfully demonstrate that PP in the pedicle stump is the sole tissue type that has the potential to initiate antler regeneration.
Total PP deletion and antler regeneration
The design of our experiment (total PP deletion) sets it apart from previous studies (total pedicle removal). In the present study, only one tissue type has been dealt with at a time, in marked contrast to the previous studies in which multiple tissue types (including bone, periosteum and skin) are involved in one deletion. Our experimental design is based on a comprehensive understanding of the histogenesis of antler regeneration obtained prior to this study. First, antler regeneration has been shown not to involve the process of dedifferentiation/metaplasia (Kierdorf et al. 2003; Li et al. 2005a); second, the growth centres for antler regeneration are histologically derived from the distal PP of a pedicle stump (Li et al. 2004, 2005b); third, pedicle skin only plays a minor role in antler regeneration as regeneration can take place in its absence (Li et al. 2006). These findings indicate that the most possible candidate tissue type for antler regeneration is the PP and hence this experiment targeted the sole tissue type, PP, rather than multiple tissue types.
We had not expected that the extent of antler regeneration from the total-PP-deleted pedicles simply depended on antler length at surgery, when we designed this experiment. One could argue that only one animal has been observed per case and that this relationship is purely accidental, resulting from a differential number of remaining PP cells on each bony shaft among these pedicles. This, however, seems unlikely as each pedicle bone surface was thoroughly scraped clean by using a scalpel after the PP has been peeled off, thus virtually all the PP cells had been removed. We believe that the relationship between antler length and regeneration potential is significant and merits further study with larger animal numbers.
The question remains as to what tissue type contributes to antler regeneration from the total-PP-deleted pedicles. There are three possibilities: the antlers are solely derived (1) from the distal end of the pedicle bone proper, (2) only from the regenerated PP, or (3) from both. The rationale for the first possibility is that, under normal conditions, the PP takes precedence of providing cells for antler regeneration and the pedicle bone contributes only a limited number of cells, if any. However, under unusual circumstances, such as in the absence of PP, pedicle bone cells can substitute for PP cells by supplying stem cells for antler regeneration; this may thus represent a backup process and highlights the crucial importance of the possession of these cranial appendages by a stag. However, careful examination of the results supports the second possibility, i.e. the antlers grow as a result of the newly regenerated PP, for two reasons. First, the PP fully regenerated in the three PP-deleted animals that had regenerated antlers at the termination of the experiment and the regenerated PP was continuous with periosteum of the regenerated antlers. Therefore, the regenerated PP participated in the process of antler regeneration. Second, bone wax applied to the sawn surface of each pedicle at surgery effectively prevented the majority of bone cells from participating in antler regeneration; this certainly favours the notion that the regenerated PP is the major source of cells for antler regeneration. If this is the case, the antler length-dependent phenomenon can be explained as follows: the antlerogenic potential of the frontal bone periosteum surrounding a pedicle base gradually decreases as antler development advances to the point at which the residual periosteal cells no longer have the ability to regenerate to the level of the distal end of a pedicle stump and thus fail to initiate antler regeneration within that antler growth season. Therefore, no antler regeneration occurred on the PP-deleted pedicle that had a 175-mm long antler. The argument against the third possibility of the involvement of both the PP and pedicle bone is that, if the pedicle bone had been a significant source of stem cells for antler regeneration, all of the total-PP-deleted pedicles would have regenerated antlers. However, one of these pedicles not only remained antlerless, but also failed to transform the apical pedicle skin into the velvet type, despite the PP-less pedicle bone and apical skin remaining in close association for more than 90 days. Consequently, we conclude that regenerating antlers in this study are derived from the regenerated PP, although the participation of a small number of cells from pedicle bone cannot be completely excluded.
Partial PP deletion and antler regeneration
The creation of a situation enabling the cut ends of the distally regenerating PP and the proximally retracting pedicle skin to meet on the pedicle shaft at a point markedly some distance away (at least 10 mm distance) from the apex of the pedicle stump in the present study successfully excludes the involvement of the pedicle bone proper but allows an opportunity for the PP solely to participate in antler regeneration. The two ectopic antlers regenerate as a result of the contribution from the PP and associated skin. Because antler regeneration can take place in the absence of pedicle skin (Fig. 6; unpublished data), we therefore conclude that PP is the key tissue type in the pedicle stump responsible for antler regeneration.

Skin-less antler (arrow) created by inserting a piece of inert membrane between bone and skin of a pedicle stump prior to antler regeneration. Pedicle skin was thus effectively excluded from participation in antler regeneration. However, antler regeneration still occurred in the absence of pedicle skin
Here one might wonder why such a complicated approach was undertaken to determine whether PP possesses the potential of launching antler regeneration, rather than simply transplanting a piece of PP subcutaneously elsewhere on the deer body to determine whether an ectopic antler could be induced, a method successfully employed in the discovery of the initial antlerogenic periosteum (Hartwig and Schrudde 1974). Indeed, we have tried this approach in 2-year-old pedicles but the results (to be published separately) show that, unlike the initial antlerogenic periosteum, PP fails to build up a sufficient tissue mass under the ectopic skin and only forms a small lump of bony tissue, even 2 years after transplantation (Fig. 7). Whether this is because the PP loses the ability to regenerate at the ectopic site or whether a sub-critical mass of tissue was transplanted is unknown at present. Because no pedicle and antler was formed at the PP-grafted sites, the partial-PP-deletion trial was undertaken as an alternative approach to determine whether PP can regenerate antlers when pedicle bone proper is successfully excluded from participation in the process.

Histological section cut through a skin tissue block sampled from the region at which PP was autologously transplanted under facial skin and remained for more than 2 years. Note that PP only formed small bony nodules (arrows) within the fibrous tissue. No ectopic antler was induced to grow from the grafted PP. HE staining. Bar 1.33 mm
Mode of antler regeneration
Epimorphic regeneration is the “holy grail” of regenerative medicine. Recently, Tsonis (2002) has proposed that epimorphic regeneration occurs primarily via two strategies. One is for differentiated cells at the damaged site to dedifferentiate to form a blastema and then subsequently redifferentiate, which is typical of epimorphic regeneration. The second strategy is for stem cells, which can be local (tissue-specific and residing in certain adult tissues), upon damage to differentiate and then reconstitute the lost part. Stem cells may also reside in the bone marrow (not tissue-specific) and migrate to the damaged site where they might differentiate into multiple cell types, such as nerve or mesenchymal cells. Based on the present study, together with our previous publications (Li et al. 2004, 2005a,b), we conclude that antler renewal, the only example of naturally occurring mammalian epimorphic regeneration, occurs via local tissue-specific stem cells, i.e. pedicle periosteal cells.
In our previous studies, we have concluded that antler regeneration is not a blastema-based process, because no dedifferentiation process is detectable during the course of antler regeneration (Li et al. 2005a) and we have defined the blastema as a cell mass formed by the dedifferentiation of all cell lineages in the amputation plane, based on previous descriptions (Wallace 1981; Goss 1983; Stocum and Maden 1990; Mescher 1996; Tsonis 2002). However, Price et al. (2005) have argued that a blastema should be defined as a mass of undifferentiated cells that has the ability to form a replacement structure and that it is irrelevant whether these cells arise from the process of dedifferentiation. If antler regeneration is to be developed as a model for the study of mammalian epimorphic regeneration, however, we believe that it is critically important to know whether antler regeneration is achieved through a process of dedifferentiation of previously differentiated cells or through differentiation from a local reserve of stem cells.
Concluding remarks
We have both substantially delayed and completely inhibited antler regeneration by total PP deletion. The unexpected finding that antler regeneration status from the PP-deleted pedicle stumps is directly correlated with antler length at the time of surgery requires further investigation. Our results from partial PP deletion have convincingly demonstrated that, in the absence of pedicle bone proper, the PP can successfully interact with the enveloping skin and launch antler regeneration at an ectopic site markedly some distance away from the orthotopic position at which antlers normally regenerate. These data therefore support our notion that stem cells for antler regeneration reside within PP tissue in a pedicle stump.
References
Bubenik AB, Pavlansky R (1956) Von welchem Gewebe geht der eigentliche Reiz zur Geweihbildung aus? II. Mitteilung: Operative Eingriffe auf den Rosenstöcken der Rehbocke, Capreolus capreolus (L., 1758). Säugetierkundliche Mitteilungen 4:97–103
Gargioli C, Slack JM (2004) Cell lineage tracing during Xenopus tail regeneration. Development 131:2669–2679
Goss RJ (1961) Experimental investigations of morphogenesis in the growing antler. J Embryol Exp Morph 9:342–354
Goss RJ (1983) Deer antlers. Regeneration, function and evolution. Academic Press, New York
Goss RJ (1995) Future directions in antler research. Anat Rec 241:291–302
Hartwig H, Schrudde J (1974) Experimentelle Untersuchungen zur Bildung der primären Stirnauswüchse beim Reh (Capreolus capreolus L.). Z Jagdwiss 20:1–13
Jaczewski Z (1955) Regeneration of antlers in red deer, Cervus elaphus L. Bull Acad Pol Sci (II) Sci Biol 3:273–278
Johnson P, Fromm D (1981) Effects of bone wax on bacterial clearance. Surgery 89:206–209
Kierdorf U, Stoffels E, Stoffels D, Kierdorf H, Szuwart T, Clemen G (2003) Histological studies of bone formation during pedicle restoration and early antler regeneration in roe deer and fallow deer. Anat Rec 273A:741–751
Li C (2003) Development of deer antler model for biomedical research. Recent Adv Res Updat 4:256–274
Li C, Suttie JM (1994) Light microscopic studies of pedicle and early first antler development in red deer (Cervus elaphus). Anat Rec 239:198–215
Li C, Clark DE, Lord EA, Stanton JA, Suttie JM (2002) Sampling technique to discriminate the different tissue layers of growing antler tips for gene discovery. Anat Rec 268:125–130
Li C, Suttie JM, Clark DE (2004) Morphological observation of antler regeneration in red deer (Cervus elaphus). J Morphol 262:731–740
Li C, Suttie JM, Clark DE (2005a) Histological examination of antler regeneration in red deer (Cervus elaphus). Anat Rec [A] Discov Mol Cell Evol Biol 282A:163–174
Li C, Suttie JM, Clark DE (2005b) Deer antler regeneration: a system which allows the full regeneration of mammalian appendages. In: Suttie JM, Haines SR, Li C (ed) Advances in antler science and product technology. Queenstown, New Zealand, pp 1–10
Li C, Yang F, Li G, Gao X, Wei H, Deng X, Clark DE (2006) Antler regeneration: a dependent process of stem tissue primed via interaction with its enveloping skin. J Exp Zool (in press)
Mescher A (1996) The cellular basis of limb regeneration in urodeles. Int J Dev Biol 40:785–795
Price JS, Allen S, Faucheux C, Althnaian T, Mount JG (2005) Deer antlers: a zoological curiosity or the key to understanding organ regeneration in mammals? J Anat 207:603–618
Stocum DL (2004) Regenerative biology and medicine: an overview. Cell Sci 1:1–19
Stocum DL, Maden M (1990) Regenerating limbs. Methods Enzymol 190:189–201
Tsonis PA (2002) Regenerative biology: the emerging field of tissue repair and restoration. Differentiation 70:397–409
Wallace H (1981) Vertebrate limb regeneration. Wiley, Chichester
Acknowledgements
The authors are grateful to Mr. Martin and the deer crew for assistance in the handling and care of the experimental deer and to Ms. Marion Labes for preparing the histological slides.
Author information
Authors and Affiliations
Corresponding author
Additional information
The authors thank the New Zealand Foundation of Research, Science and Technology and Deer Industry New Zealand for funding their research.
Rights and permissions
About this article
Cite this article
Li, C., Mackintosh, C.G., Martin, S.K. et al. Identification of key tissue type for antler regeneration through pedicle periosteum deletion. Cell Tissue Res 328, 65–75 (2007). https://doi.org/10.1007/s00441-006-0333-y
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00441-006-0333-y