
Abstract
The significance of a special kind of VE-cadherin-based, desmoplakin- and plakoglobin-containing adhering junction, originally identified in certain endothelial cells of the mammalian lymphatic system (notably the retothelial cells of the lymph node sinus and a subtype of lining endothelial cells of peripheral lymphatic vessels), has been widely confirmed and its importance in the formation of blood and lymph vessels has been demonstrated in vivo and in vitro. We have recently extended the molecular and structural characterization of the complexus adhaerens and can now report that it represents a rare and special combination of components known from three other major types of cell junction. It comprises zonula adhaerens proteins (VE-cadherin, α- and β-catenin, protein p120ctn, and afadin), desmosomal plaque components (desmoplakin and plakoglobin), and tight-junction proteins (claudin-5 and ZO-1) and forms junctions that vary markedly in size and shape. The special character and the possible biological roles of the complexus adhaerens and its unique ensemble of molecules in angiogenesis, immunology, and oncology are discussed. The surprising finding of claudin-5 and protein ZO-1 in substructures of retothelial cell-cell bridges, i.e. structures that do not separate different tissues or cell layer compartments, suggests that such tight-junction molecules are involved in functions other than the “fence” and “barrier” roles of zonulae occludentes.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
One of the most complex, tenuous, and preparatively fragile cell types of the mammalian body is the stellate cell of the lymph node sinus (“retothelial cells”, “sinus-lining cells”). Stellate cells do not form a normal endothelial layer but are characterized by numerous thin filopodia-like processes forming a closely interwoven, labyrinthine meshwork in the luminal space, either as freely suspended processes or coating the collagenous trabecular fibers (for reviews see, e.g., Fujita et al. 1973; Raviola 1975; Weiss 1977; Wacker 1994). Essential elements for the establishment and maintenance of this meshwork of retothelial cell processes are VE-cadherin-containing cell-cell adhering junctions differing markedly in size and shape. Because of their complex morphology and molecular composition (in particular, the occurrence of desmoplakin and plakoglobin but not of other desmosomal components), these junctions have been considered as an adherens junction type in their own right, are termed “complexus adhaerens”, and are integrated, in the lymph node, into a larger structural system termed a “syndesmos” (Schmelz et al. 1990, 1994; Schmelz and Franke 1993; see also Wacker et al. 1992; Wacker 1994). Similar desmoplakin-containing complexus adhaerentes have been identified in the lining endothelium cells of a specific subtype of lymphatic capillaries of various organs (Schmelz et al. 1994; Sawa et al. 1999; Ebata et al. 2001a,b; for reviews, see also Lampugnani and Dejana 1997; Sleeman et al. 2001; Jussila and Alitalo 2002; Stacker et al. 2002; Al-Rawi et al. 2005; see, however, also Fedele et al. 2004; Wessells et al. 2004) and in certain endothelial cell cultures (Valiron et al. 1996; Kowalczyk et al. 1998). Importantly, Gallicano et al. (2001) have noted the absolute requirement of desmoplakin for the formation of the vasculature system in embryogenesis and of microvessel formation in culture (Zhou et al. 2004; see also Cattelino et al. 2003).
As the retothelial jungle of cell processes in the lymph node sinus serves a series of important functions by reducing the flow speed of the percolating cells and particles, thereby facilitating the recognition, monitoring, and further treatment of the incoming components of the lymphatic fluid (including potential metastases), we have felt the need for a comprehensive cell and molecular biological analysis of the retothelial junction system of complexus adhaerentes in the lymph node sinus and in certain other parts of the lymphatic system. Here, we present the surprising finding that the complexus adhaerens is even more complex than previously shown, as it combines molecular constituents known from all three major categories of adherens junctions, i.e., zonulae adhaerentes, desmosomes, and tight junctions.
Materials and methods
Tissues and cell cultures
Freshly removed bovine or rodent tissue samples containing lymph nodes or rich in lymphatic capillaries were snap-frozen or fixed in formaldehyde or glutaraldehyde as described (Schmelz et al. 1994; Langbein et al. 2002, 2003). Human tissue samples obtained during surgery and routine pathology were similarly frozen and fixed. Aldehyde-fixed tissue specimens were dehydrated and embedded in paraffin or Epoxy resins and processed essentially as described (Schmelz et al. 1994; Langbein et al. 2002, 2003).
For comparisons, the following monolayer cell cultures and lines were used: human colon carcinoma CaCo2, glioma U333CG, keratinocytes HaCaT, endothelial HUVEC, breast carcinoma MCF-7, liver carcinoma PLC, bovine endothelial CPAE, and murine 3T3 (for sources and details, see, for example, Peitsch et al. 2001; Straub et al. 2003).
Antibodies and immunolocalization
The primary antibodies applied included diverse monoclonal and polyclonal antibodies against various cadherins (notably N-, VE-, E-cadherin, cadherin-11, desmogleins, and desmocollins), proteins binding to adhering junction plaques and actin, and tight-junction constituents (for reagents and sources, see Table 1). The specific secondary antibodies and protocols used for immunolocalization by light and electron microscopy of cryostat-sectioned material and by confocal laser-scanning immunofluorescence microscopy were as described (e.g., Langbein et al. 2002, 2003; Straub et al. 2003; for conventional electron microscopy, see Schmelz et al. 1994; Langbein et al. 2002). For light-microscopic immunolocalization of antigens in formaldehyde-fixed and paraffin-embedded material, the “antigen retrieval” technique by microwave treatment was used essentially as described (e.g., Shi et al. 2000; Peitsch et al. 2005; Riedel et al. 2005).
The cell cultures mentioned were primarily used for controls of the reactivity and specificity of the reagents used as detailed elsewhere (Mertens et al. 1996, 2001; Peitsch et al. 1999, 2001).
Results
Retothelial junctions of lymph nodes: immunofluorescence microscopy
Previously, we have shown that the cells of the retothelial meshwork of the lymph node sinus contain desmoplakin, as do the typical small desmosomes of the follicular reticulum cells of the follicles (Fig. 1). In the present study, we have attempted to determine other components of the junctions of the sinus retothelium in direct comparison with the desmosomes that connect the cells of the follicular dendritic reticulum and that are characterized by an orthodox composition, i.e., the presence of desmocollin 2, desmoglein 2, and plakophilin 2, in addition to desmoplakin and plakoglobin (see Figs. 1a, 2a; also Schmelz et al. 1994; Schmidt et al. 1999). These follicular reticulum cells also contain another kind of small junction that shows reactions of typical puncta adhaerentia (data not shown).

a Immunofluorescence microscopy of a cryostat section through a human lymph node (T trabecula) showing the localization of desmoplakin on the retothelial cells of the sinus (S) and the dendritic reticulum cells of the follicles (F). a′ Phase-contrast image. Bars 50 μm

Immunofluorescence microscopy of cryostat sections through a human lymph node showing the localization of plakoglobin (a) and desmoplakin (b). a Plakoglobin on desmosomes connecting dendritic reticulum cells of follicles (F) and those connecting the retothelial cells in the syndesmos system of the sinus (S). C Lymph node capsule, asterisk extracapsular space. b Higher magnification survey of a near-longitudinal section of a sinus. Note the cross-sections of retothelial cells with punctate or comma-shaped immunopositive structures, and the rod-like or curvilinear immunopositive structures in oblique or near-longitudinal sections. Inset Details of the region demarcated by the rectangle in b. Bars 50 μm (a), 17 μm (b)
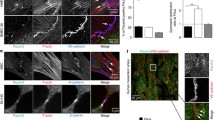
By contrast, in all three topological categories of sinus (cortical, intermediary, and medullary), the sinus-lining and retothelial cells, including their numerous processes, are connected by junctional structures that are strongly immunopositive not only for plakoglobin (Fig. 2a), but also for desmoplakin (Fig. 2b) and that vary greatly in size and appearance, from small punctate to longer whisker-like and streak-like forms (Fig. 2b; see also Schmelz and Franke 1993; Schmelz et al. 1994). This intense immunolabeling also provides a good illustration of the frequency and the ramifications of these structures, which are among the most abundant and conspicuous elements of the sinus (Figs. 2b, 3a′). Moreover, a direct comparison of the immunostaining on junctions of the follicular reticulum and the sinus retothelial cells provides an excellent control reference for the specificity and reactivity of the antibodies used. For example, among the classical desmosomal components, the significance of the negative retothelial reactions for desmoglein 2 and plakophilin 2 can directly be seen in side-by-side comparisons, with the retothelial desmoplakin reaction on the one hand and the follicular dendritic reticulum reaction on the other (Fig. 3a–b′).

Double-label immunofluorescence microscopy of cryostat sections through human lymph nodes, comparing the immunolocalization of desmoglein 2 (a) and plakophilin 2 (b) with that of desmoplakin (a′, b′). Whereas desmoglein 2 and plakophilin 2 are detected only in the small desmosomes of the dendritic reticulum cells of the follicles (F), desmoplakin is present in both the follicular dendritic reticulum (F) and the complexus adhaerentes retothelial cell system of the sinus (S). Bars 50 μm
At first glance, our negative plakophilin 2 reaction observed in the retothelium with monoclonal murine (Table 1) and polyclonal guinea pig (not shown) antibodies seems to be in contrast with the observations of Podgrabinska et al. (2002) who have detected plakophilin 2 in immunoblots of electrophoretically separated proteins of cultured lymphatic cells derived from human foreskin vessels but not in blood vessel-derived endothelial cell cultures. However, in these last-mentioned cases, plakophilin 2 is an almost ubiquitous, although minor, constitutent of the nucleus (Mertens et al. 1996, 2001). Moreover, the results might reflect the re-expression of this protein upon cell culturing (for the absence of plakophilin 2 in blood and lymph vessel endothelia in situ, see also Mertens et al. 1999). Correspondingly, the frequent colocalization of desmoplakin with VE-cadherin on retothelial cells is in direct contrast to the separate reactions of VE-cadherin on the endothelial cells of blood capillaries and of desmoplakin on follicular dendritic reticulum cells (Fig. 4a–a″,b).

Double-label immunofluorescence laser scanning microscopy of frozen tissue sections of human lymph nodes, showing the far-reaching colocalization of VE-cadherin (a) and desmoplakin (a′) in the complexus adhaerentes of the retothelial cells of a sinus (S), in contrast to the specific localization of VE-cadherin only in the endothelia of the numerous small blood vessels (V) and of desmoplakin only in the desmosomes of the dendritic reticulum cells of follicles (F) The merged image is presented in a″). The abundance and extensions of the complexus adhaerentes are illustrated with particular clarity in the merged image of the sinus shown in b. Frequent sites of colocalizations of catenins and desmoplakin in complexus adhaerentes of retothelial cells of the sinus (S) are seen in the merged image for α-catenin (c). Arrowheads denote a subset of endothelia of blood vessels (V), showing an exclusive reaction of catenins in the zonulae adhaerentes. Bars 50 μm
When we systematically examined the reactions of other adhering junction protein candidates on retothelial cells, e.g., by double immunostaining with respect to desmoplakin and plakoglobin, we noticed far-reaching but not complete colocalization with the adherens junction proteins α-catenin (Fig. 4c), β-catenin, protein p120ctn (not shown; cf. Hämmerling 2004), and afadin (Fig. 5), whereas all other cadherins and plaque components tested (compare Table 1) gave negative results. Much to our surprise, we obtained distinct and intensely positive retothelial junction reactions, showing frequent sites of colocalization with plakoglobin and desmoplakin, for claudin-5 (Fig. 6a–b′) and, to a lesser and rather variable degree, claudin-1, which were, as expected, both totally negative on other desmoplakin-positive cells, such as the dendritic reticulum cells of follicles (Fig. 6a,a′). Claudin-5 was (also as expected) also positive on the endothelia of normal blood and lymphatic vessels (e.g., Fig. 6a). We must however emphasize that the claudin-5 reaction extended over most of the retothelial junction structures, as seen, for example, by the far-reaching co-localization with desmoplakin (Fig. 6b,b′), whereas regions with disparate reactions were also frequently seen in which either claudin-5 or desmoplakin was apparent (Fig. 6). Similarly, we noted positive immunostaining for the tight-junction proteins JAM-A and ZO-1 (not shown; cf. Hämmerling 2004) on some of the complexus adhaerens structures but negative reactions for other claudins and occludin (not shown). Throughout the study, the endothelial character of the retothelial sinus and of other lymphatic structures was verified by positive reaction for the factor-VIII-related antigen and for LYVE-1, an extracellular domain of the corresponding hyaluronan receptor (not shown).

Double-label immunolocalization laser scanning microscopy of a human lymph node cryostat section showing afadin (a) and desmoplakin (a′) in an extended colocalization characteristic for the retothelial cells of a sinus (S). a″ Merged image. The adhering junctions of small blood vessels (V, arrowheads) endothelia are positive only for afadin. Bars 50 μm

Double-label immunofluorescence microscopy of frozen sections through human lymph nodes, comparing the localization of claudin-5 (a,b) and desmoplakin (a′,b′). Note the extensive colocalization in complexus adhaerentes of retothelial cells of a sinus (S), whereas an exclusive desmoplakin reaction is seen in the desmosomal structures of the follicular reticulum (F in a′). Claudin-5 is exclusively present in the adhering junctions of blood vessels (V, arrowhead). Note also that the intensity of the retothelial claudin-5 immunostaining reaction only partly corresponds to that of desmoplakin. Bars 50 μm
As we have taken special care to control our negative results, e.g., by directly comparing the reactions of the complexus adhaerens with those of other nearby structures (i.e., desmosomal components in the retothelial versus the dendritic reticulum follicular cells or in a comparison with typical puncta adhaerentia components), we feel entitled to emphasize the experimental significance of most, if not all, of our negative conclusions.
Retothelial junctions: electron microscopy
Electron microscopy confirmed and extended our previous conclusion (Schmelz and Franke 1993) that complexus adhaerens junctions markedly vary in size, as their plaque-coated regions could be traced from about 0.3 μm to more than 5 μm and often showed the typical interruptions of the plaque structure (Fig. 7a–c). Bundles of actin microfilaments and of intermediate-size filaments were seen to anchor at or laterally associate with these plaques (see, for example, Fig. 7a). However, we also noted that, in cells connected by a complexus adhaerens junction, a cell process frequently projected into an invagination of the neighboring cell and that such local filopodia-like processes also formed extended local interdigitations (e.g., Fig. 7b and, in particular, Fig. 7c). Sometimes, more than two cells were interconnected in the same region. Again, the complexus adhaerens nature of these endothelium-derived cell processes was generally verified by immunoelectron microscopy, with positive reactions for desmoplakin (Fig. 7d) and VE-cadherin (not shown).

Electron microscopy of ultrathin sections showing extended complexus adhaerens structures connecting retothelial cells of bovine lymph node sinus (T collagen-fibril-containing trabeculae, M mitochondrion). Note the typical extended plaque-coated region with small interruptions (a), a partially folded cell junction region combining several plaque-bearing regions (b, brackets), a locally intricate interdigitation complex (c; asterisks various cytoplasmic folds, arrow an example of the narrowness that such a cytoplasmic fold can attain), and the immunogold labelling of a complexus adhaerens plaque by desmoplakin antibodies (d). Bars 0.5 μm (a–c) and 0.25 μm (d)
Desmoplakin-positive junctions in endothelia of other lymphatic vessels
The unusual immunostaining reactions described above were not exclusive to lymph node sinus but were also noted in endothelia of certain low-caliber lymphatic vessels, including capillaries in various tissues. Some examples of such reactions in lymphatic vessels of tongue mucosa are presented in Fig. 8a–d″, which shows the positive immunocolocalization of lymphatic endothelial desmoplakin with α-catenin, VE-cadherin, protein ZO-1, and claudin-5 (positive reactions for β-catenin, plakoglobin, protein p120ctn, afadin, and protein JAM-A are not shown; see Hämmerling 2004). Electron microscopy revealed that these desmoplakin-positive reactions were generally associated with parts of the zonula adhaerens-like junctions encompassing these endothelial cells (not shown; for comparison see, for example, Franke et al. 1988).

Double-label immunofluorescence microscopy of cryostat sections through bovine tongue submucosa showing colocalization of α-catenin (a), VE-cadherin (b), protein ZO-1 (c) and claudin-5 (d) with desmoplakin (a′–d′) in adhering junctions of special lymphatic vessels (some are denoted by arrows; a″–d″ present the specific phase-contrast images). Note also the occurrence of α-catenin as well as desmoplakin in the specific puncta adhaerentia (a) and desmosomes (a′), respectively, of the lingual epithelium and the occurrence of other vessels which are positive for the catenins and VE-cadherin but negative for desmoplakin. Bars 50 µm
Discussion
This report confirms and markedly extends our previous conclusion that the plaque-coated complexus adhaerens junctions of a subset of lymphatic endothelia, including certain lumen-lining endothelia and retothelial cells of the lymph node sinus, represent a special kind of intercellular junction in its own right, both by morphological and molecular criteria (Schmelz et al. 1990, 1994; Schmelz and Franke 1993). We have since broadened our molecular analysis and can now state that the complexus adhaerens is a junction that combines representatives of all three of the so-called “classical” major junction categories, as it combines typical zonula adhaerens proteins (such as a type I cadherin, i.e., VE-cadherin, together with α- and β-catenin, protein p120ctn and afadin) with prominent desmosomal plaque proteins (desmoplakin and plakoglobin), and several tight-junction proteins (such as claudin-5, JAM-A, and plaque component ZO-1). Thus, the complexus adhaerens can be regarded as a recombination hybrid of members from three normally separate junction ensembles. Moreover, it can vary greatly in size and shape and, in addition to protrusions and fenestrations (Schmelz and Franke 1993), may also form single and multiple adhering interdigitations (this study).
The cell-type-specific constitutive occurrence of VE-cadherin in adhering junctions of diverse other endothelia and its functional importance for angiogenesis has been amply shown (Lampugnani et al. 1992, 1995; Breviario et al. 1995; Caveda et al. 1996; Dejana 1996; Vittet et al. 1997; Carmeliet et al. 1999; Corada et al. 1999; Gory-Fauré et al. 1999; Hordijk et al. 1999; Vorbrodt and Dobrogowska 2004). Whereas some authors have reported a colocalization with junction-bound N-cadherin (e.g., Schulze and Firth 1993; Luo and Radice 2005), others have claimed that endothelial N-cadherin is transiently or in general diffusely spread on the surface, i.e., is not integrated into the adhering junctions or enriched in the plasma membrane of the abluminal side (Salomon et al. 1992; Lampugnani and Dejana 1997; Navarro et al. 1998; Gerhardt et al. 1999; Liebner et al. 2000a,b,c; Jaggi et al. 2002; for a recent review, see Dejana 2004). Although occasional immunolocalization experiments have given us the impression that some N-cadherin and even some cadherin-11 may also act as a minor component in certain complexus-adhaerens-containing regions, we cannot yet ascribe significance to these weak and variable reactions. Studies of vasculogenesis in gene-deficient animals or gene knock-down siRNA experiments might help to determine whether VE-cadherin and N-cadherin can partly compensate for each other. Of course, we will also have to clarify whether complexus adhaerentes contain the corresponding protocadherin 12 (VE-cadherin 2; cf. Telo’ et al. 1998; Rampon et al. 2005).
With respect to tight-junction proteins, claudin-5 is widely regarded as an established general endothelial marker (Morita et al. 1999; for variations of demonstrability in certain vessels, see also Morita et al. 2003). As we have shown here, this also holds for lymphatic vessel regions characterized by complexus adhaerentes. Clearly, however, the studies of claudin-5-deficient mice by Nitta et al. (2003) have revealed that endothelial tight junctions often (perhaps generally) also claudin-12, which apparently is sufficient to maintain the tight-junction structure and most of the barrier functions in endothelia (for some minor functional deficiencies, see Nitta et al. 2003; for claims of the presence of further claudins, see, for example, Haselton and Heimark 1997; Hamm et al. 2004). As several authors have noted, some claudin-1 antibodies may cross-react with other claudins, including claudin-5 (e.g., Liebner et al. 2000a,c; Nico et al. 2003; Nitta et al. 2003), although the occasional reactions of such antibodies on endothelia and retothelia (Hämmerling 2004) have not been considered significant (see, however, for further claudins in certain endothelial tight junctions, Morcos et al. 2001; González-Mariscal et al. 2003; Wolburg et al. 2003). On the other hand, occludin has not been identified in complexus-adhaerens-containing lymphatic cells (for the absence of occludin in endothelia of certain small vessels, see also Morita et al. 2003). Localization reactions for the transmembrane proteins of the JAM family of cell-surface molecules have been found in some vessels but not in others, and so it cannot as yet be taken as a general retothelial complexus adhaerens marker (cf. Martin-Padura et al. 1998; Liu et al. 2000; Ebnet et al. 2003; for recent reviews on tight junctions in endothelial cells, see also Martin and Jiang 2001; González-Mariscal et al. 2003; Al-Rawi et al. 2005; Ji 2005).
On its cytoplasmic side, the complexus adhaerens plaque contains not only an abundance of plakoglobin and desmoplakin (Schmelz and Franke 1993; Schmelz et al. 1994; Sawa et al. 1999; Ebata et al. 2001a,b; see also Franke et al. 1987, 1988; Valiron et al. 1996; Kowalczyk et al. 1998), but also members of the actin-microfilament-anchoring protein ensemble, such as the two catenins, protein p120ctn, afadin, vinculin, and α-actinin (for blood vessel endothelia, see also Dejana 1996; Lampugnani and Dejana 1997; Navarro et al. 1998; Hamm et al. 2004), and tight-junction proteins, such as ZO-1 (e.g., Stevenson et al. 1986; Li and Poznansky 1997; Tsukita et al. 1999; Wolburg and Lippoldt 2002; Nico et al. 2003; Hamm et al. 2004; Vorbrodt and Dobrogowska 2004). At present, we are also examining the molecular composition of the intercellular junctions of the sinus-lining endothelia of the spleen; these are often characterized by prominent plaques (e.g., Heusermann and Stutte 1974; Kaiserling et al. 1989; Uehara and Miyoshi 1997; see also Schmelz et al. 1994).
This list of complexus adhaerens plaque components is probably not complete, and the specific molecular interactions in this plaque system will have to be elucidated by special techniques (for interdependences between the cadherin-catenin and the afadin-containing systems, see also Takai and Nakanishi 2003; Dejana 2004). What is obvious, however, is the essential importance of some of these plaque components for lymph and blood vessel angiogenesis, as has been demonstrated for desmoplakin with striking clarity in vivo and in vitro (Gallicano et al. 2001; Zhou et al. 2004). The demonstration of the importance of desmoplakin in embryonic vessel formation also indicates the fundamental role of the complexus adhaerens in development and suggests that the absence of desmoplakin (and thus the complexus adhaerens) in most endothelia of later stages, including adult blood vessels, reflects a secondary loss of this junction type, a conclusion that might also present an explanation for its “re-appearance” upon cell culture in vitro (e.g., Valiron et al. 1996). Moreover, the specific regulatory molecules and mechanisms involved in processes of cell recognition, signaling, and transendothelial permeability of lymphatic endothelial cells will have to be determined (for relevant data for blood endothelia and cell cultures derived therefrom, see, for example, Navarro et al. 1995; Del Maschio et al. 1996; Allport et al. 1997; Alexander et al. 1998; Kevil et al. 1998; Hordijk et al. 1999; Ratcliffe et al. 1999; Burns et al. 2000; Wong et al. 2000; Corada et al. 1999; Ferber et al. 2002; Oliver and Detmar 2002; Dejana 2004; Iyer et al. 2004).
Whether the complexus adhaerens is a relatively homogeneous molecular continuum or whether it is composed of small mosaic elements with different molecular characteristics remains uncertain. Whereas the major markers, such as desmoplakin, α-catenin, and the three arm-repeat proteins (α-catenin, plakoglobin, protein p120ctn), often show far-reaching and, in some places, complete coincidence (indicated by the yellow-orange merged color in double-label immunofluorescence microscopy), local small plaque interruptions have been seen. This extended colocalization provides a particular problem for the tight-junction markers applied. As we have not been able to resolve typical focal tight-junction membrane-membrane contacts (“kisses”), and in view of the lack of relevant electron microscopic studies in the literature, we cannot exclude that the tight-junction elements only occupy a certain proportion of the total complexus adhaerens. This is also indicated by the numerous local differences between claudin-5 and desmoplakin immunostaining (Fig. 6). Biochemical studies of the complexus adhaerens, e.g., by selective detergent solubilization, followed by immunoprecipitation or size fractionation techniques, directly or after chemical cross-linking, should help to elucidate the relative neighbor relationships of the complexus adhaerens molecules.
Our finding of typical tight-junction components, such as claudin-5 and protein ZO-1, at the complexus adhaerentes of the intercellular bridges formed by retothelial processes is surprising. Clearly, in these positions, such proteins cannot possibly serve classical tight-junction functions of continuous zonulae occludentes, i.e., the “barrier” and “fence” roles in epithelial and endothelial layers. Therefore, it will be important to examine the lymphatic system of mice deficient in the claudin-5 gene (Nitta et al. 2003) and in genes of other retothelial components will be important for determining possible, as yet overlooked, functions in this system.
Some of our findings should provide valuable cell-type markers in developmental biology and tumor pathology, in cell engineering, and in regenerative medicine. Although this has previously been demonstrated in the case of desmoplakin immunohistochemistry (e.g., Schmelz et al. 1994; Ebata et al. 2001a; see also Gallicano et al. 2001), specific systematic studies will be needed to examine and establish the value of complexus adhaerens components for the identification and characterization of certain subtypes of lymphatic endothelia, be it in normal sinus or vessels or in pathologically altered states (for an anthology of changes in endothelial structures in diseases, see Simionescu and Simionescu 1988). An extended study of the pathogenic changes of retothelial and endothelial organization in the lymph system will be published elsewhere.
References
Alexander JS, Jackson SA, Chaney E, Kevil CG, Haselton FR (1998) The role of cadherin endocytosis in endothelial barrier regulation: involvement of protein kinase C and actin-cadherin interactions. Inflammation 22:419–433
Allport JR, Ding H, Collins T, Gerritsen ME, Luscinskas FW (1997) Endothelial-dependent mechanisms regulate leukocyte transmigration: a process involving the proteasome and disruption of the vascular endothelial-cadherin complex at endothelial cell-to-cell junctions. J Exp Med 186:517–527
Al-Rawi MAA, Mansel RE, Jiang WG (2005) Lymphangiogenesis and its role in cancer. Histol Histopathol 20:283–298
Breviario F, Caveda L, Corada M, Martin-Padura I, Navarro P, Golay J, Introna M, Gulino D, Lampugnani MG, Dejana E (1995) Functional properties of human vascular endothelial cadherin (7B4/cadherin-5), an endothelium-specific cadherin. Arterioscler Thromb Vasc Biol 15:1229–1239
Burns AR, Bowden RA, MacDonell SD, Walker DC, Odebunmi TO, Donnachie EM, Simon SI, Entman ML, Smith CW (2000) Analysis of tight junctions during neutrophil transendothelial migration. J Cell Sci 113:45–57
Carmeliet P, Lampugnani MG, Moons L, Breviario F, Compernolle V, Bono F, Balconi G, Spagnuolo R, Oostuyse B, Dewerchin M, Zanetti A, Angellilo A, Mattot V, Nuyens D, Lutgens E, Clotman F, Ruiter MC de, Gittenberger-de Groot A, Poelmann R, Lupu F, Herbert JM, Collen D, Dejana E (1999) Targeted deficiency or cytosolic truncation of the VE-cadherin gene in mice impairs VEGF-mediated endothelial survival and angiogenesis. Cell 98:147–157
Cattelino A, Liebner S, Gallini R, Zanetti A, Balconi G, Corsi A, Bianco P, Wolburg H, Moore R, Oreda B, Kemler R, Dejana E (2003) The conditional inactivation of the β-catenin gene in endothelial cells causes a defective vascular pattern and increased vascular fragility. J Cell Biol 162:1111–1122
Caveda L, Martin-Padura I, Navarro P, Breviario F, Corada M, Giulino D, Lampugnani MG, Dejana E (1996) Inhibition of cultured cell growth by vascular endothelial cadherin (cadherin-5/VE-cadherin). J Clin Invest 98:886–893
Corada M, Mariotti M, Thurston G, Smith K, Kunkel R, Brockhaus M, Lampugnani MG, Martin-Padura I, Stoppacciaro A, Ruco L, McDonald DM, Ward PA, Dejana E (1999) Vascular endothelial-cadherin is an important determinant of microvascular integrity in vivo. Proc Natl Acad Sci USA 96:9815–9820
Corada M, Liao F, Lindgren M, Lampugnani MG, Breviario F, Frank R, Muller WA, Hicklin DJ, Bohlen P, Dejana E (1999) Monoclonal antibodies directed to different regions of vascular endothelial cadherin extracellular domain affect adhesion and clustering of the protein and modulate endothelial permeability.Blood 97:1679–1684
Cowin P, Kapprell H-P, Franke WW (1985) The complement of desmosomal plaque proteins in different cell types. J Cell Biol 101:1442–1454
Cowin P, Kapprell H-P, Franke WW, Tamkun J, Hynes RO (1986) Plakoglobin: a protein common to different kinds of intercellular adhering junctions. Cell 46:1063–1073
Dejana E (1996) Endothelial adherens junctions: implications in the control of vascular permeability and angiogenesis. J Clin Invest 98:1949–1953
Dejana E (2004) Endothelial cell-cell junctions: happy together. Nat Rev Mol Cell Biol 5:261–270
Del Maschio A, Zanetti A, Corada M, Rival Y, Ruco L, Lampugnani MG, Dejana E (1996) Polymorphonuclear leukocyte adhesion triggers the disorganization of endothelial cell-to-cell adherens junctions. J Cell Biol 135:497–510
Ebata N, Sawa Y, Nodasaka Y, Yamaoka Y, Yoshida S, Totsuka Y (2001a) Desmoplakin as a specific marker of lymphatic vessels. Microvasc Res 61:40–48
Ebata N, Sawa Y, Nodasaka Y, Yamaoka Y, Yoshida S, Totsuka Y (2001b) Immunoelectron microscopic study of PECAM-1 expression on lymphatic endothelium of the human tongue. Tissue Cell 33:211–218
Ebnet K, Aurrand-Lions M, Kuhn A, Kiefer F, Butz S, Zander K, Meyer zu Brickwedde M-K, Suzuki A, Imhof BA, Vestweber D (2003) The junctional adhesion molecule (JAM) family members JAM-2 and JAM-3 associate with the cell polarity protein PAR-3: a possible role for JAMs in endothelial cell polarity. J Cell Sci 116:3879–3891
Fedele C, Berens D, Rautenfeld V, Pabst R (2004) Desmoplakin and plakoglobin-specific markers of lymphatic vessels in the skin? Anat Histol Embryol 33:168–171
Ferber A, Yaen C, Sarmiento E, Martinez J (2002) An octapeptide in the juxtamembrane domain of VE-cadherin is important for p120ctn binding and cell proliferation. Exp Cell Res 274:35–44
Franke WW, Kapprell H-P, Cowin P (1987) Immunolocalization of plakoglobin in endothelial junctions: identification as a special type of zonulae adhaerentes. Biol Cell 59:205–218
Franke WW, Cowin P, Grund C, Kuhn C, Kapprell H-P (1988) The endothelial junction. The plaque and its components. In: Simionescu N, Simionescu M (eds) Endothelial cell biology in health and disease. Plenum, New York, pp 147–166
Fujita T, Miyoshi M, Murakami T (1973) Scanning electron microscope observation of the dog mesenteric lymph node. Z Zellforsch Mikrosk Anat 133:147–162
Gallicano GI, Bauer C, Fuchs E (2001) Rescuing desmoplakin function in extra-embryonic ectoderm reveals the importance of this protein in embryonic heart, neuroepithelium, skin and vasculature. Development 128:929–941
Gerhardt H, Liebner S, Redies C, Wolburg H (1999) N-cadherin expression in endothelial cells during early angiogenesis in the eye and brain of the chicken: relation to blood-retina and blood-brain barrier development. Eur J Neurosci 11:1191–1201
González-Mariscal L, Betanzos A, Nava P, Jaramillo BE (2003) Tight junction proteins. Progr Biophys Mol Biol 81:1–44
Gory-Fauré S, Prandini MH, Pointu H, Roullot V, Pignot-Paintrand I, Vernet M, Huber P (1999) Role of vascular endothelial cadherin in vascular morphogenesis. Development 126:2093–2102
Hamm S, Dehouck B, Kraus J, Wolburg-Buchholz K, Wolburg H, Risau W, Cecchelli R, Engelhardt B, Dehouck M-P (2004) Astrocyte mediated modulation of blood-brain barrier permeability does not correlate with a loss of tight junction proteins from the cellular contacts. Cell Tissue Res 315:157–166
Hämmerling B (2004) Molekulare Charakterisierung der verschiedenen Zell-Zell-Verbindungsstrukturen (“Junctions”) in Lymphknoten höherer Säugetiere [Molecular characterization of the diverse cell-cell junctions in lymph nodes of higher mammals]. MD Thesis. Faculty of Medicine, University of Heidelberg, Germany
Haselton FR, Heimark RL (1997) Role of cadherins 5 and 13 in the aortic endothelial barrier. J Cell Physiol 171:243–251
Heid HW, Schmidt A, Zimbelmann R, Schäfer S, Winter-Simanowski S, Stumpp S, Keith M, Figge U, Schnölzer M, Franke WW (1994) Cell type-specific desmosomal plaque proteins of the plakoglobin family: plakophilin 1 (band 6 protein). Differentiation 58:113–131
Heusermann U, Stutte HJ (1974) Intercellular junctions of sinus lining cells in the human spleen. Cell Tissue Res 151:337–342
Hordijk PL, Anthony E, Mul FPJ, Rientsma R, Oomen LCJM, Roos D (1999) Vascular-endothelial-cadherin modulates endothelial monolayer permeability. J Cell Sci 112:1915–1923
Iyer S, Ferreri DM, DeCocco NC, Minnear FL, Vincent PA (2004) VE-cadherin-p120 interaction is required for maintenance of endothelial barrier function. Am J Physiol Lung Cell Mol Physiol 286:L1143–L1153
Jaggi M, Wheelock MJ, Johnson KR (2002) Differential displacement of classical cadherins by VE-cadherin. Cell Commun Adhes 9:103–115
Ji RC (2005) Characteristics of lymphatic endothelial cells in physiological and pathological conditions. Histol Histopathol 20:155–175
Jussila L, Alitalo K (2002) Vascular growth factors and lymphangiogenesis. Physiol Rev 82:673–700
Kaiserling E, Wolburg H, Banks P (1989) Freeze-fracture investigation of the red pulp of human spleen. Virchows Arch B Cell Pathol 58:15–25
Kevil CG, Payne DK, Mire E, Alexander JS (1998) Vascular permeability factor/vascular endothelial cell growth factor-mediated permeability occurs through disorganization of endothelial junctional proteins. J Biol Chem 273:15099–15103
Kowalczyk AP, Navarro P, Dejana E, Bornslaeger EA, Green KJ, Kopp DS, Borgwardt JE (1998) VE-cadherin and desmoplakin are assembled into dermal microvascular endothelial intercellular junctions: a pivotal role for plakoglobin in the recruitment of desmoplakin to intercellular junctions. J Cell Sci 111:3045–3057
Kurzen H, Moll I, Moll R, Schϋfer S, Simics E, Amagai M, Wheelock MJ, Franke WW (1988) Compositionally different desmosomes in the various compartments of the human hair follicle. Differentiation 63:295–304
Lampugnani MG, Dejana E (1997) Interendothelial junctions: structure, signalling and functional roles. Curr Opin Cell Biol 9:674–682
Lampugnani MG, Resnati M, Raiteri M, Pigott R, Pisacane A, Houen G, Ruco LP, Dejana E (1992) A novel endothelial-specific membrane protein is a marker of cell-cell contacts. J Cell Biol 118:1511–1522
Lampugnani MG, Corada M, Caveda L, Breviario F, Ayalon O, Geiger B, Dejana E (1995) The molecular organization of endothelial cell to cell junctions, differential association of plakoglobin, β-catenin with vascular endothelial cadherin (VE-cadherin). J Cell Biol 129:203–217
Langbein L, Grund C, Kuhn C, Praetzel S, Kartenbeck J, Brandner JM, Moll I, Franke WW (2002) Tight junctions and compositionally related junctional structures in mammalian stratified epithelia and cell cultures derived therefrom. Eur J Cell Biol 81:419–435
Langbein L, Pape U-F, Grund C, Kuhn C, Praetzel S, Moll I, Moll R, Franke WW (2003) Tight junction-related structures in the absence of a lumen: occludin, claudins and tight junction plaque proteins in densely packed cell formations of stratified epithelia and squamous cell carcinomas. Eur J Cell Biol 82:385–400
Li C, Poznansky MJ (1997) Characterization of the ZO-1 protein in endothelial and other cell lines. J Cell Sci 97:231–237
Liebner C, Fischmann A, Rascher G, Duffner F, Grote E-H, Kalbacher H, Wolburg H (2000a) Claudin-1 and claudin-5 expression and tight junction morphology are altered in blood vessels of human glioblastome multiforme. Acta Neuropathol 100:323–331
Liebner S, Gerhardt H, Wolburg H (2000b) Differential expression of endothelial β-catenin and plakoglobin during development and maturation of the blood-brain and blood-retina barrier in the chicken. Dev Dyn 217:86–98
Liebner S, Kniesel U, Kalbacher H, Wolburg H (2000c) Correlation of tight junction morphology with the expression of tight junction proteins in blood-brain barrier endothelial cells. Eur J Cell Biol 79:707–717
Liu Y, Nusrat A, Schnell FJ, Reaves TA, Walsh S, Pochet M, Parkos CA (2000) Human junction adhesion molecule regulates tight junction resealing in epithelia. J Cell Sci 113:2363–2374
Luo Y, Radice GL (2005) N-cadherin acts upstream of VE-cadherin in controlling vascular morphogenesis. J Cell Biol 169:29–34
Martin TA, Jiang WG (2001) Tight junctions and their role in cancer metastasis. Histol Histopathol 16:1183–1195
Martin-Padura I, Lostaglio S, Schneemann M, Williams L, Romano M, Fruscella P, Panzeri C, Stoppacciaro A, Ruco L, Villa A, Simmons D, Dejana E (1998) Junctional adhesion molecule, a novel member of the immunoglobulin superfamily that distributes at intercellular junctions and modulates monocyte transmigration. J Cell Biol 142:117–127
Mertens C, Kuhn C, Franke WW (1996) Plakophilins 2a and 2b: constitutive proteins of dual location in the karyoplasm and the desmosomal plaque. J Cell Biol 135:1009–1025
Mertens C, Kuhn C, Moll R, Schwetlick I, Franke WW (1999) Desmosomal plakophilin 2 as a differentiation marker in normal and malignant tissues. Differentiation 64:277–290
Mertens C, Hofmann I, Wang Z, Teichmann M, Sepehri Chong S, Schnölzer M, Franke WW (2001) Nuclear particles containing RNA polymerase III complexes associated with the junctional plaque protein plakophilin 2. Proc Natl Acad Sci USA 98:7795–7800
Morcos Y, Hosie MJ, Bauer HC, Chan-Ling T (2001) Immunolocalization of occludin and claudin-1 to tight junctions in intact CNS vessels of mammalian retina. J Neurocytol 30:107–123
Morita K, Sasaki H, Furuse M, Tsukita S (1999) Endothelial claudin: claudin-5/TMVCF constitutes tight junction strands in endothelial cells. J Cell Biol 145:185–194
Morita K, Sasaki H, Furuse K, Furuse M, Tsukita S, Miyachi Y (2003) Expression of claudin-5 in dermal vascular endothelia. Exp Dermatol 12:289–295
Navarro P, Caveda L, Breviario F, Mândoteanu I, Lampugnani M-G, Dejana E (1995) Catenin-dependent and -independent functions of vascular endothelial cadherin. J Biol Chem 270:30965–30972
Navarro P, Ruco L, Dejana E (1998) Differential localization of VE- and N-cadherins in human endothelial cells: VE-cadherin competes with N-cadherin for junctional localization. J Cell Biol 140:1475–1484
Nico B, Frigeri A, Nicchia GP, Corsi P, Ribatti D, Quondamatteo F, Herken R, Girolamo F, Marzullo A, Svelto M, Roncali L (2003) Severe alterations of endothelial and glial cells in the blood-brain barrier of dystropic mdx mice. Glia 42:235–251
Nitta T, Hata M, Gotoh S, Seo Y, Sasaki H, Hashimoto N, Furuse M, Tsukita S (2003) Size-selective loosening of the blood-brain barrier in claudin-5-deficient mice. J Cell Biol 161:653–660
Nuber UA, Schäfer S, Stehr S, Rackwitz H-R, Franke WW (1996) Patterns of desmocollin synthesis in human epithelia: immunolocalization of desmocollins 1 and 3 in special epithelia and in cultured cells. Eur J Cell Biol 71:1–13
Oliver G, Detmar M (2002) The rediscovery of the lymphatic system: old and new insights into the development and biological function of the lymphatic vasculature. Genes Dev 16:773–783
Paffenholz R, Kuhn C, Grund C, Stehr S, Franke WW (1999) The arm-repeat protein NPRAP (neurojungin) is a constituent of the plaques of the outer limiting zone in the retina, defining a novel type of adhering junction. Exp Cell Res 250:452–464
Peitsch WK, Grund C, Kuhn C, Schnölzer M, Spring H, Schmelz M, Franke WW (1999) Drebrin is a widespread actin-associating protein enriched at junctional plaques, defining a specific microfilament anchorage system in polar epithelial cells. Eur J Cell Biol 78:767–778
Peitsch WK, Hofmann I, Prätzel S, Grund C, Kuhn C, Moll I, Langbein L, Franke WW (2001) Drebrin particles: components in the ensemble of proteins regulating actin dynamics of lamellipodia and filopodia. Eur J Cell Biol 80:567–579
Peitsch WK, Hofmann I, Bulkescher J, Hergt M, Spring H, Bleyl U, Goerdt S, Franke WW (2005) Drebrin, an actin-binding, cell type characteristic protein: induction and localization in epithelial skin tumors and cultured keratinocytes. J Invest Dermatol 125:761–774
Podgrabinska S, Braun P, Velasco P, Kloos B, Pepper MS, Jackson DG, Skobe M (2002) Molecular characterization of lymphatic endothelial cells. Proc Natl Acad Sci USA 99:16069–16074
Rampon C, Prandini M-H, Bouillot S, Pointu H, Tillet E, Franke R, Vernet M, Huber P (2005) Protocadherin 12 (VE-cadherin 2) is expressed in endothelial, trophoblast, and mesangial cells. Exp Cell Res 302:48–60
Ratcliffe MJ, Smales C, Staddon JM (1999) Dephosphorylation of the catenins p120 and p100 in endothelial cells in response to inflammatory stimuli. Biochem J 338:471–478
Raviola E (1975) Lymph nodes. In: Bloom W, Fawcett DW (eds) A textbook of histology, 10th edn. Saunders, Philadelphia, pp 471–486
Riedel I, Liang F-X, Deng F-M, Tu L, Kreibich G, Wu X-R, Sun T-T, Hergt M, Moll R (2005) Urothelial umbrella cells of human ureter are heterogeneous with respect to their uroplakin composition: different degrees of urothelial maturity in ureter and bladder. Eur J Cell Biol 84:393–405
Salomon D, Ayalon O, Patel-King R, Hynes RO, Geiger B (1992) Extrajunctional distribution of N-cadherin in cultured human endothelial cells. J Cell Sci 102:7–17
Sawa Y, Shibata K-I, Braithwaite MW, Suzuki M, Yoshida S (1999) Expression of immunoglobulin superfamily members on the lymphatic endothelium of inflamed human small intestine. Microvasc Res 57:100–106
Schmelz M, Moll R, Franke WW (1990) A new type of intercellular junction: desmosomal proteins in the extended junctions of certain endothelial cells of the lymphatic system. Cell Biol Int Rep 14:54
Schmelz M, Franke WW (1993) Complexus adhaerentes, a new group of desmoplakin-containing junctions in endothelial cells: the syndesmos connecting retothelial cells of lymph nodes. Eur J Cell Biol 61:274–289
Schmelz M, Moll R, Kuhn C, Franke WW (1994) Complexus adhaerentes, a new group of desmoplakin-containing junctions in endothelial cells. II. Different types of lymphatic vessels. Differentiation 57:97–117
Schmidt A, Langbein L, Prätzel S, Rode M, Rackwitz H-R, Franke WW (1999) Plakophilin-3-a novel cell-type specific desmosomal plaque protein. Differentiation 64:291–306
Schulze C, Firth JA (1993) Immunohistochemical localization of adherens junction components in blood-brain barrier microvessels of the rat. J Cell Sci 104:773–782
Shi S-R, Gu J, Taylor CR (2000) Antigen retrieval techniques: immunohistochemistry and molecular morphology. Eaton, Natick
Simionescu N, Simionescu M (1988) Endothelial cell biology in health and disease. Plenum, New York
Sleeman JP, Krishnan J, Kirkin V, Baumann P (2001) Markers for the lymphatic endothelium: in search of the Holy Grail? Microsc Res Tech 55:61–69
Stacker SA, Achen MG, Jussila L, Baldwin ME, Alitalo K (2002) Lymphangiogenesis and cancer metastasis. Nat Rev Cancer 2:573–583
Stevenson BR, Siliciano JD, Mooseker MS, Goodenough DA (1986) Identification of ZO-1: a high molecular weight polypeptide associated with the tight junction (zonula occludens) in a variety of epithelia. J Cell Biol 103:755–766
Straub BK, Boda J, Kuhn C, Schnoelzer M, Korf U, Kempf T, Spring H, Hatzfeld M, Franke WW (2003) A novel cell-cell junction system: the cortex adhaerens mosaic of lens fiber cells. J Cell Sci 116:4985–4995
Takai Y, Nakanishi H (2003) Nectin and afadin: novel organizers of intercellular junctions. J Cell Sci 116:17–27
Telo’ P, Breviario F, Hubert P, Panzeri C, Dejana E (1998) Identification of a novel cadherin (vascular endothelial cadherin-2) located at intercellular junctions in endothelial cells. J Biol Chem 273:17565–17572
Tsukita S, Furuse M, Itoh M (1999) Structural and signalling molecules come together at tight junctions. Curr Opin Cell Biol 11:628–633
Uehara K, Miyoshi M (1997) Junctions between the sinus endothelial cells of rat spleen. Cell Tissue Res 287:187–192
Valiron O, Chevrier V, Usson Y, Breviario F, Job D, Dejana E (1996) Desmoplakin expression and organization at human umbilical vein endothelial cell-to-cell junctions. J Cell Sci 109:2141–2149
Vittet D, Buchou T, Schweitzer A, Dejana E, Huber P (1997) Targeted null-mutation in the vascular endothelial-cadherin gene impairs the organization of vascular-like structures in embryoid bodies. Proc Natl Acad Sci USA 94:6273–6278
Vorbrodt AW, Dobrogowska DH (2004) Molecular anatomy of interendothelial junctions in human blood-brain barrier microvessels. Folia Histochem Cytobiol 42:67–75
Wacker H-H (1994) Sinuswandzellen: Immunakzessorische Zellen des Lymphknotensinus [Sinus lining cells: immune accessory cells of lymph node sinuses]. Veröffentl Pathol 143:1–217
Wacker H-H, Heidebrecht HJ, Radzun HJ, Parwaresch MR (1992) Sinuswandzellen: Morphologie, Funktion und Neoplasie [Sinus lining cells: morphology, function, and neoplasia]. Verh Dtsch Ges Pathol 76:219–225
Weiss L (1977) Lymphatic vessels and lymph nodes. In: Weiss L, Greep RO (eds) Histology. McGraw-Hill, New York, pp 523–544
Wessells H, King SH, Schmelz M, Nagle RB, Heimark RL (2004) Immunohistochemical comparison of vascular and sinusoidal adherens junctions in cavernosal endothelium. Urology 63:201–206
Wolburg H, Lippoldt A (2002) Tight junctions of the blood-brain barrier: development, composition and regulation. Vasc Pharmacol 38:323–337
Wolburg H, Wolburg-Buchholz K, Kraus J, Rascher-Eggstein G, Liebner S, Hamm S, Duffner F, Grote E-H, Risau W, Engelhardt B (2003) Localization of claudin-3 in tight junctions of the blood-brain barrier is selectively lost during experimental autoimmune encephalomyelitis and human glioblastoma multiforme. Acta Neuropathol 105:586–592
Wong EYM, Morgan L, Smales C, Lang P, Gubby SE, Staddon JM (2000) Vascular endothelial growth factor stimulates dephosphorylation of the catenins p120 and p100 in endothelial cells. Biochem J 346:209–216
Zhou X, Stuart A, Dettin LE, Rodriguez G, Hoel B, Gallicano GI (2004) Desmoplakin is required for microvascular tube formation in culture. J Cell Sci 117:3129–3140
Acknowledgements
We thank Jutta Osterholt for expert photographic data processing, Eva Gundel for careful typing of the manuscript, and Cäcilia Kuhn for skillful technical help.
Author information
Authors and Affiliations
Corresponding author
Additional information
This work was supported by a grant from the Deutsche Forschungsgemeinschaft (DFG grant MO 345/5-2).
This study is part of a thesis presented to the Faculty of Medicine of the University of Heidelberg, Germany, to fulfil the requirements of the doctoral degree (MD) of the first author.
Rights and permissions
About this article
Cite this article
Hämmerling, B., Grund, C., Boda-Heggemann, J. et al. The complexus adhaerens of mammalian lymphatic endothelia revisited: a junction even more complex than hitherto thought. Cell Tissue Res 324, 55–67 (2006). https://doi.org/10.1007/s00441-005-0090-3
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00441-005-0090-3