
Abstract
In the adventitia of large arteries, dendritic cells are located between nerve fibers, some of which contain substance P. The aim of the present study was to examine whether neurokinin 1 receptor (NK-1R) was expressed by dendritic cells in the arterial wall. Parallel sections of aortic and carotid artery segments were immunostained with anti-NK-1R and cell-type-specific antibodies. Dendritic cells in the arterial wall expressed NK-1R, albeit at a low level. Other cells, which intensely expressed NK-1R, were located along the border between the media and adventitia. They did not co-express any dendritic cell markers, including fascin, CD1a, S100, or Lag-antigen, and were negative for CD68, CD3, and mast cell tryptase. These NK-1R+ cells were laser-capture microdissected and studied by means of electron-microscopic analysis. The microdissected cells were in direct contact with nerve endings, and their ultrastructure was typical of the interstitial cells of Cajal present in the gastrointestinal tract. Further systematic electron-microscopic analysis revealed that the cells displaying the features typical of interstitial cells of Cajal were a basic element of the human arterial wall architectonics. Arterial interstitial cells of Cajal were negative for c-kit but they expressed vasoactive intestinal peptide receptor 1 (VIPR1). Destructive alterations of contacts between arterial interstitial cells of Cajal and nerve endings were observed in arterial segments with atherosclerotic lesions. The functional significance of the arterial interstitial cells of Cajal and their possible involvement in atherosclerosis and other vascular diseases need clarification.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
The abilities of dendritic cells to function as antigen-presenting cells and as co-stimulators of T cell activation are well documented (Banchereau and Steinman 1998; Lotze and Thomson 2001). Activated dendritic cells are a significant source of cytokine production (Banchereau and Steinman 1998; Lotze and Thomson 2001), and therefore, the signals responsible for dendritic cell activation in atherosclerotic arteries are of interest (Bobryshev 2000). Dendritic cells reside in the arterial wall, are involved in atherosclerosis (Bobryshev and Lord 1995, 1998; Bobryshev 2000; Millonig et al. 2001; Angeli et al. 2004; Yilmaz et al. 2004), and have been observed between nerve endings in the adventitia of large arteries (Bobryshev and Lord 1998).
Neuronal input plays an important role in the induction of immune responses with neuronally derived peptides interacting directly with cells of the immune system (Gallucci and Matzinger 2001; Steinhoff et al. 2003; O’Connor et al. 2004). Compelling evidence has implicated neuropeptide substance P (SP) in the regulation of pro-inflammatory responses by dendritic cells (Kradin et al. 1997; Marriott et al. 2000; Marriott and Bost 2001; O’Connor 2004). SP can elicit transcription factor activation in dendritic cells (Marriott et al. 2000), and SP (NK-1) receptors are present on human dendritic cells (Marriott and Bost 2001; O’Connor 2004). The arterial adventitia contains SP+ fibers (Olesen et al. 1995; Laine et al. 2000) and, therefore, the determination of whether dendritic cells express neurokinin 1 receptor (NK-1R) in the arterial wall is of interest.
The aim of the present study has been to examine whether NK-1R is expressed by dendritic cells in human large arteries. The study has identified dendritic cells expressing NK-1R in the arterial wall, albeit at a low level. Other cells, which intensely express NK-1R, have been located along the border between the media and the adventitia. They do not co-express any dendritic cell markers, including fascin, CD1a, S100, or Lag-antigen, and are negative for CD68, CD3, and mast cell tryptase. These NK-1R+ cells have been laser-capture (LC)-microdissected and studied by means of electron-microscopic analysis. The microdissected cells have been found to form direct contacts with nerve endings, and their ultrastructure is typical of interstitial cells of Cajal (ICC), which represent pacemaker cells in the gastrointestinal tract. Further systematic electron-microscopic analysis has revealed that the cells displaying the features typical of ICC are a basic element of the architectonics of human large arteries.
Analysis of the literature shows that the possibility that ICC-like cells are present in the arterial wall has been discussed for a long time. In a work on the structure and significance of the peripheral extension of the autonomic nervous system, Meyling (1953) devotes considerable description to cells that the author calls “autonomic interstitial cells”. In a later electron-microscopic study, Dahl and Nelson (1964) have noted that, in human posterior anterior cerebellar, superior cerebellar, and basilar arteries, cells containing irregular nuclei and multipolar cellular processes are rich in vesicular structures and endoplasmic reticulum and are different from surrounding smooth muscle cells. Isolated pressurized cerebral arteries are known for their ability to undergo spontaneous rhythmic contractions, which have been suggested (Lee 1995) to be generated by the multipolar cells described by Dahl and Nelson (1964). The electron-microscopic observations of Dahl and Nelson (1964) and Dahl et al. (1965) have been reviewed in 1995 by Lee and have received further attention in the studies of Pucovsky et al. (2003), Bolton et al. (2004), and Harhun et al. (2004), in which cells with physiological characteristics typical of intestinal ICC have been purified from guinea-pig mesenteric arteries and rabbit portal veins.
The present work, for the first time, provides evidence for the in situ existence of arterial cells with typical structural features of ICC. They form direct contacts with both nerve endings and smooth muscle cells at the media-adventitia border of human normal large arteries. This study has also revealed that destructive alterations of contacts between arterial ICC and nerve endings occur at the media-adventitia border in atherosclerotic segments.
Materials and methods
Tissue specimens
For this study, arterial wall segments from 12 aortas and 10 carotid arteries obtained from patients whose ages ranged from 39 to 71 years were used. The aortic specimens (entire wall segments) were collected during aortic reconstructions, and carotid specimens were obtained at endarterectomy at St Vincent’s Hospital, Sydney. Some characteristics of these specimens have been reported previously (Bobryshev and Lord 2002). The present study was carried out according to the principles outlined in the Helsinki Declaration and was approved by the institutional review board of St Vincent’s Hospital, Sydney. Informed consent was obtained from the patients in each case. Because the specimens were relatively large, atherosclerotic lesions of various types could be found, as could various sites of the arterial wall unaffected by atherosclerosis. For electron-microscopic analysis, small tissue pieces (about 1 mm3) were cut off from the different layers of the arterial wall and were fixed in 2.5% glutaraldehyde in phosphate-buffered saline (PBS). The arterial specimens were then divided into two parts. One part from each specimen was embedded in OCT compound, frozen in liquid nitrogen, and stored at −70°C until cryostat sectioning, whereas the other part of the specimen was fixed in 10% buffered formalin, processed, and embedded in paraffin. Before immunohistochemical staining, tissue sections were stained with Mayer’s hematoxylin to determine the degrees of atherosclerotic alteration of the arterial wall (Bobryshev and Lord 2002). Additionally, for further en face immunostaining, tissue sheets from four segments of macroscopically normal aortic wall were prepared as wholemount specimens according to Rekhter et al. (1991) as used previously (Bobryshev and Lord 2002).
Antibodies and immunohistochemistry
The antibodies used in the work included: anti-neurokinin 1 receptor (Novus Biologicals; NK-1R, NB3000-119; dilution, 1:1000), anti-vasoactive intestinal peptide receptor 1 (Antibody Solutions; VIPR1; AS58-P; 1:200), c-kit (Santa Cruz Biotechnology; CD117; sc-1493; 1:100), and anti-substance P (UCB; SP, i675/002; 1:200). Identification of cell types expressing NK-1R was carried out by the analysis of sets of consecutive parallel sections (6 μm thickness) that were single-immunostained with anti-NK-1R and various cell-type-specific antibodies. For dendritic cell identification, fascin (DakoCytomation, 55K-2; 1:100), CD1a (DakoCytomation; NA1/34; 1:50), S100 (DakoCytomation; S100; 1:700), and Lag-antibody (kind gift of Prof. S. Imamura, Kyoto University Hospital; 1 μg/ml) were used as previously (Bobryshev and Lord 1995; Bobryshev et al. 1997). Macrophages were identified with anti-CD68 (DakoCytomation; EBM11; 1:50). Smooth muscle cells were identified with an antibody to α-smooth muscle actin (DakoCytomation; 1A4; 1:400). Endothelial cells were identified with von Willebrand factor antibody (DakoCytomation; F8/86; 1:50). T cells were identified with anti-CD3 (DakoCytomation; CD3; 1:50). Granulocytes were identified with anti-CD15 (Dako; MMA; 1:50). Mast cells were identified with an antibody to mast cell tryptase (DakoCytomation; AA1; 1:200) or by staining with toluidine blue. Schwann cells were identified with anti-glial fibrillary acidic protein (GFAP; DakoCytomation; M0761; 1:50).
Immunohistochemical staining of serial tissue sections was carried out by using labelled streptavidin-biotin complex kit (LSAB, DakoCytomation; KO678) according to the manufacturer’s protocol, and the reaction product was visualized with Fast Red. Because the intensity of NK-1R expression in the dendritic cells (identified by the LSAB technique) was found to be specific but too weak (see below), a Dako-catalyzed signal amplification (CSA) system (DakoCytomation, K1500) in a combination with Dako CSA link (K1498) was used, with the procedures being carried out according the manufacturer’s protocol. Numbers of NK-1R + cells visualized by the LSAB technique were evaluated along the border between the media and adventitia in cross-sections of 12 non-diseased aortic segments by using a computer-associated microscope.
To examine a possible co-localization of NK-1R and SP, en-face double-immunostaining of wholemount specimens was carried out as previously described (Bobryshev and Lord 2002). For investigation of a possible co-localization of NK-1R with VIPR1, fluorescent immunohistochemistry was performed with RPE- and fluorescein isothiocyanate (FITC)-labeled secondary antibodies.
As negative controls in all single- and double-immunostaining experiments, primary antibodies were replaced with irrelevant isotope-matched antibodies.
LC-microdissection and electron microscopy of NK-1R+ cells
For LC-microdissection, arterial segments fixed in a solution containing 4% paraformaldehyde, 0.05% glutaraldehyde in PBS were cut into thin tissue slices (about 1.5 mm in thickness). Immunohistochemical reactions were carried out in the “floating” tissue slices by using the LSAB technique as above. After the immunohistochemical procedures were completed, the tissue slices were washed in PBS and re-fixed in 0.5% glutaraldehyde in PBS. After being washed in PBS, the tissue slices were oriented and embedded in OCT compound in liquid nitrogen. After being cut on a cryostat, sections were placed onto slides covered with polyethylene-naphtalate (PEN) membranes (PALM Microlaser Technologies; 1440–1000). A PALM Laser-MicroBeam System (PALM Microlaser Technologies), which enabled the contact-free isolation of single cells, was used for microdissection. Microdissected cells were catapulted into lids of 0.5-ml reaction tubes by using the laser pressure catapulting technique of the instrument.
Preparation of microdissected cells was carried out according to a modified technique described by Grant and Jerome (2002). The entire lids of 0.5-ml reaction tubes with microdissected cells were fixed in 1% glutaraldehyde in PBS, postfixed in 1% OsO4, and embedded in Araldite. Serial ultrathin sections of the embedded lids containing microdissected cells were cut and placed on formvar-coated grids. The ultrathin sections were stained with uranyl acetate and lead citrate and were examined with the aid of an electron microscope. After affirmation that the microdissected cells displayed ultrastructural characteristics typical of ICC, a systematic electron-microscopic examination of tissue samples taken from the deep media, the media-adventitia border, and from the adventitia was carried out.
Results
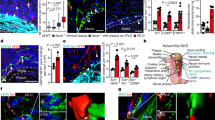
Immunohistochemical staining with the LSAB kit showed that all aortic and carotid artery specimens contained NK-1R+ cells, but the intensity of NK-1R immunopositivity varied markedly between arterial cells. In aortic specimens, cells that intensely expressed NK-1R were located along the media-adventitia border, some being present in the outermost portion of the tunica media (Fig. 1). In carotid artery specimens, which did not contain the adventitia, cells intensely expressing NK-1R were observed in the deepest portion of the media. A comparison of parallel consecutive sections immunostained with anti-NK-R1 and antibodies against dendritic cell markers showed that cells that were strongly immunopositive for NK-1R did not express any of the dendritic cell markers, including fascin, CD1a, S100, and Lag-antigen. This comparison also suggested that dendritic cells expressed NK-1R, even though the expression was low. The Dako CSA system visualized NK-1R expression in vascular dendritic cells and revealed that dendritic cells in the adventitia enriched by SP+ nerve endings also expressed NK-1R. Some macrophages (CD68+), T cells (CD3+), mast cells (mast cell tryptase+), and endothelial cells of the vasa vasorum (von Willebrand factor+) displayed NK-1R immunopositivity when the CSA system was used.

NR-1R+ cells located along the media-adventitia border in an aortic specimen. Immunohistochemistry. Bar 15 μm
The cells that intensely expressed NK-1R without signal amplification were found to be negative not only for fascin, CD1a, S100, and Lag-antigen, but also for CD68, CD3, CD15, α-smooth muscle actin, von Willebrand factor, and mast cell tryptase. The nature of these cells was of particular interest. Several NK-1R+ cells were LC-microdissected from aortic specimens (Fig. 2a, b) for ultrastructural analysis, which revealed that the microdissected cells were in intimate contacts with nerve endings (Fig. 2c–e). Nerve endings contained a large number of free and clustered “clear” vesicles of 30–50 nm in diameter and some vesicles with electron-dense cores (Fig. 2c, e). The ultrastructural architectonics of the microdissected cells was strikingly different from that of Schwann cells (Gray 1970; Ushiki et al. 1990; Fraher 2002) and from all other cell types known to reside in the aortic wall, including smooth muscle cells, fibroblasts, macrophages, lymphocytes, mast cells, pericytes, and dendritic cells, on the basis of the ultrastructural criteria for the identification of arterial cell types detailed earlier (Bobryshev and Lord 1995, 1996)). The cytoplasm and cellular processes of the microdissected cells contained a large number of caveoles and uncoated vesicles, the sizes of which varied from 30 nm to 100 nm in diameter (Fig. 2c, d). Intermediate filaments and the cisterns of granular and smooth endoplasmic reticulum were well developed, and the cells were rich in mitochondria but had few lysosomes. The nuclei were irregularly shaped with the heterochromatin being distributed mostly along the nuclear membrane. The microdissected cells did not contain the tubulovesicular system and Birbeck-granule-like structures unique to vascular dendritic cells (Bobryshev and Lord 1995, 1996). No basement membrane (basal lamina) was observed around the plasmalemma. The ultrastructural features of the microdissected cells from the aorta were typical of those of ICC (Faussone-Pellegrini and Thuneberg 1999; Vanderwinden and Rumessen 1999), the pacemaker cells of the gastrointestinal tract (Faussone-Pellegrini and Thuneberg 1999; Thuneberg 1999; Vanderwinden and Rumessen 1999; Sperelakis and Daniel 2004). LC-microdissected NK-1R+ cells from carotid arteries showed identical ultrastructural characteristics to those observed in NK-1R+ cells microdissected from the aorta. A comparative immunohistochemical analysis of parallel sections confirmed that, in aortic and carotid artery specimens, NK-R1+ cells did not express GFAP, a marker for Schwann cells.

Ultrastructural appearance of an NK-1R+ cell microdissected from the media-adventitia border of an aortic segment. a, b Consecutive stages of laser-capture (LC)-microdissection. c LC-microdissected region. d Detail of c showing the presence of a large number of vesicles and caveolae in the cytoplasm of the microdissected cell. e Detail of c showing an association of the microdissected cell (stars) with several axons (arrows). Bars 15 μm (b), 1 μm (c)
Immunostaining of NK-1R with the LSAB kit and of c-kit with the anti-c-kit antibody (CD117) in parallel sections revealed the presence of c-kit+ cells in aortic and carotid specimens, but no c-kit+ cells co-expressing NK-1R were found in any specimen. All c-kit+ cells showed typical mast cell morphology, and toluidine blue gave purple-stained granules in the cytoplasm of c-kit+ cells.
Fluorescent immunoanalysis of sections stained with the anti-NK-1R and anti-VIPR1 antibodies showed that, in all specimens, 20%–40% of NK-1R+ cells were positive for VIPR1 (Fig. 3a–c). En face immunostaining of tissue sheets prepared from the media-adventitia border demonstrated direct contacts between SP+ nerve fibers and NK-1R+ cells (Fig. 3d). The numbers of cells that intensely expressed NK-1R were quantified in tissue cross-sections along the media-adventitia border in non-diseased aortic segments and varied from 11 to 102 cells per 0.25 mm2 (mean ± SEM: 43.4±8.6; SD: 29.8).

A group of NK-1R+/VIPR1+ cells (a–c) and contacts (d) between NK-1R+ cells (red, large arrows) and SP+ nerve fibers (blue, small arrows), located at the border between the media and adventitia. a–c NK-1R+/VIPR1+ cells were identified by fluorescent IHC. d Contacts between NK-1R+ cells and SP+ nerve fibers were visualized by en face double-immunostaining in a wholemount specimen prepared from a tissue sheet of the media-adventitia junction of an aortic segment; Nomarski optics. Bars 10 μm (c, d)
The finding that ICC were present in the arterial wall led to a systematic ultrastructural analysis of the aortic wall. Tissue samples along the media-adventitia border and the underlying adventitia taken from non-diseased aortic segments were carefully examined, and ICC were identified in all specimens. In agreement with immunohistochemical data, ICC were observed to be located in the deep portion of the media with their most frequent location being along the media-adventitia junction. A few ICC were detected in the most internal portion of the adventitia, but no ICC were found in more outer portions of the adventitia. During electron-microscopic examinations, ICC were seen frequently, which suggested that only some ICC strongly expressed NK-1R.
In the adventitia, the majority of nerve fibers were unmyelinated with some nerve endings being “naked”. Nerve fibers were seen in association with Schwann cells, the ultrastructural characteristics of which were distinctive (Ushiki et al. 1990; Fraher 1996) and different from those of ICC (Fig. 4a–f). No Schwann cells were detected in the internal portion of the media-adventitia border or in the deep portion of the media, but nerve fibers and bundles of nerve fibers were present (Fig. 5a, b). ICC contacted nerve fibers in these locations (Fig. 5c–e). Reconstruction analysis of serial ultrathin sections revealed that ICC established contacts with nerve terminals through the cell processes. Synaptic-like structures with presynaptic densities and clustered vesicles were observed in these contacts (Fig. 5d, e).

Schwann cell in the aortic adventitia (a, b) and ICC located along the media-adventitia border (c, d) and in the deepest portion of the media (e, f) in non-diseased aortic segments. b Detail of a. a Several unmyelinated nerve fibers are embedded in the Schwann cell cytoplasm. b Well-developed basal (basement) membrane (arrows). d Detail of c. e Contact zone (arrow) between an ICC and a smooth muscle cell (SMC). f Complex interconnections between ICC processes. Electron microscopy. Bars 1 μm (a), 5 μm (c, e), 0.5 μm (f)

a “Naked” nerve fibers (arrows) in the deep portion of the aortic media. b Group of nerve fibers in the deep portion of the aortic media. c–e Bundle of axonal varicosities apposed to the ICC plasmalemma (c), and several axons (arrows in e) associated with an ICC process (star in e) located at the media-adventitia border of a non-diseased aortic segment. d Detail of c. Electron microscopy. Bars 0.25 μm (a, b, d, e)
Electron-microscopic examination showed that ICC were distributed irregularly and often formed clusters along the media-adventitia border. In the media, only some ICC were in direct contact with nerve fibers, but ICC formed local networks through their long cellular processes. Complex labyrinth-like contacts were observed between ICC, but no gap junctions were detected (Fig. 4f). ICC were in direct contact with smooth muscle cells in the deep portion of the media (Fig. 5e). No specialized structures, such as gap junctions, were detected in zones of ICC/smooth muscle cell contacts.
In arterial segments with advanced atherosclerotic lesions, cells strongly expressing NK-1R were seen along the media-adventitia border in which they were distributed irregularly (as found in the specimens unaffected by atherosclerosis). However, in contrast to non-atherosclerotic segments, a few NK-1R+ cells were detected in areas underlying the plaque necrotic core (Fig. 6a). All carotid plaques contained a few NK-1R+ ICC. NK-1R+ ICC were found in eight of ten aortic plaques. Electron-microscopic examination confirmed the presence of ICC amongst foam cells (Fig. 6b) and revealed that these ICC had no structural contacts with smooth muscle cells. Some ICC displayed signs of degeneration. Condensation of the mitochondria matrix and the complete destruction of mitochondria were evident in ICC with edematous cytoplasm (Fig. 6c, d). Some ICC also displayed signs of destructive alterations at the media-adventitia border in atherosclerotic segments (Fig. 6e, f). Degeneration of nerve endings contacting injured ICC was also observed (Fig. 6e, f).

NK-1R+ cells (a) and destructive alterations of arterial ICC (b–d) in atherosclerotic segments of the aorta. a IHC. b-f Electron micriscopy. b Foam cell (FC). d Detail of c. Note the destruction of mitochondria and vacuolization of the cytoplasm. f Detail of e. Note the edema of the perinuclear space in an ICC (star), the degenerating nerve ending (black arrow), and intact nerve endings (white arrows). Bars 15 μm (a), 1 μm (b), 3 μm (c, e)
Discussion
The present work reports several novel observations. First, the study has revealed that dendritic cells in the arterial wall express NK-1R, even though this expression is low and identifiable only when an amplifying kit is used. This observation is in agreement with the report of Marriott and Bost (2001) describing NK-1R expression by human dendritic cells in vitro. The detection of NK-1R+ expression in vascular dendritic cells suggests that they may be responsive to the release of SP and, thus, may serve in the arterial wall as a link between the nervous and immune systems. The study has also revealed that other cell types, including macrophages, T cells, mast cells, and endothelial cells of the vasa vasorum, express NK-1R. The effects of SP on endothelial cells, macrophages, and T cells have been examined in a number of in vitro studies (Kang et al. 2001; Annunziata et al. 2002; Guo et al. 2002). The present observations therefore indicate that these effects identified in vitro may be involved in the neuronal regulation of non-neuronal cells in the arterial wall.
The key finding of the present work is that a subset of NK-1R+ cells display a unique combination of ultrastructural features enabling these NK-1R+ cells to be identified as a subset of the ICC family. The family of ICC consists of intestinal ICC and ICC subtypes recently recognized in the urogenital tract (Klemm et al. 1999; Sergeant et al. 2000; Pezzone et al. 2003; van der AA et al. 2004; Metzger et al. 2004). ICC are thought to represent mesenchymal cells arranged in a characteristic manner in close relation to nerves and smooth muscle cells in the outer muscular coats of the gastrointestinal tract (Faussone-Pellegrini and Thuneberg 1999; Thuneberg 1999; Vanderwinden and Rumessen 1999; Sperelakis and Daniel 2004). Evidence suggests that ICC are pacemakers in specified regions of the gut in some species, including humans (Faussone-Pellegrini and Thuneberg 1999; Thuneberg 1999; Vanderwinden and Rumessen 1999; Takayama et al. 2002; Iino et al. 2004; Sperelakis and Daniel 2004). Electrophysiological studies have indicated that ICC are actively involved in nerve transmission in the gastrointestinal tract (Koh et al. 2003; Ward et al. 2004).
In the present work, ICC residing in the arterial wall have been designated as arterial ICC (AICC). Systematic electron-microscopic examination of aortic and carotid specimens have revealed that AICC represent a basic element of the architectonics of the human large arteries.
Our study suggests that AICC possess distinctive characteristics and represent a unique subtype within the ICC family. Because of the absence of c-kit expression, AICC might be more closely related to a subset of c-kit-negative ICC in the gastrointestinal tract than to the majority of intestinal ICC that express c-kit. At the present, there is no specific immunohistochemical marker(s) for the identification of AICC. c-kit (CD117) is the most commonly used marker for the identification of intestinal ICC (Komuro et al. 1996; Faussone-Pellegrini and Thuneberg 1999; Vanderwinden and Rumessen 1999). However, c-kit is not expressed by all ICC in the gastrointestinal tract (Faussone-Pellegrini and Thuneberg 1999; Vanderwinden and Rumessen 1999). Furthermore, c-kit is expressed by a variety of cell types including stem cells in the arterial wall (Hu et al. 2004). In the present study, c-kit+ cells have been found in the adventitia of the arterial wall, but all adventitial c-kit+ cells have been identified as mast cells. In the gastrointestinal tract, ICC are known to contain vimentin, desmin, and smooth muscle myosin (Faussone-Pellegrini and Thuneberg 1999; Vanderwinden and Rumessen 1999), but these proteins are expressed in various combinations by the different smooth muscle cell phenotypes (Babaev et al. 1993; Thomas and Campbell 2000; Martinez-Gonzalez et al. 2001; Hao et al. 2003). Several studies have demonstrated the expression of NK-1R by ICC in the gastrointestinal tract (Portbury et al. 1996; Lavin et al. 1998; Jun et al. 2004). According to the present study, only a proportion of ICC in the arterial wall strongly express NK-1R, suggesting that AICC may represent a heterogeneous cell population. We have shown that some arterial NK-1R+ ICC co-express VIPR1, another marker for ICC identification (Epperson et al. 2000). Further studies are needed to investigate the combinations of markers expressed by different AICC phenotypes.
In the absence of unique immunochemical marker(s), electron microscopy remains “the gold standard” for the identification of ICC (Komuro et al. 1996, 1999; Komuro 1999; Faussone-Pellegrini and Thuneberg 1999; Thuneberg 1999; Vanderwinden and Rumessen 1999). Although the ultrastructural criteria for the identification of ICC and their intercellular interactions are well established and reliable (Komuro et al. 1996, 1999; Faussone-Pellegrini and Thuneberg 1999; Vanderwinden and Rumessen 1999), the lack of electron-microscopic studies of the arterial adventitia and deep portion of the media may explain why the presence of ICC in the human arterial wall has not been previously recognized.
The arterial wall is innervated with nerve endings localized mostly in the adventitia (Olesen et al. 1995; Laine et al. 2000). Direct contacts between SP+ nerve endings and AICC along the media-adventitia border have been demonstrated in the present study. Moreover, nerve endings intrude into the deep portion of the media in which AICC seem to be interposed between autonomic axons and smooth muscle cells, a feature that also occurs in some intestinal ICC (Ward and Sanders 2001). Previous studies have suggested that disturbances of innervation in the arterial wall may affect or even initiate the development of atherosclerotic lesions (Gutterman 1999). A comparison between AICC numbers in non-diseased arterial segments and those in segments affected by atherosclerosis is of interest. Because of the high level of variation in AICC numbers along the media-adventitia border in non-diseased aortic segments, such a comparative analysis would need a selection of representative areas and, thus, would require the use of autopsy material rather than surgically discarded tissue specimens. The present study suggests that, in advanced atherosclerosis, some AICC lose their contacts with nerve fibers and smooth muscle cells, and that some AICC migrate from the media-adventitia border to the disorganized media and atherosclerotic plaques in which AICC undergo destructive alterations. If AICC function as a link between smooth muscle cells and nerve endings, the injury or loss of AICC in atherosclerotic arteries might affect arterialsmooth muscle cell functions. If AICC are shown to be altered functionally in atherosclerotic vessels, the challenge will then be to determine whether such changes are primary or secondary in the disease process.
References
Angeli V, Llodra J, Rong JX, Satoh K, Ishii S, Shimizu T, Fisher EA, Randolph GJ (2004) Dyslipidemia associated with atherosclerotic disease systemically alters dendritic cell mobilization. Immunity 21:561–574
Annunziata P, Cioni C, Santonini R, Paccagnini E (2002) Substance P antagonist blocks leakage and reduces activation of cytokine-stimulated rat brain endothelium. J Neuroimmunol 131:41–49
Babaev VR, Bobryshev YV, Stenina OV, Tararak EM, Gabbiani G (1993) Heterogeneity of smooth muscle cells in atheromatous plaque of human aorta. Am J Pathol 136:1031–1042
Banchereau J, Steinman RM (1998) Dendritic cells and the control of immunity. Nature 392:245–252
Bobryshev YV (2000) Dendritic cells and their involvement in atherosclerosis. Curr Opin Lipidol 11:511–517
Bobryshev YV, Lord RS (1995) Ultrastructural recognition of cells with dendritic cell morphology in human aortic intima. Contacting interactions of vascular dendritic cells in athero-resistant and athero-prone areas of the normal aorta. Arch Histol Cytol 58:307–322
Bobryshev YV, Lord RSA (1996) Langhans cells of human arterial intima: uniform by stellate appearance but different by nature. Tissue Cell 28:177–194
Bobryshev YV, Lord RSA (1998) Mapping of vascular dendritic cells in atherosclerotic arteries suggests their involvement in local immune-inflammatory reactions. Cardiovasc Res 37:799–810
Bobryshev YV, Lord RSA (2002) Expression of heat shock protein-70 by dendritic cells in the arterial intima and its potential significance in atherogenesis. J Vasc Surg 35:368–375
Bobryshev YV, Ikezawa T, Watanabe T (1997) Formation of Birbeck granule-like structures in vascular dendritic cells in human atherosclerotic aorta. Lag-antibody to epidermal Langerhans cells recognizes cells in the aortic wall. Atherosclerosis 133:193–202
Bolton TB, Gordienko DV, Povstyan OV, Harhun MI, Pucovsky V (2004) Smooth muscle cells and interstitial cells of blood vessels. Cell Calcium 35:643–657
Dahl E, Nelson E (1964) Electron microscopic observations on human intracranial arteries. Arch Neurol 10:158–164
Dahl E, Flora G, Nelson E (1965) Electron microscopic observations on normal human intracranial arteries. Neurology 15:132–140
Epperson A, Hatton WJ, Callaghan B, Doherty P, Walker RL, Sanders KM, Ward SM, Horowitz B (2000) Molecular markers expressed in cultured and freshly isolated interstitial cells of Cajal. Am J Physiol Cell Physiol 279:C529–C539
Faussone-Pellegrini MS, Thuneberg L (1999) Guide to the identification of interstitial cells of Cajal. Microsc Res Tech 47:248–266
Fraher J (1996) Node distribution and packing density in the rat CNS-PNS transitional zone.Microsc Res Tech 346:507–521
Fraher J (2002) Axons and glial interfaces: ultrastructural studies. J Anat 200:415–430
Gallucci S, Matzinger P (2001) Danger signals: SOS to the immune system.Curr Opin Immunol 13:114-119
Grant K, Jerome WG (2002) Laser capture microdissection as an aid to ultrastructural analysis. Microsc Microanal 8:170–175
Gray EG (1970) The fine structure of nerve. Comp Biochem Physiol 36:419–448
Guo CJ, Lai JP, Luo HM, Douglas SD, Ho WZ (2002) Substance P up-regulates macrophage inflammatory protein-1beta expression in human T lymphocytes. J Neuroimmunol 131:160–167
Gutterman DD (1999) Adventitia-dependent influences on vascular function. Am J Physiol 277:H1265-H1272
Hao H, Gabbiani G, Bochaton-Piallat ML (2003) Arterial smooth muscle cell heterogeneity: implications for atherosclerosis and restenosis development. Arterioscler Thromb Vasc Biol 23:1510–1520
Harhun MI, Gordienko DV, Povstyan OV, Moss RF, Bolton TB (2004) Function of interstitial cells of Cajal in the rabbit portal vein. Circ Res 95:619–626
Hu Y, Zhang Z, Torsney E, Afzal AR, Davison F, Metzler B, Xu Q (2004) Abundant progenitor cells in the adventitia contribute to atherosclerosis of vein grafts in ApoE-deficient mice. J Clin Invest 113:1258–1265
Iino S, Ward SM, Sanders KM (2004) Interstitial cells of Cajal are functionally innervated by excitatory motor neurones in the murine intestine. J Physiol (Lond) 556:521–530
Jun JY, Choi S, Yeum CH, Chang IY, You HJ, Park CK, Kim MY, Kong ID, Kim MJ, Lee KP, So I, Kim KW (2004) Substance P induces inward current and regulates pacemaker currents through tachykinin NK1 receptor in cultured interstitial cells of Cajal of murine small intestine. Eur J Pharmacol 495:35–42
Kang BN, Jeong KS, Park SJ, Kim SJ, Kim TH, Kim HJ, Ryu SY (2001) Regulation of apoptosis by somatostatin and substance P in peritoneal macrophages. Regul Pept 101:43–49
Klemm MF, Exintaris B, Lang RJ (1999) Identification of the cells underlying pacemaker activity in the guinea-pig upper urinary tract. J Physiol (Lond) 519:867–884
Koh SD, Ward SM, Ordog T, Sanders KM, Horowitz B (2003) Conductances responsible for slow wave generation and propagation in interstitial cells of Cajal. Curr Opin Pharmacol 3:579–582
Komuro T (1999) Comparative morphology of interstitial cells of Cajal: ultrastructural characterization. Microsc Res Tech 47:267–285
Komuro T, Tokui K, Zhou DS (1996) Identification of the interstitial cells of Cajal. Histol Histopathol 11:769–786
Komuro T, Seki K, Horiguchi K (1999) Ultrastructural characterization of the interstitial cells of Cajal. Arch Histol Cytol 62:295–316
Kradin R, MacLean J, Duckett S, Schneeberger EE, Waeber C, Pinto C (1997) Pulmonary response to inhaled antigen: neuroimmune interactions promote the recruitment of dendritic cells to the lung and the cellular immune response to inhaled antigen. Am J Pathol 150:1735–1743
Laine P, Naukkarinen A, Heikkila L, Penttila A, Kovanen PT (2000) Adventitial mast cells connect with sensory nerve fibers in atherosclerotic coronary arteries. Circulation 101:1665–1669
Lavin ST, Southwell BR, Murphy R, Jenkinson KM, Furness JB (1998) Activation of neurokinin 1 receptors on interstitial cells of Cajal of the guinea-pig small intestine by substance P. Histochem Cell Biol 110:263–271
Lee RM (1995) Morphology of cerebral arteries. Pharmacol Ther 66:149–173
Lotze MT, Thomson AW (2001) Dendritic cells: biology and clinical applications, 2nd edn. Academic Press, San Diego
Marriott I, Bost KL (2001) Expression of authentic substance P receptors in murine and human dendritic cells. J Neuroimmunol 114:131–141
Marriott I, Mason MJ, Elhofy A, Bost KL (2000) Substance P activates NF-kappaB independent of elevations in intracellular calcium in murine macrophages and dendritic cells. J Neuroimmunol 102:163–171
Martinez-Gonzalez J, Berrozpe M, Varela O, Badimon L (2001) Heterogeneity of smooth muscle cells in advanced human atherosclerotic plaques: intimal smooth muscle cells expressing a fibroblast surface protein are highly activated by platelet-released products. Eur J Clin Invest 31:939–949
Metzger R, Schuster T, Till H, Stehr M, Franke FE, Dietz HG (2004) Cajal-like cells in the human upper urinary tract. J Urol 172:769–772
Meyling HA (1953) Structure and significance of the peripheral extension of the autonomic nervous system.J Comp Neurol 99:495-543
Millonig G, Niederegger H, Rabl W, Hochleitner BW, Hoefer D, Romani N, Wick G (2001) Network of vascular-associated dendritic cells in intima of healthy young individuals. Arterioscler Thromb Vasc Biol 21:503–508
O’Connor TM, O’Connell J, O’Brien DI, Goode T, Bredin CP, Shanahan F (2004) The role of substance P in inflammatory disease. J Cell Physiol 201:167–180
Olesen IJ, Gulbenkian S, Valenca A, Antunes JL, Wharton J, Polak JM, Edvinsson L (1995) The peptidergic innervation of the human superficial temporal artery: immunohistochemistry, ultrastructure, and vasomotility. Peptides 16:275–287
Pezzone MA, Watkins SC, Alber SM, King WE, Groat WC de, Chancellor MB, Fraser MO (2003) Identification of c-kit-positive cells in the mouse ureter: the interstitial cells of Cajal of the urinary tract. Am J Physiol Renal Physiol 284:F925–F929
Portbury AL, Furness JB, Young HM, Southwell BR, Vigna SR (1996) Localisation of NK1 receptor immunoreactivity to neurons and interstitial cells of the guinea-pig gastrointestinal tract. J Comp Neurol 367:342–351
Pucovsky V, Moss RF, Bolton TB (2003) Non-contractile cells with thin processes resembling interstitial cells of Cajal found in the wall of guinea-pig mesenteric arteries. J Physiol (Lond) 552:119–133
Rekhter MD, Andreeva ER, Mironov AA, Orekhov AN (1991) Three-dimensional cytoarchitecture of normal and atherosclerotic intima of human aorta. Am J Pathol 138:569–580
Sergeant GP, Hollywood MA, McCloskey KD, Thornbury KD, McHale NG (2000) Specialised pacemaking cells in the rabbit urethra. J Physiol (Lond) 526:359–66
Sperelakis N, Daniel EE (2004) Activation of intestinal smooth muscle cells by interstitial cells of Cajal in simulation studies. Am J Physiol Gastrointest Liver Physiol 286:G234–G243
Steinhoff M, Stander S, Seeliger S, Ansel JC, Schmelz M, Luger T (2003) Modern aspects of cutaneous neurogenic inflammation. Arch Dermatol 139:1479–1488
Takayama I, Horiguchi K, Daigo Y, Mine T, Fujino MA, Ohno S (2002) The interstitial cells of Cajal and a gastroenteric pacemaker system. Arch Histol Cytol 65:1–26
Thomas AC, Campbell JH (2000) Contractile and cytoskeletal proteins of smooth muscle cells in rat, rabbit, and human arteries. Tissue Cell 32:249–256
Thuneberg L (1999) One hundred years of interstitial cells of Cajal. Microsc Res Tech 47:223–238
Ushiki T, Hayashi S, Ide C (1990) The filamentous meshwork in the Schwann cell basement membrane as revealed by transmission and scanning electron microscopy. Arch Histol Cytol 53:523–530
van der AA F, Roskams T, Blyweert W, Ost D, Bogaert G, De Ridder D (2004) Identification of kit positive cells in the human urinary tract. J Urol 171:2492–2496
Vanderwinden JM, Rumessen JJ (1999) Interstitial cells of Cajal in human gut and gastrointestinal disease. Microsc Res Tech 47:344–360
Ward SM, Sanders KM (2001) Interstitial cells of Cajal: primary targets of enteric motor innervation. Anat Rec 262:125–135
Ward SM, Sanders KM, Hirst GD (2004) Role of interstitial cells of Cajal in neural control of gastrointestinal smooth muscles. Neurogastroenterol Motil 16 (Suppl 1):112–117
Yilmaz A, Lochno M, Traeg F, Cicha I, Reiss C, Stumpf C, Raaz D, Anger T, Amann K, Probst T, Ludwig J, Daniel WG, Garlichs CD (2004) Emergence of dendritic cells in rupture-prone regions of vulnerable carotid plaques. Atherosclerosis 176:101–110
Author information
Authors and Affiliations
Corresponding author
Additional information
This work was supported by the St Vincent’s Clinic Foundation, Sydney, Australia.
Rights and permissions
About this article
Cite this article
Bobryshev, Y.V. Subset of cells immunopositive for neurokinin-1 receptor identified as arterial interstitial cells of Cajal in human large arteries. Cell Tissue Res 321, 45–55 (2005). https://doi.org/10.1007/s00441-004-1061-9
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00441-004-1061-9