
Abstract
To study the mitochondrial DNA (mtDNA) polymorphisms in a total of 232 individuals from five ethnic populations (Daur, n=45; Ewenki, n=47; Korean, n=48; Mongolian, n=48; Oroqen, n=44) in northern China, we analyzed the control region sequences and typed for a number of characteristic mutations in coding regions (especially the region 14576–16047), by direct sequencing or restriction-fragment-length-polymorphism (RFLP) analysis. With the exception of 14 individuals belonging to the European-specific haplogroups R2, H, J, and T, the mtDNAs considered could be assigned into the East Asian-specific haplogroups described recently. The polymorphisms in cytochrome b sequence were found to be very informative for defining or supporting the haplogroups status of East Asian mtDNAs in addition to the reported regions 10171–10659 and 14055–14590 in our previous study. The haplogroup distribution frequencies varied in the five ethnic populations, but in general they all harbored a large amount of north-prevalent haplogroups, such as D, G, C, and Z, and thus were in agreement with their ethnohistory of northern origin. The two populations (Ewenki and Oroqen) with small population census also show concordant features in their matrilineal genetic structures, with lower genetic diversities observed.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
China stands at a geographical crossroads between Southeast Asia, Central Asia, and Siberia, and consists of a very important region for tracing the migration and expansion of the anatomically modern human to these areas. Although a number of recent studies using different genetic markers have shed some light on the peopling in East Asia, especially in China (Ding et al. 2000; Karafet et al. 2001; Kivisild et al. 2002; Yao et al. 2000b, 2002a), debates have not been compromised. One of the major problems causing the controversy is due to the fact that the samples considered were not fully matched: populations from southern China were much more sampled than populations from northern China (c.f. Ding et al. 2000). Among the available mtDNA data of Chinese ethnic populations (Oota et al. 2002; Yao et al. 2000a, 2002a, 2002b, 2003a; Yao and Zhang 2002), relatively fewer populations from northern China were analyzed compared with those from southern and central China. To better understand the migration pattern of East Asia, more data from northern China is thus necessary.
Based on a satisfactorily resolved East Asian mtDNA phylogeny that was constructed either by combining information provided by control region and coding region (Yao et al. 2002a) or by complete sequences (Kivisild et al. 2002; Kong et al. 2003), we dissected the mtDNA lineages identified in 232 subjects in five ethnic groups sampled from northern China into different haplogroups, then compared the genetic structure of these populations on the basis of distribution frequency of each haplogroup. We followed the strategy for haplogroup classification as described in Yao et al. (2002a, 2003a) and Kivisild et al. (2002). In brief, the mtDNAs were tentatively assigned into respective haplogroups according to their specific mutations observed in the hypervariable segment I (HVS-I) and by (near-)matching with the reported data with coding region information available, then, other specific mutations in the hypervariable segment II (HVS-II) and/or coding region were typed to further characterize the haplogroup status of the mtDNAs. In addition, the region 14576–16047 [according to the revised Cambridge reference sequence (rCRS; Andrews et al. 1999)], which covering the complete cytochrome b gene (Cyt b), was sequenced in a total of 63 individuals [25 from this study and 38 from our previous studies (Yao et al. 2002a, 2002b, 2003a) as well as our unpublished data set], with an intention to learn whether this region is as informative as regions 10171–10659 and 14055–14590 in Yao et al. (2002a) for defining and/or supporting the haplogroups of East Asian mtDNAs. Our results showed that there were differences in the maternal genetic structures of the five ethnic populations considered, and the information provided by the region (14576–16047) was helpful for discerning 15 (sub-)haplogroups.
Material and methods
Sampling
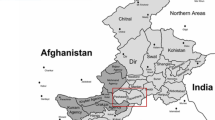
A total of 232 individuals from five ethnic populations were sampled in Inner Mongolia, China: 45 Daurs from Ewenkizu Zizhiqi county, 47 Ewenkis from Xin Barag Zuoqi county, 48 Koreans from Arunqi county, 48 Mongolians from Xin Barag Zuoqi county, and 44 Oroqens from Oroqen Zizhiqi county (Fig. 1). All of the individuals were confirmed to be unrelated before sampling and were given informed consent.

Geographic locations of the five ethnic populations in northern China
DNA amplification and sequencing
The mtDNA HVS-I sequence was amplified and sequenced in all the samples using the procedures described elsewhere (Yao et al. 2000a, 2002a, 2002b). In order to confirm the haplogroup status of some individuals, a number of characteristic mutations in HVS-II sequence and/or coding regions were further detected by direct sequencing or RFLP analysis using the same primer pairs and conditions as in Yao et al. (2002a). Moreover, all of the individuals were screened for the mtDNA 9-bp deletion in the COII/tRNALys intergenic region according to our previous studies (Yao et al. 2000b, 2001).
The region 14576–16047, which covers the whole Cyt b gene sequence and harbors two characteristic polymorphisms (14783 and 15043) of macro-haplogroup M, was amplified by using primer pair L14575 (5′-ACCCGACCACACCGCTAACA-3′)/H16048 (5′-GTCAATACTTGGGTGGTACC-3′). After being purified on spin columns (Watson BioTechnologies, Shanghai), the PCR products were directly sequenced for both strands by using the two primers for amplification and six internal primers (L14752, 5′-ACTACAAGAACACCAATGACC-3′; L14989, 5′-ATGGCTGAATCATCCGCTAC-3′; L15391, 5′-TAGGAATCACCTCCCATTCC-3′, L15598, 5′-ACACAATTCTCCGATCCGTC-3′; H15086, 5′-AGGAGGATAATGCCGATGTT-3′; H15400, 5′-TGTAGTAAGGGTGGAAGGTG-3′). The numbers in the primer names refer to the position of the 3′ end of the primer sequence relative to rCRS (Andrews et al. 1999). L and H stand for light and heavy strands, respectively.
Data analyses
The sequences were edited and aligned by DNAstar software and compared with the rCRS (Andrews et al. 1999). The length polymorphisms of the A and C stretches in region 16180–16193 (triggered by the 16189 T/C substitution) were disregarded in the analysis.
The mtDNAs were classified into the specific (sub-)haplogroups by using the strategy and haplogroup annotation system as fully described in recent studies (Richards et al. 2000; Kivisild et al. 2002; Yao et al. 2002a, 2003a; Kong et al. 2003). After each mtDNA was assigned into the most-derived named haplogroup, the haplogroup distribution frequencies in each of the five populations were then estimated. We also compared the haplogroup distribution pattern in our samples with that of the recently reported data from Siberia (Buryat and Yakut; Pakendorf et al. 2003). The haplotype diversity and nucleotide diversity (Nei 1987) in the populations were computed by using the DnaSP package (Rozas and Rozas 1999). In addition, using the information provided by the complete sequences of the Cyt b genes analyzed in this study and in the reported complete sequences (Kivisild et al. 2002; Kong et al. 2003), we reanalyzed the recently reported Cyt b data from Koreans (Lee et al. 2002) and tried to pinpoint the potential errors in their sequence data.
Nomenclature
Gene mutation nomenclature used in this article follows the recommendations of den Dunnen and Antonarakis (2001). Gene symbols used in this article follow the recommendations of the HUGO Gene Nomenclature Committee (Povey et al. 2001). The authors have made every attempt to perform the study in accordance with the recommendations made by Cooper et al. (2002).
Results
Haplogroup identification
The mtDNA sequence variation in the 232 individuals was listed in Table 1. It should be noted that all of the samples, with the exception of two individuals, could be classified into the most-derived named mtDNA haplogroups (Fig. 2). The two mtDNAs that could not be assigned further were labeled M and N, respectively. The one M mtDNA from the Korean sample (Kor92), with mutation motif 16145-16148-16188-16189-16223, might belong to a new (sub-)haplogroup of M that is still not defined. This motif can also be found in the Tibetan samples from Yunnan (Yao and Zhang 2002). The one N haplotype from the Oroqen sample (Oro16) near-matches a Han Chinese sample from Wuhan, Hubei (WH6976; Yao et al. 2002a).

Classification tree of the mtDNA haplogroups identified in 232 northern Chinese samples. This tree is constructed with reference to the classification trees of Yao et al. (2002a), Kivisild et al. (2002), and Kong et al. (2003). The characteristic mutations (relative to the revised CRS; Andrews et al. 1999) considered here are indicated on the branches with an arbitrary order. The suffix indicates a transversion, d indicates deletion, and recurrent mutations are underlined. The revised CRS branches out from the R node by seven haplogroup-specific mutations at sites 73, 1438, 2706, 4769, 7028, 11719, and 14766, plus four private mutations at sites 263, 750, 8860, and 15326
Table 2 shows the sequence polymorphisms identified in region 14576–16047 in the 63 individuals. This region provides useful information in supporting the poorly characterized haplogroups that only defined by the control region motifs (Kivisild et al. 2002; Yao et al. 2002a, 2003a). For instance, haplogroup B5b, a sub-haplogroup of B5, which was formerly defined by HVS-I transitions at sites 16140, 16189, and 16243, is newly confirmed by five specific mutations (15223, 15508, 15662, 15851, and 15927) in this region. Similarly, haplogroup B5a is identified by the 15235 mutation, haplogroup B4b is supported by a transition at site 15535, haplogroup B4c is recognized by 15346 mutation, haplogroups M8a, C, and Z share a specific transversion at site 15487. Haplogroup M10, which was formerly defined by mutations 10646 and 16311 (Yao et al. 2002a), could be well recognized by mutations at sites 15040, 15071, and 15218. The newly described haplogroup G1 (Bandelt et al. 2003; Kong et al. 2003), which is a sub-haplogroup of haplogroup G, is characteristic of mutations 15323 and 15497. Mutation 15860 and the HVS-I motif (16223-16325-16362) further defined a sub-branch of G1, G1a. The region 14576–16047 also provides information for defining several European specific haplogroups (Finnilä et al. 2001; Herrnstadt et al. 2002), such as JT (characterized by mutation 15452A), T (recognizable by mutations 14905, 15607, and 15928), and H4 (characterized by mutations 14766 and 14582). Note that a subset of J is also identifiable by a transition at site 14798 (Finnilä et al. 2001; Herrnstadt et al. 2002). As a result, 14 individuals from the Daur, Ewenki, and Mongolian samples considered here could be assigned into the west European-specific haplogroups R2, H4, J1, T1, and T2 (Table 1). Our extensive searching for the reported data in Chinese showed that haplogroups J and T also occurred in samples from Liaoning, Shaanxi, Hunan, and Xinjiang Provinces (Oota et al. 2002; Yao et al. 2000a, 2002a). The question about the invasion and spread of these western-Eurasian-specific lineages across China is still unspecific, and further analysis is needed to resolve it.
Errors in the reported Korean data
The mtDNA coding region information has been employed in forensic science recently (Tzen et al. 2001; Lee et al. 2002). The now available data with coding region and control region information (Kivisild et al. 2002; Yao et al. 2002a; Kong et al. 2003; this study) could be used as a benchmark to check the potential reading errors or artificial recombination in the reported data set. In the 98 Korean samples that were sequenced for the complete Cyt b sequence and the two hypervariable segments of control region (Lee et al. 2002), several obvious errors caused by possible sample crossover can be easily discerned: sample H84 is a crossover of M7a1 with D4a; F531.2 is a crossover of F1b and D4a; H81 is a crossover of A5 with D4a; H98 is a crossover of A5 and M. Besides these recombination errors, there are many overlooked polymorphisms: 16290 might be missed in H98; 15326 might be overlooked in H108 and F907.1; 249d might be disregarded in SB41; in the three G1a samples, F916.2 and F844.1 all lacked 15497, while F408.2 missed 15323. The transversion at site 15487, which is shared by haplogroups M8a, C, and Z, was neglected in sample F496.1. Moreover, oversight of mutation 16362 seems to be frequent for the D4a types that were identifiable via transition at site 14979. These seemingly artificial errors caused by sample crossover or other reasons are not infrequent. The 8.8-kb length of mtDNA sequences of Native Americans reported by Silva et al. (2002) also contained such problems (Yao et al. 2003b, 2003c). Even with extreme caution during the bench work, such errors may occur (c.f. Kong et al. 2003; Yao et al. 2003a). Thus, additional quality control measures, such as independent typing for the region by different individuals (Herrnstadt et al. 2002), matching or near-matching with reliable data sets, and detecting errors by phylogenetic analysis as described recently (Bandelt et al. 2001, 2002; Yao et al. 2003b, 2003d; Yao and Zhang 2003) should be extensively employed to avoid possible errors in the data.
Haplogroup distribution
Table 3 shows the haplogroup distribution frequencies in the five northern ethnic groups, as well as in the samples reported by Pakendorf et al. (2003), from which several features can be discerned: (1) haplogroups D, D5, G, and A are distributed widely among the seven populations; (2) haplogroups G2 (including G2a), M9a, Y, and the sub-haplogroups of F1 (including F1a, F1b, and F1c) have limited distributions in these samples; (3) some north-prevalent haplogroups — namely, D, G, C, and Z (Yao et al. 2002a, 2003a) — have relative high frequencies (altogether more than 45%) in these populations; (4) the frequency of haplogroup B is quite high (more than 7.1%) in Daur, Ewenki, Korean, Mongolian, and Buryat, but is lower in either Oroqen (2.3%) or Yakut (0.0%); (5) haplogroup M7 shows high frequency in Daur (20.0%) and Korean (8.4%), which, however, were either absent or with low frequency in other populations.
Discussion
The emerging mtDNA phylogeny of East Asian mtDNAs and the available data set with coding region and control region information can serve as the foundation for the East Asian mtDNA haplogroup assignment, and this has been fully described in a series of recent studies (Kivisild et al. 2002; Yao et al. 2002a, 2003a; Kong et al. 2003). The dissection of the 232 mtDNAs from northern China in this study by the same strategy revealed that: (1) most of the mtDNAs could be classified into the most-derived named mtDNA haplogroups (Fig. 2), and (2) some haplogroups, such as D, G, C, and Z, were prevalent in these northern samples and the matrilineal genetic profile was consistent with the genetic pattern observed recently (Yao et al. 2002a).
Two mtDNA coding region segments (10171–10659 and 14055–14590) analyzed in Yao et al. (2002a) were found to be very informative for East Asian mtDNA haplogroup characterization. However, these segments provided little information in supporting the status of haplogroups G1, G1a, M8, M10, B4b, B4c, B5a, and B5b. Our analyses of the region 14576–16047 showed that it contained many characteristic polymorphisms for these haplogroups and filled the lacunae. We suggested that when discerning the haplogroup status of the major haplogroups in East Asian mtDNAs based on short coding-region segments, these three segments should be the ideal choices.
The comparison of the matrilineal genetic structure of the ethnic populations could reflect their ethnohistory more or less (Yao et al. 2002a, 2002b; Yao and Zhang 2002). The Koreans in China are mainly the descendents of migrants from the Korean Peninsula (Du and Yip 1993). Our analysis of the Korean sample revealed that it contained the specific haplogroups A5 and M7a1, which are also prevalent in South Koreans (Kivisild et al. 2002, and references therein), thus revealing a common matrilineal genetic background of the Koreans in China and in the Korean Peninsula.
The Daurs were said to be the descendants of Khitan people in Liao Dynasty (916 AD–1125 AD). Other hypotheses suggested that their ancestors could be traced back to the local people in northern Heilongjiang Province and to some tribes of the Heishui region (same province) during the Shui and Tang Dynasty (581 AD–907 AD) (Du and Yip 1993). Our results demonstrated that Daur contained a high amount of the haplogroups prevalent in northern China (>46%), thus consistent with their northern origin.
According to historical documents, the Ewenkis traced their origin to the populations who lived around Lake Baikal and adjacent eastern regions more than 2,000 years ago, and were divided into three long-separated branches (Solon, Tungus, and Yakut; Du and Yip 1993). Our results supported the suggestion that the Ewenki is a typically northern population, for more than 63% of its maternal components are composed of haplogroups D, G, C, and Z. Furthermore, the high frequencies of haplogroups C and D but lower haplotype diversity (0.956±0.011; Table 4) observed in Ewenki sample suggested that Ewenki might have undergone recurrent genetic drifts because of its small population size (about 26,000, 1990 census) and episodes of population fragmentation during its development (Du and Yip 1993).
The Oroqens were regarded as the earliest inhabitants who lived in the Heilong River valley (Du and Yip 1993). The Oroqen population size experienced serious reduction during the past ten decades: the population size was about 18,000 in 1895, but was reduced to 2,256 by 1953. In 1990, the sample size increased to 6,965 (Du and Yip 1993). The genetic structure of our Oroqen sample showed concordant features with its small population size and recorded history: high frequencies (86.4%) of north-prevalent haplogroups, such as D, G, C, and Z, were found, but with low haplotype diversity (0.948±0.015; Table 4).
The formation and development of the Mongolian population was a complex process affected by integrating many Turkic-speaking tribes and some ethnic groups such as Han, Manchu, and Daur. Although the south-prevalent hapologroups F, R9b (formerly R10 in Yao and Zhang, 2002; c.f. Kong et al. 2003), and N9a were found with a low frequency in the Mongolian sample, the main maternal components of the population are composed of the north-prevalent haplogroups, which occupy more than 58% of the total samples.
In short, we identified another coding region segment (region 14576–16047) that is informative for discerning the haplogroup status of East Asian mtDNAs besides the previously reported ones (Yao et al. 2002a). Although the matrilineal genetic components of the five northern Chinese ethnic populations differed, the observed genetic profile was in general consistent with that of the Chinese Han regional samples (Yao et al. 2002a). The presence of northern population prevalent haplogroups D, G, C, and Z in these populations gave direct information in supporting their northern origin. Therefore, the matrilineal structures of these five northern Chinese ethnic populations reflected both the regional features and their ethnohistory.
References
Andrews RM, Kubacka I, Chinnery PF, Lightowlers RN, Turnbull DM, Howell N (1999) Reanalysis and revision of the Cambridge reference sequence for human mitochondrial DNA. Nat Genet 23:147
Bandelt H-J, Lahermo P, Richards M, Macaulay V (2001) Detecting errors in mtDNA data by phylogenetic analysis. Int J Legal Med 115:64–69
Bandelt H-J, Quintana-Murci L, Salas A, Macaulay V (2002) The fingerprint of phantom mutations in mtDNA data. Am J Hum Genet 71:1150–1160
Bandelt H-J, Herrnstadt C, Yao Y-G, Kong Q-P, Kivisild T, Rengo C, Scozzari R, Richards M, Villems R, Macaulay V, Howell N, Torroni A, Zhang Y-P (2003) Identification of Native American founder mtDNAs through the analysis of complete mtDNA sequences: some caveats. Ann Hum Genet (in press)
Cooper DN, Nussbaum RL, Michael Krawczak M (2002) Proposed guidelines for papers describing DNA polymorphism-disease associations. Hum Genet 110:207–208
Ding Y-C, Wooding S, Harpending H, Chi H-C, Li H-P, Fu Y-X, Pang J-F, Yao Y-G, Xiang YJG, Moyzis R, Zhang Y-P (2000) Population structure and history in East Asia. Proc Natl Acad Sci USA 97:14003–14006
Du R, Yip VF (1993) Ethnic groups in China. Science Press, Beijing
den Dunnen JT, Antonarakis SE (2001) Nomenclature for the description of human sequence variations. Hum Genet 109:121–124
Finnilä S, Lehtonen MS, Majamaa K (2001) Phylogenetic network for European mtDNA. Am J Hum Genet 68:1475–1484
Herrnstadt C, Elson JL, Fahy E, Preston G, Turnbull DM, Anderson C, Ghosh SS, Olefsky JM, Beal MF, Davis RE, Howell N (2002) Reduced-median-network analysis of complete mitochondrial DNA coding-region sequences for the major African, Asian, and European haplogroups. Am J Hum Genet 70:1152–1171; 71:448–449 (erratum)
Karafet T, Xu L, Du R, Wang W, Feng S, Wells RS, Redd AJ, Zegura SL, Hammer MF (2001) Paternal population history of east Asia: sources, patterns, and microevolutionary process. Am J Hum Genet 69:615–628
Kivisild T, Tolk H-V, Parik J, Wang Y, Papiha SS, Bandelt H-J, Villems R (2002) The emerging limbs and twigs of the East Asian mtDNA tree. Mol Biol Evol 19:1737–1751
Kong Q-P, Yao Y-G, Sun C, Bandelt H-J, Zhu C-L, Zhang Y-P (2003) Phylogeny of East Asian mitochondrial DNA lineages inferred from complete sequences. Am J Hum Genet (in press)
Lee SD, Lee YS, Lee JB (2002) Polymorphism in the mitochondrial cytochrome B gene in Koreans: an additional marker for individual identification. Int J Legal Med 116:74–78
Nei M (1987) Molecular evolutionary genetics. Columbia University Press, New York
Oota H, Kitano T, Jin F, Yuasa I, Wang L, Ueda S, Saitou, Stoneking M (2002) Extreme mtDNA homogeneity in continental Asian populations. Am J Phys Anthropol 118:146–153
Pakendorf B, Wiebe V, Tarskaia LA, Spitsyn VA, Soodyall H, Rodewald A, Stoneking M (2003) Mitochondrial DNA evidence for admixed origins of central Siberian populations. Am J Phys Anthropol 120:211–224
Povey S, Lovering R, Bruford E, Wright M, Lush M, Wain H (2001) The HUGO Gene Nomenclature Committee (HGNC). Hum Genet 109:678–680
Richards M, Macaulay V, Hickey E, Vega E, Sykes B, Guida V, Rengo C, Sellitto D, Cruciani F, Kivisild T, Villems R, Thomas M, Rychkov S, Rychkov O, Rychkov Y, Golge M, Dimitrov D, Hill E, Bradley D, Romano V, Cali F, Vona G, Demaine A, Papiha S, Triantaphyllidis C, Stefanescu G, Hatina J, Belledi M, Di Rienzo A, Novelletto A, Oppenheim A, Norby S, Al-Zaheri N, Santachiara-Benerecetti S, Scozari R, Torroni A, Bandelt H-J (2000) Tracing European founder lineages in the Near Eastern mtDNA pool. Am J Hum Genet 67:1251–1276
Rozas J, Rozas R (1999) DnaSP version 3: an integrated program for molecular population genetics and molecular evolution analysis. Bioinformatics 15:174–175
Silva Jr WA, Bonatto SL, Holanda AJ, Ribeiro-Dos-Santos AK, Paixão BM, Goldman GH, Abe-Sandes K, Rodriguez-Delfin L, Barbosa M, Paçó-Larson ML, Petzl-Erler ML, Valente V, Santos SEB, Zago MA (2002) Mitochondrial genome diversity of Native Americans supports a single early entry of founder populations into America. Am J Hum Genet 71:187–192
Tzen C-Y, Wu T-Y, Liu H-F (2001) Sequence polymorphism in the coding region of mitochondrial genome encompassing position 8389–8865. Forensic Sci Int 120:204–209
Yao Y-G, Zhang Y-P (2002) Phylogeographic analysis of mtDNA variation in four ethnic populations from Yunnan Province: new data and a reappraisal. J Hum Genet 47:311–318
Yao Y-G, Zhang Y-P (2003) Pitfalls in the analysis of ancient human mtDNA. Chinese Sci Bull 48:826–830
Yao Y-G, Lü X-M, Luo H-R, Li W-H, Zhang Y-P (2000a) Gene admixture in the silk road of China: evidence from mtDNA and melanocortin 1 receptor polymorphism. Genes Genet Syst 75:173–178
Yao Y-G, Watkins WS, Zhang Y-P (2000b) Evolutionary history of the mtDNA 9-bp deletion in Chinese populations and its relevance to the Peopling of East and Southeast Asia. Hum Genet 107:504–512
Yao Y-G, Yuan Z-G, Zhou Z-D, Geng P-L, Li Q-W, Zhang Y-P (2001) Frequency of the mtDNA 9-bp deletion among Chinese ethnic groups. Prog Nat Sci 11:358–364
Yao Y-G, Kong Q-P, Bandelt H-J, Kivisild T, Zhang Y-P (2002a) Phylogeographic differentiation of mitochondrial DNA in Han Chinese. Am J Hum Genet 70:635–651
Yao Y-G, Nie L, Harpending H, Fu Y-X, Yuan Z-G, Zhang Y-P (2002b) Genetic relationship of Chinese ethnic populations revealed by mtDNA sequence diversity. Am J Phys Anthropol 118:63–76
Yao Y-G, Kong Q-P, Man X-Y, Bandelt H-J, Zhang Y-P (2003a) Reconstructing the evolutionary history of China: a caveat about inferences drawn from ancient DNA. Mol Biol Evol 20:214–219
Yao Y-G, Macaulay V, Kivisild T, Zhang Y-P, Bandelt H-J (2003b) To trust or not to trust an idiosyncratic mitochondrial data set. Am J Hum Genet 72:1341–1346
Yao Y-G, Macaulay V, Kivisild T, Zhang Y-P, Bandelt H-J (2003c) Reply to Silva et al. Am J Hum Genet 72:1348–1349
Yao Y-G, Kong Q-P, Sun C, Zhang Y-P (2003d) Can the occurrence of rare insertion/deletion polymorphisms in human mtDNA be verified from phylogeny? Chinese Sci Bull 48:663–667
Acknowledgements
We thank Shi-Fang Wu for technical assistance and two anonymous reviewers for their critical comments on the manuscript. We also thank Dr. Brigitte Pakendorf for providing us the sequence data. The work was supported by Natural Science Foundation of China (NSFC) and Natural Science Foundation of Yunnan Province.
Author information
Authors and Affiliations
Corresponding author
Electronic database information
Electronic database information
Accession numbers and URLs for the sequence data of mtDNA control region (including HVS-I and HVS-II) and the region 14576–16047 in this article are as follows: GenBank, http://www.ncbi.nlm.nih.gov/web/Genbank (accession numbers: AY243965–AY244330 and AY243877–AY243939).
Rights and permissions
About this article
Cite this article
Kong, QP., Yao, YG., Liu, M. et al. Mitochondrial DNA sequence polymorphisms of five ethnic populations from northern China. Hum Genet 113, 391–405 (2003). https://doi.org/10.1007/s00439-003-1004-7
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00439-003-1004-7