
Abstract
In plants, gametophytic apomixis is a form of asexual reproduction that leads to the formation of seed-derived offspring that are genetically identical to the mother plant. A common set of RFLP markers, including five rice anchor markers previously shown to be linked to apomixis in Paspalum simplex, were used to detect linkage with apomixis in P. notatum and P. malacophyllum. A comparative map of the region around the apomixis locus was constructed for the three Paspalum species, and compared to the rice map. The locus that controls apomixis in P. simplex was almost completely conserved in the closely related species P. malacophyllum, whereas it was only partially represented in the distantly related species P. notatum. Although strong synteny of markers was noted between this locus and a portion of rice chromosome 12 in both P. simplex and P. malacophyllum, the same locus in P. notatum was localized to a hybrid chromosome which carries markers that map to rice chromosomes 2 and 12. All three Paspalum species showed recombination suppression at the apomixis locus; in the case of P. notatum, this might be due to a heterozygosity for a translocation that most probably negatively interferes with chromosomal pairing near the locus. A common set of markers that show linkage with apomixis in all three Paspalum species define a portion of the apomixis-controlling locus that is likely to contain genes critical for apomictic reproduction.
Similar content being viewed by others
Avoid common mistakes on your manuscript.
Introduction
The alternation of gametophytic and sporophytic cycles and the fusion of parental gametes provide the major sources of genetic variability in the majority of organisms, thereby ensuring the adaptation of species to varying conditions by the production of new allelic combinations. However, some plants have evolved an alternative reproductive system that circumvents both meiosis and fertilization, giving rise to seed-derived progenies that are genetically identical to the mother plant. This type of clonal reproduction through seeds is defined as apomixis (Nogler 1984). With rare exceptions (Pichot 2001), apomictic reproduction is specific for the female gametophyte, whereas the male gametophyte in apomictic plants gives rise to normal haploid pollen. This normal pollen can fertilize occasional meiotic egg cells that arise in apomictic plants, thereby maintaining a sort of reservoir of genetic variability in otherwise clonal, apomictic populations. The potential benefits of apomixis to agriculture are enormous, and vary from the full exploitation of heterosis by reseeding the hybrid seed without loss of hybrid vigor to the possibility of synthesizing environmentally safe transgenic varieties when apomixis is induced in a male-sterile background (Ramulu et al. 1999; Daniell 2002). However, with the exception of some fruit trees and selected forage grasses, no apomictic crops have been reported to date, and attempts to transfer the apomictic trait from wild relatives to crops by sexual crosses have been largely unsuccessful (Vielle-Calzada et al. 1996). As a consequence, research efforts are now focused on the induction of artificial apomixis in mutagenized sexual models, and on understanding the molecular mechanisms of apomixis in wild apomictic species. Both strategies are directed towards the transfer of gene(s) for apomixis to crop plants through genetic engineering.
The grass genus Paspalum is composed of more than 400 species, which are adapted to several environments, and is characterized by an extraordinary variety of reproductive systems including allogamy (due to self-incompatibility systems), autogamy (cleistogamy) and apomixis. In general, diploid species are sexual, whereas polyploids are apomictic (Quarin 1992). Among the different Paspalum species, P. simplex Morong and P. notatum Flügge are the most widely studied in the context of the molecular genetics of apomixis. Despite the fact that these two species are genetically rather remote from each other, they share several characteristics related to apomixis, including the co-occurrence of sexual, self-incompatible, diploid cytotypes with sexually compatible polyploid apomictic individuals. The development of unreduced female gametophytes is similar, and occurs through apospory, whereby the megaspore mother cell usually degenerates and a cell from the nucellus gives rise to a single or multiple embryo sacs. The genetic control of apomixis in both species is consistent with the involvement of a single-dose dominant allele. However, strongly distorted segregation ratios, always skewed in favor of sexual genotypes, have been detected in segregating populations of both species, suggesting that this gene has a lethal pleiotropic effect with incomplete penetrance or, alternatively, that it is linked to a lethal factor which causes gamete death when present in the homozygous state (Pupilli et al. 2001; Martinez et al. 2001). Some additional characteristics of these two species, and of P. malacophyllum Trin., are summarized in Table 1. The apomixis-controlling-locus (ACL) of P. simplex has been localized to a relatively large chromosomal segment that is characterized by strong repression of recombination and synteny of markers with the telomeric portion of the long arm of rice chromosome 12 (RC12) (Pupilli et al. 2001). We have identified five rice markers linked to apomixis in P. simplex which span 14.5 cM in the rice map and, accordingly, we hypothesized that the size of the ACL in P. simplex is similar or even larger. If it is assumed that 1 cM corresponds roughly to 1 Mb in model species, then the physical size of the ACL of P. simplex would be at least 15 Mb. In the absence of recombination, physical dissection of ACL by cloning and direct sequencing of large DNA fragments is the only map-based strategy available to isolate the possible genetic determinants of apomixis in this species. Apomixis-specific SCARs (Sequence Characterized Amplified Markers) have already been isolated in P. simplex (Busti et al., unpublished) and have been used to screen a BAC library of the same species (Calderini et al., unpublished). Partial sequencing of positive BACs reveals a high density of non-coding regions, together with transposable elements and rare genes. Large-scale sequencing of all BACs encompassing the ACL in P. simplex would therefore be a very laborious, expensive, and largely unprofitable effort. The identification of sub-regions of the ACL that are conserved among different apomictic species may facilitate the selection of BACs that are more likely to contain candidate genes for apomixis. Recently, in an independent study, one of the five rice markers linked to apomixis in P. simplex was also found to be linked to apomictic reproduction in P. notatum (Martinez et al. 2004).
The objective of the present research was to determine by comparative mapping whether there are conserved areas of the ACL in P. simplex that are correlated with apomixis in the distantly related species P. notatum. For these experiments, the closely related species P. malacophyllum was used as a control, in which at least partial conservation of the ACL of P. simplex would be expected. More specifically, our objectives were: (1) to identify a common set of molecular markers that show linkage with apomixis in three Paspalum species; (2) to investigate the extent to which the synteny with rice markers detected in P. simplex at the ACL is conserved in the differently related species; and (3) to estimate the genetic length of the ACL in P. notatum on the basis of the linkage of fragments segregating from the sexual parent that are orthologous to those linked to apomixis.
Materials and methods
Plant materials
The mapping population of P. simplex described in Pupilli et al. (2001) was screened for close linkage with apomixis using additional rice markers mapped to the telomeric portion of the long arm of RC12. A population of 89 plants (43 apomictic and 46 sexual) belonging to an F1 population (Martinez et al. 2004) was used for comparative mapping of apomixis in P. notatum. An apomictic genotype of P. malacophyllum (TK2449) was crossed with the same sexual parent used to generate the P. simplex mapping population, as previously reported (Martinez et al. 2001). Thirty-six F1 individuals were generated from this cross, and their apomictic or sexual phenotypes were analyzed by molecular marker-assisted progeny tests as reported for the mapping population of P. simplex (Pupilli et al. 2001). The mode of reproduction of the P. notatum individuals was assessed by cytological and embryological analysis (Martinez et al. 2001) and corroborated using molecular markers (Ortiz et al. 1997).
Experimental procedures
Genomic DNA was extracted from the P. notatum individuals and from P. simplex X P. malacopyllum F1s according to Ortiz et al. (1997), and from P. simplex according to Pupilli et al. (2001). Genomic DNA (7–10 μg) from each parental line used in the three crosses was digested with Eco RI, Hin dIII, Bam HI, Pst I, Eco RV or Bg lII for detection of informative polymorphisms (i.e. bands present in the apomictic and absent in the sexual parental line, or vice versa). The digested DNAs were electrophoresed in agarose gels, blotted onto filters, hybridized with 32P-labeled probes, washed, and exposed to X-ray films as previously described (Pupilli et al. 2001). All the rice probes belonged to the New Landmarker Set (Nagamura et al. 1997) or were mapped subsequently to the areas of interest on chromosomes 2 and 12, as reported in a more recent version of the rice map constructed by the Rice Genome Project (RGP), Japan (Harushima et al. 1998). The oat probe CDO507, which belongs to a set of anchor markers for comparative mapping in grasses (Van Deynze et al. 1998), was developed at Cornell University (Ithaca, N.Y.). Inserts were amplified from recombinant plasmids by PCR with M13 (forward and reverse) universal primers. Amplified inserts were checked for molecular weight by electrophoresis, and column-purified with the Jet Quick Purification kit (Genomed). AFLPs linked to apomixis in P. simplex were converted into RFLP probes as described by Labombarda et al. (2002).
Data analysis
Linkage relationships were established only for alleles segregating from the same parental line as single-dose restriction fragments (SDRFs). Similarly, apomixis was considered as an SDRF in the mapping population, and its presence was assigned to the heterozygous dominant and its absence to the homozygous recessive genotype. Linkage and map order were calculated with MAPMAKER/EXP3 (Lander et al. 1987; Lincoln et al. 1992). Data were analyzed by the F2 backcross option, and linkage was detected with a LOD score of 3. Recombination frequencies were converted to map distances (cM) using the Haldane mapping function. The recombination frequencies for markers putatively linked in the repulsion phase were calculated according to the formula r =[3( a + d)/ n] (Hackett et al. 1998), where r is the recombination frequency, a + d the sum of recombinants and n is the total number of progeny analyzed.
Results
Screening of markers located on RC12 to detect linkage with apomixis in three Paspalum species
All five rice probes, C901, R1759, C996A, C1069 and C454 (Pupilli et al. 2001), together with two homologous hemizygous probes derived from AFLPs, EM180 (Labombarda et al. 2002) and B11 (Busti at al., unpublished), linked to apomixis in P. simplex, were used to detect informative polymorphisms between the parental lines of P. notatum and P. simplex X P. malacophyllum F1s. Subsequently, the DNAs of a subpopulation of P. notatum made up of 10 apomictic and 10 sexual plants, together with all the 36 F1s of the P. simplex X P. malacophyllum population, were digested with appropriate restriction enzymes, and screened for co-segregation of informative markers with apomixis. For both parent combinations, at least one restriction enzyme detected informative polymorphisms with all the probes used, but of these, only four (C996A, C1069, C454 and EM180), detected hybridizing fragments linked 100% in coupling phase with apomixis in P. notatum. When the same probes were hybridized to the DNA of the parental lines of the P. simplex X P. malacopyllum F1, three rice probes (R1759, C1069 and C454) and both genomic probes detected hybridizing fragments in the P. malacophyllum parental line that were identical to those linked to apomixis in P. simplex. C996A revealed a fragment of lower molecular weight (data not shown). These fragments were inherited by only six F1s, suggesting that these plants were apomictic. Progeny test analyses, combined with the use of molecular markers, on these six plants, together with 10 other F1s as controls, confirmed that only the former set had inherited the apomixis trait from the pollinating parent (data not shown). Since the rice probes C1069 and C454 were the only ones that detected almost identical (in molecular weight) hybridizing fragments linked to apomixis in all the three species analyzed, new probes from the rice region near these markers were further explored for close linkage with apomixis in P. simplex. Fifteen markers were utilized that mapped to a 7.8-cM area extending from marker E31743 to E60272 in rice (http://rgp.dna.affrc.go.jp/publicdata/estmap2001/Chr12.html), in addition to the markers C454 and C1069. These markers showed variable extents of homology with P. simplex DNA, but only two of these, namely C60087 and R2241, which mapped next to C454 and C1069, were linked to apomixis. When hybridized to P. notatum DNA, these two markers were found to be unlinked to apomixis, whereas both detected apomixis-linked fragments in P. malacophyllum. It thus appeared that at least part of the ACL of P. simplex was deleted in P. notatum, whereas the same locus was almost intact in P. malacophyllum. However, since only three rice markers out of the seven linked to apomixis in P. simplex were also apomixis-related in P. notatum, it cannot be definitively concluded that the ACL of P. notatum is located on the equivalent of the telomeric portion of the long arm of RC12 as it is in P. simplex.
Genome-wide screening of rice markers to detect further linkage with apomixis in P. notatum
To investigate whether the conserved block identified by the markers C454, C1069 and C996A might have been translocated to another chromosome in P. notatum, another set of 20 markers was screened for informative polymorphisms between the parental lines of the P. notatum mapping population. These markers were selected from the New Landmarker Set of RGP on the basis of the following criteria: (1) they must show homology to the DNA of P. simplex and other turf grasses (Caceres et al. 2000; Busti et al. 2004), and (2) they must represent all rice chromosomes. In the subset chosen, rice chromosomes 1 and 5 were represented by three markers each, chromosomes 2, 8, 4 and 11 by two markers, and chromosomes 3, 9, 6, 7, 10 and 12 by one marker each. These probes showed various extents of homology to the DNA of P. notatum, and eight detected informative polymorphisms. When these probes were hybridized to the digested DNAs of the 20-member reference population, only C560 detected a fragment that was 100% linked to apomixis in coupling phase (Fig. 1a). All informative probes were also hybridized to the digested DNA of the P. simplex X P. malacophyllum population, but none detected fragments that co-segregated with the six apomictic F1s. This indicates that, although the ACL of both P. malacophyllum and P. simplex have retained strong synteny with their homoeologous counterpart on RC12, the locus in P. notatum is rearranged. Since the probe C560 mapped close to the telomeric end of the long arm of rice chromosome 2 (RC2), its linkage with apomixis and the three markers from RC12 suggests that a translocation has indeed occurred in the ACL of P. notatum.

Hybridization pattern obtained with four rice probes and one (EM180) P. simplex homologous probe in a 20-plant reference population of P. notatum segregating for apomixis. The genomic DNAs were digested with Hin dII ( a, c , d and e) or with Eco RI ( b). The fragments that co-segregate with apomixis are indicated by arrows. F, female sexual parental line, M, male apomictic parental line, APO, 10 apomictic F1s; SEX, 10 sexual F1s
Comparison of the ACL in the three Paspalum species
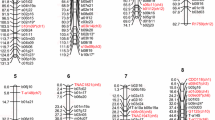
To confirm that part of the P. notatum chromosome that is homoeologous to the RC2 belonged to the ACL, another set of rice probes which mapped near C560 was screened for linkage with apomixis in P. notatum. This set was composed of five probes from the New Landmarker Set that had not previously been analyzed (C1221, C1408, R3393, C1445 and C1470) and which span 38.9 cM from the most telomeric (C1470) to the most centromeric marker (R3393), together with another seven markers covering 11.2 cM from C560 towards the telomere (C253, C1119, C348 and C978), and another 13.7 cM towards the centromere (R2609, C932 and C520). In addition, the oat probe CDO507, which is located approximately 8 cM apart from C560 in the integrated RGP, RGN and IRRI map of rice (http://www.grs.nig.ac.jp/rice/oryzabase/maps/map.jsp), was also used. Of these, only six markers (C1445, C1408, CDO507, C348, C253 and C932) revealed informative polymorphisms between the parental lines of the mapping population. When these probes were hybridized with the DNAs of the 20-plant reference population, C1408 showed a faint hybridizing pattern identical to that of C560, suggesting that these two probes may recognize the same locus with different levels of homology. C932 detected a hybridizing fragment linked to apomixis (Fig. 1b), while the other probes did not reveal any fragment linked to apomixis. In summary, five rice probes (three from RC12 and two from RC2), together with one homologous P. simplex marker, showed complete linkage with apomixis in the 20-plant reference population of P. notatum. To confirm their tight linkage with apomixis, these probes were hybridized with DNAs of the entire mapping population of 89 F1s. All the fragments that co-segregated with apomixis in the reference population were 100% linked in coupling phase to apomixis in the larger population, indicating that the ACL of P. notatum is characterized by repression of recombination. In no case was repulsion-phase linkage with apomixis detected. To investigate whether the two apomixis-linked markers of P. notatum located in RC2 were also linked to apomixis in the other species under study, the corresponding probes were hybridized with the digested DNAs of the apomictic and sexual genotypes P. simplex and P. malacophyllum. No co-segregation with apomixis was detected, indicating that the homoeology between ACL and RC2 is specific for P. notatum. The results of comparative mapping of apomixis in the three species analyzed are depicted in Fig. 2, and enable us to focus on the ACL of P. simplex in comparison with those of P. malacophyllum and P. notatum. In particular, the portion of the ACL of P. simplex that corresponds to the more telomeric position in rice progressively lost its linkage with apomixis as the genetic distances among the species increases. This may indicate that the genetic determinants of apomixis are likely to be located in the part of the ACL of P. simplex more distal to the telomere, and that the linkage of apomixis with the telomeric markers is a consequence of recombination suppression rather than the presence of critical genes in the corresponding area. The presence of the residual “centromeric” ACL of P. simplex within the equivalent locus of P. notatum appears to block recombination in a chromosome region that is unrelated to RC12.

Alignment of the ACL in three Paspalum species in relation to its homoeologous rice counterpart. The markers indicated in bold are conserved in the ACL of the three Paspalum species. Genetic distances are expressed in cM
Reconstruction of the sexual chromosome homologous to the one that bears the apomixis-controlling locus in P. notatum
Since the sexual parent of the P. notatum mapping population was a heterozygous hybrid genotype (Martinez et al. 2001), we expected to find markers segregating as SDRFs from this parent. This would allow reconstruction of the ancestral sexual chromosome that was homologous to the one that carries the apomixis-controlling locus, and permit us to address the following questions. First, is recombination between the apomixis-linked markers of P. notatum blocked in the sexual background, and if not, can the genetic size of ACL be estimated based on the linkage of these markers in the sexual parental line? Second, have these markers lost their synteny with respect to their rice counterparts in sexual genotypes, or is the translocation event we detected specific for the apomictic cytotype? We approached these questions by hybridizing the DNAs of 89 F1 P. notatum individuals with an additional set of probes, including the two markers from RC12 (C901 and R1759) that were linked to apomixis in P. simplex, and four markers of RC2 (C1445, C1222, R3393 and CDO507) which were located near the ACL of P. notatum but were nonetheless unrelated to apomixis. The fragments segregating from the sexual parent enabled the identification of two linkage groups (Fig. 3). One of these was homoeologous to the telomeric portion of the long arm of RC12 (P. notatum sex 12), while the other was homoeologous to the subtelomeric region of the long arm of RC2 (P. notatum sex 2). The colinearity of these markers and the distances between them were highly conserved between RC12 and its homoeologs in P. notatum and, to a lesser extent, between RC2 and its counterpart. EM180 mapped near the markers C454 and C1069, whereas C996A could not be mapped since it did not reveal any segregating fragment from the sexual parent. However, although the two markers C1069 and C454 did not recombine in P. notatum —probably a result of their tight linkage as in rice—EM180 and these two markers did show recombination. Similarly, the markers C560 and C932 also recombined in sexual P. notatum chromosomes. Assuming that the genetic distance of about 6 cM between C996A and C1069 and C454 in rice is conserved in P. notatum, it can be estimated that the genetic size of the ACL in P. notatum is about 8–10 cM. The probe CDO507 gave a strongly hybridizing pattern with two fragments segregating from the apomictic parent and one from the sexual parent. The former fragments segregated independently of apomixis, and the latter was mapped at 5.2 cM from C932 in the sexual counterpart of the ACL, a distance that is comparable to that reported in the integrated rice map (http://www.grs.nig.ac.jp/rice/oryzabase/maps/map.jsp). This implies a high level of gene conservation between P. notatum and rice, at least in this area. However, the most striking information derived from linkage analyses of the sexual counterparts of the ACL is that a chromosomal block identified by the markers C1069, C454, C996A and EM180 is no longer syntenic with the other markers from RC12. A new linkage relationship with markers of a different chromosomal background has thus been established during the evolution from a sexual to an apomictic cytotype within the same P. notatum species.

Reconstruction of the homologous sexual and apomictic chromosomes in P. notatum in relation to their respective homoeologous counterparts in rice. Genetic distances are expressed in cM
To summarize, it appears evident from comparative mapping of apomixis in Paspalum that only a relatively small portion of the large ACL of P. simplex is linked to apomictic reproduction among different Paspalum species. The high level of conservation of this area detected at the map level should also exist at the sequence level in at least some regions, as is evidenced by the nearly identical polymorphism revealed by the marker C1069 (Fig. 4) between apomictic and sexual genotypes of the three species analyzed.

Hybridization pattern of the apomixis-linked rice EST C1069 in three Paspalum species. The genomic DNAs were digested with Eco RI. F, female sexual parental line; M, male apomictic parental line; APO, three apomictic F1s; SEX, three sexual F1s
Discussion
Comparative mapping efforts have revealed various levels of conservation in gene order within the grass family (Bennetzen and Freeling 1993). Moore et al. (1995) showed that all the maps of Gramineae can be combined into a single map with rice as a base genome. The ongoing development of molecular tools available for this model species, in addition to the recent release of the complete sequence of the rice genome (Goff et al. 2002; Yu et al. 2002), has made possible the isolation of genes of interest once the homoeologous region in which the gene is located has been identified using molecular markers conserved between species. In fact, one would expect to be able to isolate any locus of interest if it is located in a chromosomal region in which appropriate markers and genes are abundant. In general, rice chromosomes are richer in genes compared to their homoeologous counterparts in other grasses (Moore et al. 1997). The effectiveness of comparative map-based cloning methods would be confirmed if the colinearity of marker order (macro-colinearity) is conserved at the sequence level (micro-colinearity). Although some published data appear to satisfy this condition, (Chen et al. 1997; Feuillet and Keller 1999), a number of other studies have revealed significant gene rearrangements within otherwise macro-colinear regions; for example, in a comparison of the Adh1 locus between maize, sorghum, and rice (Tikhonov et al. 1999; Tarchini et al. 2000) and of the disease resistance gene rpg1 between barley and rice (Kilian et al. 1997). Overall, recent comparative sequence analyses of large BAC or YAC fragments from barley, maize, rice, sorghum and wheat have shown a surprisingly high frequency of local rearrangements (small inversions or duplications), in addition to changes in gene content, order and orientation (Bennetzen and Ramakrishna 2002). Although some types of rearrangements, such as small inversions or gene duplication, are not expected to have large effect on micro-colinearity, even small-scale deletions and translocations can greatly hamper map-based gene cloning using rice as a model species. For this reason, as Feuillet and Keller (2002) have pointed out, “...approaches based on colinearity between grass genomes must also be performed using more closely related species, e.g. within tribes or subtribes”.
On the basis of these considerations, we decided to investigate the conservation of marker order in the ACL among different Paspalum species first. The results can then be referred back to the rice map in order to undertake the map-based cloning of the apomixis locus in the homoeologous region of rice. Although P. simplex retains a remarkable level of marker synteny between its ACL and the telomeric region of the long arm of RC12, the majority of the additional rice markers that map to the same area were not linked to apomixis, with the exception of the two closest to the apomixis-linked markers C1069 and C454. This indicates that although no gross rearrangements occurred in this area when rice and Paspalum diverged, a relatively large number of small-scale rearrangements may have occurred subsequently. Both small- and large-scale rearrangements were detected at the ACL of Paspalum and the extent of variation was related to the genetic distances between the species analyzed. The hybridization patterns revealed by the rice probes R1759, C1069 and C454 and the two AFLP-derived, homologous, hemizygous probes detected identical apomixis-linked fragments in the P. simplex and P. malacophyllum genomes, whereas the probe C996A showed apomixis-linked bands of different molecular weights. This would indicate the occurrence of small-scale rearrangements which are associated with gain and/or loss of restriction sites at this locus. When the structure of the ACL was compared between P. simplex and P. notatum, both small- and large-scale rearrangements were detected: small-scale rearrangements were revealed as minor (C1069, Fig. 4) or larger (C454 and C996, not shown) differences in molecular weights of the corresponding apomixis-related bands. The large-scale rearrangements were due to the loss of synteny of the distal markers R1759 and C901 with the other apomixis-linked markers of RC12, and the gain of two other RC2 markers. Linkage analysis of the homologous sexual counterpart of the ACL in P. notatum showed strict colinearity of markers with the telomeric region of the long arm of RC12, including those markers (C1069 C454 and C996A) that were linked to markers of RC2 in apomictic P. notatum. This indicates that the large-scale rearrangements related to apomictic reproduction in P. notatum had not yet taken place when P. notatum and P. simplex diverged, but more probably occurred intraspecifically during the recent evolution of Paspalum.
Several mechanisms have been proposed to explain the suppression of recombination at the apomixis locus in Pennisetum squamulatum (Ozias-Akins et al. 1998; Goel et al. 2003), P. simplex (Labombarda et al. 2002) and Tripsacum dactyloides (Grimanelli et al. 1998). A recurrent feature of these hypotheses is that structural rearrangements occurring in the vicinity of the ACL have produced dissimilarity in the sequences of homologous chromosomes, thereby producing a negative effect on recombination. Among these rearrangements, heterozygous inversions and translocations are known to affect the products of crossing-over occurring in the vicinity of these events. In a recent report, Goel et al. (2003) suggested that an inversion at the apomixis locus might have occurred when the two related species P. squamulatum and Cenchrus ciliaris diverged, since the position of this locus relative to centromeric sequences differs in the two species. The ability to compare the structure of the ACL in sexual and apomictic cytotypes of P. notatum allowed us to detect, for the first time, a heterozygous translocation event in apomictic genotypes that probably accounts for the repression of recombination in the vicinity of this locus. On the other hand, since marker synteny is strictly conserved between rice and the ACL of P. simplex, no translocations were detected in this species. However, heterozygous inversion(s) that change the relative positions of the markers within this locus cannot be ruled out, although such events cannot be detected in Paspalum since physical mapping has not yet been performed. Inversion of part of the ACL has already been proposed in P. simplex to explain the lack of allelism or even repulsion-phase linkage between the apomixis-linked RFLP fragments and their putative sexual counterparts segregating from the apomictic parent (Pupilli et al. 2001). It remains to be determined if the translocation that generated the hybrid chromosome containing the apomixis locus in P. notatum was due to the relocation of the homoeolog of RC12 onto the equivalent of RC2, or if the segment identified by two markers on chromosome 2 translocated onto a chromosome12 background. Within the context of the present study, no direct evidence can be provided to demonstrate which hypothesis is valid, although the evolution of one of the two homologous APO-related sequences of P. simplex in P. notatum may hold important clues. These sequences were isolated in P. simplex from the hemizygous region of the ACL, and represent unique alleles in its genome. Of these, only EM180 was linked to apomixis in P. notatum, where it has undergone amplification. As shown in Fig. 1e, two apomixis-related fragments are detectable, which according to the definition of Bennetzen (2000) can be recognized as the orthologs of P. simplex EM180, together with at least two other fragments of 3 and 6 kb which segregate independently of apomixis from the apomictic parent (and hence can be defined as paralogous). Recent studies using comparative analysis of gene families suggest that genetic amplification and gene mobility may be synergistically associated. In particular, gene copies that are translocated to non-orthologous positions are prone to being amplified, generating multiple, paralogous copies (Song et al. 2002). This might be one mechanism that led to amplification of EM180 in the Paspalum lineage. One or a few copies of this sequence could have been translocated to a non-orthologous location during the evolution of the ACL from P. simplex - sexual P. notatum - apomictic P. notatum. This would generate various copies of paralogous sequences scattered throughout the genome by amplification. Sequence analysis of EM180 revealed it is a non-coding region that most probably belongs to an intergenic region. In general, such regions do not show sequence conservation among grasses (Bennetzen 2000). However, in this case the forces that induce sequence divergence are blocked by the recombination suppression at the ACL. Since EM180 was mapped to the homoeologous region of RC12 in both apomictic P. simplex and sexual P. notatum, we believe that this region translocated to the homoeologous region of RC2 in P. notatum. In addition, on the basis of the available sequence of the highly conserved and apomixis-linked marker C1069, BLAST analysis of the predicted peptide showed significant similarity with a mutator-like transposase that might have played a role in the excision of the conserved ACL homoeologous to RC12. This marker, together with C454, C996A, and EM180 delimited a more restricted area of the ACL of P. simplex that showed linkage with apomixis in the three Paspalum spp. This block spans approximately 8–10 cM and is characterized by instability in P. notatum, where it has translocated to another chromosomal position. This translocation had two major effects: (1) it suppressed recombination in an otherwise recombinationally competent area in the vicinity of the insertion site, and (2) induced apomictic reproduction. The most straightforward hypothesis is that the genetic determinants contained in this block are necessary, but not sufficient, to induce apomictic reproduction in Paspalum spp. since this block should rearrange, at least to some extent.
In conclusion, the present study has identified a gene block that may be used for mining genes involved in apomicitc reproduction in Paspalum. The physical isolation of the BAC clones contained in this block is underway, and candidate genes will be identified that are involved in apomictic reproduction. Confirmation of their involvement can be demonstrated by analyses of their tissue-specific expression and their ability to induce apomixis-related phenotypes in transformed Paspalum and model sexual systems.
References
Bennett MD, Smith JB (1976) Nuclear DNA amounts in angiosperms. Philos Trans R Soc Lond B 274:227–274
Bennetzen JL (2000) Comparative sequence analysis of plant nuclear genomes: microcolinearity and its many exceptions. Plant Cell 12:1021–1029
Bennetzen JL, Freeling M (1993) Grasses as a single genetic systems: genome composition, colinearity and compatibility. Trends Genet 9:259–261
Bennetzen JL, Ramakrishna W (2002) Numerous small rearrangements of gene content, order and orientation differentiate grass genomes. Plant Mol Biol 48:821–827
Busti A, Caceres ME, Calderini O, Arcioni S, Pupilli F (2004) RFLP markers for cultivar identification in tall fescue ( Festuca arundinacea Schreb.) Genet Res Crop Evol, in press
Caceres ME, Pupilli F, Quarin CL, Arcioni S (1999) Feulgen densitometry of embryo sacs permits discrimination between sexual and apomictic plants in Paspalum simplex. Euphytica 110:161–167
Caceres ME, Pupilli F, Piano E, Arcioni S (2000) RFLP markers are an effective tool for the identification of creeping bentgrass ( Agrostis stolonifera L.) cultivars. Genet Res Crop Evol 47:455–459
Caceres ME, Matzk F, Busti A, Pupilli F, Arcioni S (2001) Apomixis and sexuality in Paspalum simplex: characterization of the mode of reproduction in segregating progenies by different methods. Sex Plant Reprod 14:201–206
Chen M, SanMiguel P, De Oliveira AC, Woo SS, Zhang H, Wing RA, Bennetzen JL (1997) Microcolinearity in Sh2 -homologous regions of the maize, rice and sorghum genomes. Proc Natl Acad Sci USA 94:3431–3435
Daniell H (2002) Molecular strategies for gene containment in transgenic crops. Nat Biotechnol 20:581–586
Feuillet C, Keller B (1999) High gene density is conserved at syntenic loci of small and large grass genomes. Proc Natl Acad Sci USA 96:8665–8670
Feuillet C, Keller B (2002) Comparative genomics in the grass family: molecular characterization of grass genome structure and evolution. Ann Bot 89:3–10
Goel S, Chen Z, Conner JA, Akiyama Y, Hanna WW, Ozias Akins P (2003) Delineation by fluorescence in situ hybridization of a single hemizygous chromosomal region associated with aposporous embryo sac formation in Pennisetum squamulatum and Cenchrus ciliaris. Genetics 163:1069–1082
Goff S, et al (2002) A draft sequence of the rice genome ( Oryza sativa L. ssp. japonica). Science 296:92–100
Grimanelli D, Leblanc O, Espinosa E, Perotti E, Gonzales de Leon D, Savidan Y (1998) Mapping diplosporous apomixis in tetraploid Tripsacum: one gene or several genes? Heredity 80:33–39
Hackett CA, Bradshaw JE, Mayer RC, McNicol, JW, Milbourne D, Waugh R (1998) Linkage analysis in tetraploid species: a simulation study. Genet Res 71:143–154
Harushima Y, et al (1998) A high density rice genetic linkage map with 2275 markers using a single F2 population. Genetics 148:479–494
Kilian A, Chen J, Han F, Steffenson B, Kleinhofs A (1997) Towards map-based cloning of the barley stem rust resistance genes Rpg1 and rpg4 using rice as an intergenomic cloning vehicle. Plant Mol Biol 35:187–195
Labombarda P, Busti A, Caceres ME, Pupilli F, Arcioni S (2002) An AFLP marker tightly linked to apomixis reveals hemizygosity in a portion of the apomixis-controlling locus in Paspalum simplex. Genome 45:513–519
Lander E, Green P, Abrahmson J, Barlow A, Daly M, Lincoln S, Newburg L (1987) MAPMAKER: an interactive computer package for constructing primary genetic linkage maps in experimental and natural populations. Genomics 1:171–181
Lincoln S, Daly M, Lander E (1992) Constructing genetic maps with MAPMAKER/EXP 3.0. Whitehead Institute Technical Report (3rd edn). Whitehead Institute, Cambridge, Mass.
Martinez EJ, Urbani MH, Quarin CL, Ortiz JPA (2001) Inheritance of apospory in bahiagrass, Paspalum notatum. Hereditas 135:19–25
Martinez EJ, Hopp HE, Stein J, Ortiz JPA, Quarin CL (2004) Genetic characterization of apospory in tetraploid Paspalum notatum based on the identification of linked molecular markers. Mol Breeding, in press
Moore G, Aragon-Alcaide L, Roberts M, Reader S, Miller T, Foote T (1997) Are rice chromosomes components of a holocentric chromosome ancestor? Plant Mol Biol 35:17–23
Moore SR, Devos KM, Wang Z, Gale MD (1995) Cereal genome evolution: grasses, line up and form a circle. Curr Biol 7:737–739
Nagamura Y, Antonio BA, Sasaki T (1997) Rice molecular genetic map using RFLPs and its applications. Plant Mol Biol 35:79–87
Nogler GA (1984) Gametophytic apomixis. In: Johri EM (ed) Embryology of angiosperms. Springer Verlag, Berlin pp 475–518
Ortiz JPA, Pessino S, Leblanc O, Hayward MD, Quarin CL (1997) Genetic fingerprinting for determining the mode of reproduction in Paspalum notatum: a subtropical apomictic forage grass. Theor Appl Genet 95:850–856
Ozias-Akins P, Roche D, Hanna WW (1998) Tight clustering and hemizygosity of apomixis-linked markers in Pennisetum squamulatum implies genetic control of apospory by a divergent locus that may have no allelic form in sexual genotypes. Proc Natl Acad Sci USA 95:5127–5132
Pichot C, El Maataoui M, Raddi S, Raddi P (2001) Surrogate mother for endangered Cupressus. Nature 412:39
Pupilli F, Labombarda P, Caceres ME, Quarin CL, Arcioni S (2001) The chromosome segment related to apomixis in Paspalum simplex is homoeologous to the telomeric region of the long arm of rice chromosome 12. Mol Breeding 8:53–61
Quarin CL (1992) The nature of apomixis and its origin in Panicoideae grasses. Apomixis Newslett 5:7–15
Quarin CL (1999) Effect of pollen source and pollen ploidy on endosperm formation and seed set in pseudogamous apomictic Paspalum notatum. Sex Plant Reprod 11:331–335
Ramulu KS, Sharma VK, Naumova TN, Dijkhuis P, van Lookeren Campagne MM (1999) Apomixis for crop improvement. Protoplasma 208:196–205
Song R, Llaca V, Messing J (2002) Mosaic organization of orthologous sequences in grass genomes. Genome Res 12:1459–1555
Tarchini R, Biddle P, Wineland R, Tingey S, Rafalski A (2000) The complete sequence of 340 kb of DNA around the rice Adh1-Adh2 region reveals interrupted colinearity with maize chromosome 4. Plant Cell 12:381–391
Tikhonov AP, SanMiguel PJ, Nakajima Y, Gorenstein NM, Bennetzen JF, Avramova Z (1999) Colinearity and its exceptions in orthologous adh regions of maize and sorghum. Proc Natl Acad Sci USA 96:7409–7414
Van Deynze AE, Sorrels ME, Park WD, Ayres NM, Fu H, Cartinhour SW, Paul E, McCouch SR (1998) Anchor probes for comparative mapping of grass genera. Theor Appl Genet 97:356–369
Vielle-Calzada JP, Crane CF, Stelly DM (1996) Apomixis: the asexual revolution. Science 274:1322–1323
Yu J, et al (2002) A draft sequence of the rice genome ( Oryza sativa L. ssp. indica). Science 296:79–92
Acknowledgements
This study was carried out within the framework of the Bilateral Agreement (2002–2003) MAE-SETCIP for Italian and Argentinian Research Institutions [Project No. 24(44B)]. The research was supported by the EC project Apotool (Contract No. GLG-CT-2000-0063). We thank Dr. T. Sasaki, the Japanese Rice Genome Research Program of the National Institute of Agrobiological Resources (NIAR), and the Institute of the Society of Techno-Innovation in Agriculture, Forestry and Fisheries (STAFF), Kannondai, Tsukuba, Japan for providing the rice clones, and Dr. S. McCouch of Cornell University (Ithaca, N.Y.) for the oat probe. The work was carried out in compliance with the current laws governing genetic experimentation in Italy and Argentina. This is Contribution No. 31 from the Institute of Plant Genetics-CNR, Research Division of Perugia
Author information
Authors and Affiliations
Corresponding author
Additional information
Communicated by R. Hagemann
Rights and permissions
About this article
Cite this article
Pupilli, F., Martinez, E.J., Busti, A. et al. Comparative mapping reveals partial conservation of synteny at the apomixis locus in Paspalum spp. . Mol Genet Genomics 270, 539–548 (2004). https://doi.org/10.1007/s00438-003-0949-5
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00438-003-0949-5