
Abstract
We recently engaged in a two-part study of Clinostomum Leidy, 1856 across a geographic range comprising central Mexico southwards to Costa Rica, in Central America. In the first study, we investigated the species boundaries by using DNA sequences of mitochondrial and nuclear molecular markers, implementing several analytical tools and species delimitation methods. The result of that approach revealed five highly divergent genetic lineages that were interpreted as independent evolutionary units, or species. Here, we present the second part of the study, where we describe three of the five species for which we have sexually mature adult specimens obtained from the mouth cavity of fish-eating birds. Additionally, we characterise morphologically the metacercariae of the other two species, collected from freshwater fishes; these species cannot be formally described since no adults were found in their definitive hosts. We further discuss the characters that are more reliable for species identification within Clinostomum, such as the cirrus sac shape and relative position with respect to testes and ovary, the shape of the reproductive organs, and the diverticulated condition of the caeca.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Clinostomum Leidy, 1856 is one of the genera allocated in the family Clinostomidae Lühe, 1901 along with other six genera according to Kanev et al. (2002), i.e. Clinostomoides Dollfus, Clinostomatopsis Dollfus, 1932, Euclinostomun Travassos, 1928, Odhneriotrema Travassos, 1928 and Nephrocephalus Odhner, 1902, and Ithyoclinostomum Witenberg, 1925. Clinostomum is a cosmopolitan genus comprising 15 well-recognised species, and at least another 14 species that have not yet been described (Locke et al. 2015; Pérez-Ponce de León et al. 2016; Caffara et al. 2017). Sexually mature adults are found in the mouth cavity and oesophagus of birds belonging at least to 12 families, although most records have been made in herons (Ardeidae) distributed worldwide (Matthews and Cribb 1998; Dzikowski et al. 2004; Gustinelli et al. 2010; Caffara et al. 2011, 2014, 2017; Sereno-Uribe et al. 2013; Pinto et al. 2015; Acosta et al. 2016; Locke et al. 2015; Pérez-Ponce de León et al. 2016). In a recent study, Pérez-Ponce de León et al. (2016) uncovered unexpected high levels of genetic diversity in Clinostomum across a range spanning between Mexico and Costa Rica. Five putative species were validated after analysing this genetic diversity following a species delimitation approach. Patterns of host association and biogeography provided additional support for the species hypotheses, although a detailed taxonomic analysis was not performed to provide the proper species description of these molecularly recognised species. The present study achieves the next step in our research. Here, we properly describe three of these species for which we sampled adult specimens from their definitive hosts; additionally, the metacercariae of two species are morphologically characterised, and we take a closer look at the diagnostic morphological characters used to delimit species in sexually mature specimens, discussing those that are more reliable for species identification of this taxonomically complex group of trematodes.
Materials and methods
Samples collected
Between February 2013 and February 2015, sexually mature adults of three lineages of Clinostomum were collected from 54 individuals of fish-eating birds representing four species in five locations of Mexico (Table 1). In addition, the metacercariae of five lineages of Clinostomum were collected in 156 fish individuals representing 18 species; fish were sampled in 20 locations in Mexico, one locality in Honduras, and six localities in Costa Rica (Table 1). Birds were captured with a shotgun, immediately kept on ice and examined 2 h after capture. Fish were captured with seine nets and electrofishing, kept alive and transported to the laboratory, sacrificed with an overdose of anaesthetic (sodium pentobarbital), and immediately examined. For birds, the buccal cavity and oesophagus were examined for adult specimens whilst for fish, all internal organs were placed in Petri dishes with 0.6% saline and examined for parasites under a stereomicroscope.
Morphological study
For morphological study, collected trematodes were fixed by sudden immersion in hot (nearly boiling) 4% formalin. Specimens were stained with Mayer’s paracarmine, Harris haematoxylin or Gomori’s trichrome, dehydrated in a graded ethanol series, cleared in methyl salicylate, and mounted as permanent slides with Canada balsam. Drawings were made with a drawing tube attached to the light microscope. Measurements are presented in micrometres (μm) unless otherwise stated, with the range followed by the mean in parentheses (Tables 2 and 3). Specimens were deposited in the Colección Nacional de Helmintos (CNHE), Instituto de Biología, Universidad Nacional Autónoma de México (UNAM). For morphological comparison, type and voucher specimens of the following species deposited in the CNHE were studied: Cinostomum tataxumui Sereno-Uribe, Pinacho-Pinacho, García-Varela, and Pérez-Ponce de León, 2013, ex Ardea alba L. (holotype CNHE 8337); ex Ardea herodias L. (paratype CNHE 8341); ex Tigrisoma mexicanum Swainson (paratype CNHE 8343); ex Dormitator maculatus Bloch (metacercariae CNHE 8339–8340); ex Dormitator latifrons Richardson (metacercariae CNHE 8344); Clinostomum marginatum (Rudolphi, 1819), ex A. alba (CNHE 8345-8346), ex Etheostoma pottsi Girard (CNHE 6316); ex Lepomis macrochirus Rafinesque (CNHE 6314), ex Micropterus salmoides Lacépède (CNHE 6325); Clinostomum heluans Braun, 1899, ex A. herodias (CNHE 22-16); ex A. alba (CNHE 10665).
Morphometrical analyses
A multivariate analysis of variance (MANOVA) was performed considering 40 measurements of morphological traits taken from adults of the three new species described in this study, and also 30 measurements taken from metacercariae of the five genetic lineages. In total, 23 adult specimens were measured (9 of lineage 2, 6 of lineage 4, and 8 of lineage 5) and 47 metacercariae (5 of lineage 1, 7 of lineage 2, 17 of lineage 3, 10 of lineage 4, and 8 of lineage 5). The analysis of variance was conducted using Scheffé’s F procedure for post hoc comparisons, because it is robust to violations of assumptions inherent in multiple comparison procedures (e.g. the assumption of homogeneity of variances). Additionally, a principal component analysis (PCA) was implemented to explore and describe patterns of variation of our data. The MANOVA and PCA were performed using the software STATISTICA version 7.1 (StatSoft Inc 2005).
Results
The molecular characterisation of adults and metacercariae of Middle American Clinostomum within a species delimitation framework allowed us to validate five new species. For three of them, a formal taxonomic description is presented since adults were recovered from their definitive hosts. For the other two species, metacercariae are morphologically characterised awaiting the collection of adults from their definitive hosts.
Family Clinostomatidae Lühe, 1901
Genus Clinostomum Leidy, 1856
Clinostomum caffarae n. sp.
Description (based on the holotype and eight paratypes; measurements taken from stained and mounted adult worms, Table 2) (Fig. 1a).

Clinostomum. caffarae n. sp. a Adult (Holotype) from A. alba, Lago de Catemaco, Veracruz, ventral view. b Metacercariae (paratype) from Astyanax aeneus, Santa María del Oro, Jalisco, ventral view. Scale bars: 1 mm
Body elongated, wider in ventral sucker region. Tegument lacking spines. Oral sucker small, surrounded by prominent oral collar. Pharynx small, opening into oesophageal bulb. Ventral sucker in anterior third of body, round, larger than oral sucker. Caeca smooth, with small lateral diverticules in testicular region and extending to post-testicular region. Testes in tandem, entire to slightly irregular. Anterior testis post-equatorial, slightly displaced to left. Posterior testis in posterior part of middle third of body. Cirrus sac oval, dextral, overlapping right margin of anterior testis and right caecum; contains large seminal vesicle, narrow ejaculatory duct, and inconspicuous cirrus. Genital pore at level of anterior right margin of anterior testis. Ovary relatively large, ovoid, larger than wide, intertesticular, on right side of body. Vitellaria entirely extracaecal; vitelline follicles distributed in two fields alongside the body, extending from the posterior margin of ventral sucker to posterior end of body. Uterine sac tubular, intracaecal; ascending loop from intertesticular region, passing around left margin of anterior testis and extending to the mid-level between ventral sucker and anterior testis; descending uterine loop running straight to genital pore. Eggs yellowish, elliptical, often located within uterine sac, uterine duct, and ootype region. Metraterm muscular, poorly developed. Excretory vesicle V-shaped. Excretory pore terminal.
Synonym: Clinostomum sp. Lineage 2 after Pérez-Ponce de León et al. (2016).
Type-host: Ardea alba L.
Other hosts: Egretta thula (Molina).
Type-locality: Lago de Catemaco, Veracruz, Mexico.
Type-material: CNHE 10778 (holotype), 10779 (paratypes).
Etymology: The epithet caffarae is for Professor Mónica Caffara in recognition of her contribution to understand the morphological and genetic diversity of the trematode genus Clinostomum.
Representative DNA sequences: KU156797–99, KU156803 (cox1), KU156758, KU156760–61 (ITS).
Metacercaria.
Description (based on measurements of seven specimens; measurements in Table 3) (Figs. 1b and 5d–f).
Body shape similar to adults. Tegument lacking spines. Oral collar well developed. Oral sucker smaller than ventral sucker. Pharynx small, opening into oesophageal bulb. Ventral sucker large, round. Caeca with diverticules from ventral sucker level to posterior end of body. Testes smaller that in adult. Anterior testis with smooth borders, irregularly shaped; posterior testis kidney-shaped, with smooth borders. Cirrus sac ovoid, extending along the right border of anterior testis, overlapping right caecum. Genital pore al level of anterior border of anterior testis. Ovary ovoid, intertesticular, on right side of body. Vitellaria undeveloped. Excretory vesicle V-shaped.
Hosts: Astyanax aeneus Günther.
Localities: México: Santa María del Oro and Río Las Rosas, Jalisco.
Specimen deposition: CNHE (10781–10782).
Representative DNA sequences: KU156786–91 (cox1), KU156747–52 (ITS).
Clinostomum arquus n. sp.
Description (based on the holotype and five paratypes; measurements taken from stained and mounted gravid adult worms, see Table 2) (Fig. 2a).

Clinostomum arquus n. sp. a Adult (Holotype) from E. thula, Lago de Catemaco, Veracruz, ventral view. b Metacercariae (paratype) from Profundulus punctatus, Río Chacalapa, Oaxaca, ventral view. Scale bars: 1 mm
Body elongated, wider in gonadal region. Tegument lacking spines. Oral sucker small, surrounded by prominent oral collar. Pharynx small, opening into pharyngeal bulb. Ventral sucker in anterior third of body, round, larger than oral sucker. Caeca smooth, with small lateral diverticules from level of posterior margin of ventral sucker to posterior end of body. Testes in tandem, entire, post-equatorial. Anterior testis wider than larger. Posterior testis in posterior third of body, almost round. Cirrus sac curved, dextral, overlapping right margin of anterior testis; contains large seminal vesicle, narrow ejaculatory duct, and inconspicuous cirrus. Genital pore at level of anterior right margin of anterior testis. Ovary small, ovoid to round, intertesticular, on right side of body. Vitellaria entirely extracaecal; vitelline follicles distributed in two fields alongside the body, extending from level of posterior margin of ventral sucker to posterior end of body, not reaching end of caeca. Uterine sac tubular, intracaecal; ascending loop from ootype area at intertesticular region, passing around left margin of anterior testis and extending to the mid-level between ventral sucker and anterior testis; descending uterine loop running straight to genital pore. Eggs yellowish, elliptical, often located within uterine sac, uterine duct, and ootype region. Metraterm not observed. Excretory vesicle V-shaped. Excretory pore terminal.
Synonyms: Clinostomum sp. Lineage 4, after Pérez-Ponce de León et al. (2016); and Clinostomum sp. 3 after Locke et al. (2015).
Type-host: Egretta thula Molina.
Type-locality: Lago de Catemaco, Veracruz, Mexico.
Type-material: CNHE 10783 (holotype), 10784 (paratypes).
Etymology: The epithet arquus is derived from the Latin arquus, meaning anything arched or curved, referring to the cirrus sac shape. It stands as a noun in apposition.
Representative DNA sequences: KU156800–03 (cox1), KU156759 (ITS).
Metacercaria.
Description (based on measurements of eight specimens; measurements in Table 3) (Figs. 2b and 5j–l).
Body shape similar to adults. Tegument lacking spines. Oral collar well developed. Oral sucker smaller than ventral sucker. Pharynx small, opening into oesophageal bulb. Ventral sucker large, round. Caeca with diverticules from ventral sucker level to posterior end of body. Testes and ovary smaller that in adult. Anterior testis entire, irregularly shaped; posterior testis oval. Cirrus sac curved, extending along the right border of anterior testis, slightly overlapping right caecum. Genital pore al level of anterior border of anterior testis. Ovary ovoid, intertesticular, on right side of body. Vitellaria and uterine sac undeveloped. Excretory vesicle V-shaped.
Hosts: Profundulus punctatus (Günther); P. balsanus Ahl; Pseudoxiphophorus jonesii (Günther).
Localities: Mexico: Río Chacalapa, Presa Los Ocotes, El Platanar, Río Santa María Huatulco, Oaxaca; Río El Triunfo, Chiapas.
Specimen deposition: CNHE (10785–10786).
Representative DNA sequences: KU156779–81, KU156792–93, KJ477559–69, KJ477581, KJ504184–91 (cox1), KU156753, KM676409–11, KJ477581, KJ477587–96, KJ477663–64, KJ477676–77 (ITS).
Clinostomum cichlidorum n. sp.
Description (based on the holotype and seven paratypes; measurements taken from stained and mounted gravid adult worms, see Table 2) (Fig. 3a).

Clinostomum cichlidorumn. n. sp. a Adult (Holotype) from T. mexicanum, Río Champotón, Campeche, ventral view. b Metacercariae (paratype) from Amatitlania siquia, Río las Vueltas, Costa Rica, ventral view a. Scale bars: 1 mm
Body elongated, wider in gonadal region. Light constriction at ventral sucker level. Tegument lacking spines. Oral sucker small, surrounded by prominent oral collar. Pharynx small, opening into pharyngeal bulb. Ventral sucker in anterior third of body, round, larger than oral sucker. Caeca diverticulated along the entire extension, from cecal bifurcation to posterior end of body. Testes in tandem, deeply lobated, post-equatorial; short distance between testes. Anterior testis wider than long, short distance from posterior margin of ventral sucker. Posterior testis in posterior third of body. Cirrus sac straight, dextral, overlapping right margin of anterior testis and ovary; contains convoluted seminal vesicle and ejaculatory duct; cirrus not observed. Genital pore at mid-level of anterior testis. Ovary small, round, intertesticular, on right side of body. Vitellaria extracaecal in pre-testicular area, confluent in post-testicular space; vitelline follicles extending from level of posterior margin of ventral sucker to posterior end of body, not reaching end of caeca. Uterine sac tubular, short, intracaecal; ascending loop from ootype area at intertesticular region, passing around left margin of anterior testis and extending to reach posterior border of posterior testis; descending uterine loop running straight to genital pore. Eggs yellowish, elliptical. Metraterm not observed. Excretory vesicle V-shaped. Excretory pore terminal.
Synonym: Clinostomum sp. Lineage 5 after Pérez-Ponce de León et al. (2016).
Type-host: Tigrisoma mexicanum Swainson.
Other hosts: Cochlearius cochlearius (L.); Ardea alba L.
Type-locality: Río Champotón, Campeche, Mexico.
Other localities: Laguna Silvituc, Campeche; El Espino, Tabasco; Río Verde, Oaxaca, Mexico.
Type-material: CNHE 10787 (holotype), 10788–90 (paratypes).
Etymology: The species is named after the host group, cichlids (family Cichlidae), for which the metacercariae exhibit a narrow host-specificity.
Representative DNA sequences: KU156794–96, KJ477506–10, KJ477536–45, KJ504197–201, KJ504203 (cox1), KU156754–57, KJ477619–22, KJ477645–53, KJ477667–68, KJ477670 (ITS).
Metacercaria.
Description (based on measurements of six specimens; measurements in Table 3) (Figs. 3b and 5m–o).
Body shape similar to adults. Tegument lacking spines. Oral collar well developed. Oral sucker smaller than ventral sucker. Pharynx small, opening into pharyngeal bulb. Ventral sucker large, round. Caeca with diverticules, more prominent from ventral sucker level to posterior end of body. Testes smaller that in adult, irregularly shaped. Anterior testis displaced to the right side of body; posterior testis wider than anterior testis. Cirrus sac ovoid, extending along the dextral border of anterior testis, overlapping right caecum and ovary. Genital pore at mid-level of anterior border of anterior testis. Ovary round, intertesticular, on right side of body. Vitellaria undeveloped.
Hosts: Cichlasoma urophthalmus (Günther), C. trimaculatum (Günther), Thorichthys meeki Brind, Herichthys cyanoguttatus Baird and Girard, Parachromis managuensis (Günther), Amatitlania nigrofasciata (Günther), Amatitlania siquia Schmitter-Soto, Amphilophus sp. (Cichlidae).
Localities: Mexico: Laguna Silvituc, Laguna Santa Cruz, Laguna El Milagro, Campeche; San Felipe, Río Atlapexco, Hidalgo; Laguna Los Milagros, Laguna Azul, Quintana Roo; Mitlapa, San Luis Potosí; Río Verde, Flores Magón, Oaxaca. Costa Rica: Río Las Vueltas, Río Irigaray, Horquetas de Sariquipí.
Specimen deposition: CNHE (10791–10793).
Representative DNA sequences: KU156784–85, KU156809–18, KJ477465–73, KJ477490–97, KJ477511–35, KJ477556–58, KJ504194–96, KJ504204–05 (cox1), KU156746, KU156770–73, KJ477578–86, KJ477608–11, KJ477623–44, KJ477660, KJ477665–66, KJ477671 (ITS).
Clinostomum sp. Lineage 1 sensu Pérez-Ponce de León et al. (2016).
(Metacercariae)
Description (based on measurements of five specimens; measurements in Table 3) (Figs. 4a and 5a–c).

Metacercariae of Clinostomum sp. Lineage 1 (a) from Rhamdia sp. El Paraíso, Honduras, ventral view. Metacercariae of Clinostomum sp. Lineage 3 (b) from Rhamdia guatemalensis, Lago de Catemaco, Veracruz, ventral view. Scale bars: 1 mm

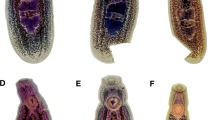
Scanning electron micrographs of Clinostomum metacercariae Lineages 1–5. a–e Whole body of metacercariae. Scale bars: 1 mm. f–j Anterior end showing the oral sucker collar of metacercariae. Scale bars: 250 μm. k–o Body surface at level of ventral sucker of metacercariae. Scale bars: 250 μm
Body oval, elongated, wider in gonadal region. Light constriction at ventral sucker level. Tegument possessing spines along the body. Oral collar well developed, smaller than ventral sucker. Pharynx not observed. Esophageal bulb inconspicuous. Ventral sucker in anterior third of body. Caeca run laterally to posterior end of body with diverticulated margins from posterior margin of ventral sucker to posterior end of body. Testes posterior to the mid-line of body, irregularly shaped, with irregular borders; intertesticular space short. Cirrus sac ovoid located dextrally to the border of anterior testis, overlapping ovary, opening directly into the genital pore at mid-level of anterior testis. Ovary small, rounded, located in intertesticular space on right side of body. Uterine sac tubular, poorly developed, extending anteriorly between posterior border of ventral sucker and anterior testis. Vitellaria undeveloped.
Hosts: Rhamdia sp.
Other hosts: Rhamdia guatemalensis (Günther).
Localities: El Paraiso, Honduras.
Other localities: Mexico: Río San Juan, Chiapas.
Specimens deposited: CNHE (10794).
Representative DNA sequences: KU156782–83, KU156804, KU156806 (cox1), KU156743–45, KU156763, KU156766 (ITS).
Clinostomum sp. Lineage 3 sensu Pérez-Ponce de León et al. (2016).
(Metacercariae)
Description (based on measurements of 17 specimens; measurements in Table 3) (Figs. 4b and 5g–i).
Body oval, elongated, wider in gonadal region. Light constriction at ventral sucker level. Tegument lacking spines. Oral collar well developed, smaller than ventral sucker. Pharynx not observed. Esophageal bulb inconspicuous. Ventral sucker in anterior third of body. Caeca run laterally to posterior end of body, with diverticulated margins from posterior margin of ventral sucker to posterior end of body. Testes posterior to the mid-line of body, irregularly shaped, with irregular borders; intertesticular space short. Cirrus sac ovoid, slightly curved, dextrally located between right margin of anterior testis and caecum. Genital pore anterior to anterior testis. Ovary small, ovoid, located in intertesticular space on right side of body. Uterine sac not observed. Vitellaria undeveloped.
Hosts: Rhamdia guatemalensis (Günther).
Other hosts: Rhamdia sp.; Rhamdia rogersi (Regan); Gobiomorus maculatus (Günther).
Locality: Lago de Catemaco, Veracruz, Mexico.
Other localities: Mexico: Laguna El Milagro, Campeche; Emiliano Zapata, Tabasco; Honduras: El Paraiso; Costa Rica: Tilarán, Río Las Vueltas, Río Grande, Quebrada Ganado.
Specimens deposited: CNHE (10795–10797).
Representative DNA sequences: KU156807, KU156808, KU156819–20, KJ477443–50, KJ477474–89, KJ477502–05, KJ477546–47 (cox1), KU156767–69, KU156774–75, KJ477571–77, KJ477597–607, KJ477615–18, KJ477654–55 (ITS).
Taxonomic remarks
The three new species we describe herein can be differentiated by a combination of morphological characters. Clinostomum caffarae n. sp. is characterised by lacking spines on the tegument, by having entire to slightly irregular testes, a cirrus sac overlapping the right margin of anterior testis and right caecum, a genital pore at the level of anterior right margin of anterior testis and vitelline follicles in two fields alongside the body entirely extracaecal. The metacercariae of this species are exclusively found in the Banded tetra, Astyanax aeneus in western Mexico. Clinostomum arquus n. sp. lacks spines on the tegument and possesses entire and almost round testes and a characteristic curved cirrus sac overlapping the right margin of the anterior testis; the metacercariae of this species are found in killifishes of the genus Profundulus Hubbs across the states of Oaxaca and Chiapas in Mexico. Finally, Clinostomum cichlidorum n. sp. is characterised by lacking spines on the tegument, and by having diverticulated caeca along their entire extension, deeply lobated testes, a straight cirrus sac overlapping right margin of anterior testis and ovary and vitelline follicles extracaecal in the pre-testicular area, but confluent in the post-testicular space; additionally, the distance between testes, and the distance between the posterior border of ventral sucker and the anterior testis in the new species are short; the metacercariae of C. cichlidorum n. sp. are exclusively found in several species of cichlids across a wide geographical range, from southern Mexico southwards to Costa Rica. Multivariate analyses of morphometric traits through MANOVA and PCA were conducted to establish the differentiation amongst the adults and metacercariae described or characterised in this study. The MANOVA post hoc test showed that 25 of the 40 characters considered for adults were significantly different between lineages (P ˂ 0.05) in at least one pairwise group comparison. In adults, only two characters were significantly different between the three lineages, i.e. distance between testes (DBT) and distance between anterior testis and ventral sucker (VS-AT) (Fig. 6; Supplementary Table S1). For metacercariae, the test showed that 29 of the 35 characters were significantly different between lineages (P ˂ 0.05) in at least one pairwise group comparison. However, we did not find significantly different pairwise comparisons between the five metacercariae lineages (Fig. 6; Supplementary Table S2). The PCA analysis of adults (Fig. 6b) show that three clusters are formed, each corresponding with one of the three species; however, the analysis of metacercariae (Fig. 6c) shows that polygons of the five lineages overlap to some extent, with the exception of Clinostomum Lineage 1, although the small sample size of that lineage may influence the result of the analysis. The first two axes of the PCA for both, adults and metacercariae (Figs. 6b, c) accounted for 62.2 and 53.2% of the variance, respectively.

Multivariate analysis of variance (MANOVA) and principal component analysis (PCA) amongst species/genetic lineages of Middle American Clinostomum. Analyses consider 40 measurements of morphological traits. a Difference amongst the three species (Lineages 2, 4 and 5) based on VS-AT (distance between ventral sucker and anterior testis) and DBT (distance between testes) (b). Principal component analysis scatterplots of adults (c). Principal component analysis scatterplots of metacercariae
Of the three new species herein described, C. cichlidorum n. sp. is morphologically similar to C. tataxumui and C. marginatum, two additional species of Clinostomum that occur in the same geographical area in Middle America (Sereno-Uribe et al. 2013). The three species possess deeply lobated testes and vitelline follicles confluent in the post-testicular area; however, they can be easily differentiated because in the latter two, follicles are also confluent in the pre-testicular region. Additionally, the new species we describe herein has a cirrus sac located almost intertesticularly, overlapping the ovary, whereas in C. tataxumui, the cirrus sac is located next to the right border of anterior testis, displacing it to the left, and in C. marginatum, the cirrus sac is situated on the anterior border of anterior testis. Interestingly, the species also differ in host association between the metacercariae and their second intermediate host. Whilst the metacercariae of C. tataxumui are exclusively found in brackish water eleotrids, those of C. marginatum are found in several species of Nearctic freshwater fishes (see Sereno-Uribe et al. 2013; Pérez-Ponce de León et al. 2016). At the moment, the species differentiation between the two metacercariae we validated molecularly and the other congeneric species cannot be established, until adult forms are obtained from their definitive hosts.
Discussion
The three new species of Clinostomum herein described, and the two genetic lineages, were first recognised and delimited by using a molecular species delimitation approach (Pérez-Ponce de León et al. 2016). Clinostomum sp. Lineage 1 and 3 were recognised as independent species in the aforementioned analysis; however, both are described solely as metacercariae. For Lineage 1, no adults were found in the fish-eating birds we studied. In the case of Lineage 3, only four adults were collected, three from Tigrisoma mexicanum and one from Ardea herodias from Emiliano Zapata, Tabasco; however, all these specimens were used for sequencing work and no specimen was kept as hologenophore (sensu Pleijel et al. 2008) for morphological study. Likewise, our two-part study demonstrated the utility of DNA sequences to establish more robust species limits amongst digeneans. In particular, several studies had shown the utility of ITS and cox1 as molecular markers to discriminate amongst species of clinostomids (e.g. Caffara et al. 2011, 2017; Sereno-Uribe et al. 2013; Locke et al. 2015). In our study, five putative species were recognised and delimited through a molecular phylogenetic analysis that used both molecular markers, and a framework for species delimitation (Pérez-Ponce de León et al. 2016). This methodological approach was necessary because of the extensive number of metacercariae and adults that were collected from both intermediate and definitive hosts across a wide geographical range in Middle America, based on the premise that the genetic divergences within and between species are influenced by sampling effort (Locke et al. 2015; Caffara et al. 2017). Unfortunately, we only sampled enough adult specimens for three of the five genetic lineages to complete the second part of our study, and to properly describe these species. Pante et al. (2015a) discussed the disconnection between the work of taxonomists describing species as working hypotheses and the use of species delimitation approaches aimed at describing patterns of gene flow. These authors actually reinforced the view that molecular studies have shown a high prevalence of cryptic species, representing a species hypothesis, but usually not properly described and named; this is, however, a necessary step during the investigation of cryptic species (see Pérez-Ponce de León and Nadler 2010; Nadler and Pérez-Ponce de León 2011). Satler et al. (2013) further argued that turning DNA-based discoveries into taxonomic entities, i.e. species which are robustly and formally described and named, remains challenging. In fact, Pante et al. (2015b) provided evidence through a meta-analysis of the articles published between 2006 and 2013 that most new species are still described without the help of molecular data and that most of the new species are described without applying an integrative taxonomy approach. This is not the case, however, for clinostomid trematodes.
The genus Clinostomum currently contains 16 valid species, including (in chronological order): C. complanatum Rudolphi, 1814, C. marginatum Rudolphi, 1819, C. detruncatum Braun, 1899, C. heluans Braun, 1899, C. attenuatum Cort, 1913, C. hornum Nicoll, 1914, C. australiense Johnson, 1917, C. intermedialis Lamont, 1920, C. phalacrocoracis Dubois, 1930, C. kassimovi Vaidova and Feizullaev, 1958, C. philippinensis Velazquez, 1959, C. cutaneum Paperna, 1964, C. tilapiae Ukoli, 1966, C. wilsoni Matthews and Cribb, 1998, C. tataxumui Sereno-Uribe, Pinacho-Pinacho, García-Varela and Pérez-Ponce de León, 2013, and C. album Rosser, Alberson, Woodyard, Cunningham, Pote, and Griffin, 2017 (Locke et al. 2015; Pérez-Ponce de León et al. 2016; Rosser et al. 2017). Eight out of the 16 species have been validated using a combined morphological and molecular approach (Caffara et al. 2017; this study); the number of DNA sequences of Clinostomum species has accumulated very rapidly after studies conducted by different research groups around the world. At least another 12 species have been molecularly recognised in three recent studies, i.e. Locke et al. (2015), Pérez-Ponce de León et al. (2016), and Caffara et al. (2017). Three of these putative species are formally described in our study, and the metacercarial stage of the other two is also described, but not named until adult forms are collected in their definitive hosts. Therefore, after the present study, the number of validated species of Clinostomum through an integrative taxonomy approach rises to 11.
The description of new species of Clinostomum is complex because the morphological characters used traditionally to distinguish species can be considered of low utility, and not reliable. The genus Clinostomum has had a confusing taxonomic history because many of the species are very similar to each other, differing only in minor characters (Matthews and Cribb 1998). For instance, the validity and priority of two species, C. complanatum and C. marginatum, have been in dispute for nearly 200 year (see Dzikowski et al. 2004; Caffara et al. 2011). Actually, the study by Caffara and co-workers elegantly demonstrated the validity of C. marginatum by combining novel genetic data and morphological analyses of two life cycle stages, finding reliable morphological differences in the genital complex of both developmental stages; this difference was corroborated further by comparing ITS and cox1 DNA sequences of both species, leading the authors to conclude that only C. marginatum occurs in the Americas. However, the taxonomic confusion within the genus Clinostomum arises from inadequate species descriptions; some congeneric species were erroneously described from metacercariae rather than from adults, and one species was even described from a cercaria (see discussion in Matthews and Cribb 1998). Probably, the most important revisionary study of the genus Clinostomum was conducted by Ukoli (1966); this author synonymised 20 of the species described until 1966 with C. complanatum, and only considered 13 valid species. Baer (1933) constructed a classification key for species of Clinostomum using morphological characters such as the anterior extent of the vitellaria, arrangement of vitelline follicles, location of the genital complex, uterus shape, distance of ventral sucker from the anterior end of body, presence or absence of vitelline reservoir, body length, and diameter of oral sucker. After his revisionary work through the examination of the original descriptions, measurements, and illustrations of most congeneric species, Ukoli (1966) concluded that the most reliable characters for species separation were the anterior extent of the vitellaria, the presence of lateral evaginations in the uterus, the location of the genital complex along the antero-posterior axis of body, the testes shape, the position of the genital pore relative to the anterior testis, and the position of the cirrus sac. In his opinion, all other characters were likely subject to variations due to fixation method, state of contraction or expansion, and developmental stage of the specimens. Years later, Feizullaev and Mirzoeva (1983, 1986) examined hundreds of specimens of Clinostomum and concluded that only a single species, C. complanatum, should be recognised. This proposal apparently had a dramatic influence in the taxonomic literature because species identifiable as belonging to Clinostomum all over the world were thereafter referred routinely as C. complanatum, a proposal not currently accepted because, as pointed out by Matthews and Cribb (1998), this “view oversimplifies the complexity of the genus”. The taxonomic revision of the latter authors allowed them to conclude that three species, i.e. C. australiense, C. hornum, and C. wilsoni, are valid and all restricted to Australia, and that C. complanatum occurs in Australia because it is a cosmopolitan species.
Matthews and Cribb (1998) also suggested an approach to subject a small group of putative species of Clinostomum from a geographic area to a molecular analysis to establish species limits, and to investigate the morphological characters that could be more reliable for species delineation. The fundamental principle of our studies on Clinostomum of the last few years follows this idea (see Pérez-Ponce de León et al. 2016). The validation of the congeneric species occurring in Australian birds conducted by Matthews and Cribb (1998) considered morphological characters such as body size and shape, the oral sucker/ventral sucker ratio, size of the oral collar in relation to the body, presence or absence of diverticula in caeca, anterior testis shape and size, cirrus sac size, position of the genital pore, and ovary shape and size. The most recent species descriptions in the genus have used a combination of some of the characters endorsed by Ukoli (1966) and Matthews and Cribb (1998). For instance, C. tataxumui was separated from other congeneric species from the same geographic area by the body size, the position of the genital complex along anterior-posterior axis of the body, and the position of the cirrus sac with respect to the anterior testis; although a few additional characters were added, such as the distance between testes and the post-testicular space (Sereno-Uribe et al. 2013). Rosser et al. (2017) used the size of the body and some morphological features such as the size of oral and ventral suckers, testes and cirrus sac, and the distribution of vitelline follicles, to distinguish C. album from other species occurring in North America.
In the most recent assessment of Clinostomum species, Caffara et al. (2017) validated the metacercariae of C. tilapiae from African fish using morphological and molecular data. In addition, these authors studied metacercariae other than C. tilapiae and recognised four further putative species of Clinostomum from Africa, distinguished by a combination of morphological (structure of the genital complex) and molecular data. Morphological characterisation of the four morphotypes was based on the following features: position of the genital complex in relation to the antero-posterior axis of body, cirrus sac size and shape, and its position relative to the anterior testis, testes shape, the position of the ovary with respect to the cirrus sac, caeca shape (diverticulated or not), and presence or absence of minute spines on the tegument. Overall, considering the assessments by other authors, and our own criteria to establish a morphological differentiation between species that have been validated molecularly, like the species we recognised across a geographic area comprising Middle America, the morphological characters that seem to be most reliable for species delineation within Clinostomum are related with the genital complex (see Fig. 7). Only in one case, the body size and relative distance between anatomic structures can be used as a reliable character to discriminate amongst Clinostomum species; C. heluans is clearly differentiated from all congeneric species because is the longest member in the genus and gonads are restricted to the posterior end of body (Briosio-Aguilar et al. 2018); also, to discriminate between C. marginatum and C. complanatum, the body width was used, along with distance between ventral and oral sucker (Caffara et al. 2011). However, in our opinion, in the case of species of Clinostomum, no single character can be taken to discriminate amongst species, and a combination of characters has to be used routinely to establish more robust species limits, and preferably in combination with molecular data. Still, the distinction amongst morphologically similar species not necessarily closely related, like the ones we describe in this study, relies on the structure of the genital complex. This includes the position of the gonads along the body, whether they are in the middle or last third of body, the testes shape, whether they are smooth, irregular, or lobated, the position of the cirrus sac with respect to the testes, either displacing the anterior testes to the left, or occupying an intertesticular position, the position of the genital pore relative to the anterior testis borders.

Schematic representation of the genital complex of a Clinostomum marginatum, b C. tataxumui, c C. caffarae n. sp., d C. arquus n. sp., and e C. cichlidorum n. sp.
References
Acosta AA, Caffara M, Fioravanti ML, Utsunomia R, Zago AC, Franceschini L, Josè da Silva R (2016) Morphological and molecular characterization of Clinostomum detruncatum (Trematoda: Clinostomidae) metacercaria infecting Synbranchus marmoratus. J Parasitol 102:151–156. https://doi.org/10.1645/15-773.
Baer JG (1933) Note sur un nouveau trématode, Clinostomum lophophallum sp. nov., avec quelques considérations générales sur la famille des Clinostomidae. Rev Suisse Zool 40:317–342
Briosio-Aguilar R, Pinto HA, Rodríguez-Santiago MA, López-García K, García-Varela M, Pérez-Ponce de León G (2018) Link between the adult and the metacercaria of Clinostomum heluans Braun, 1899 (Trematoda: Clinostomidae) through DNA sequences, and its phylogenetic position within the genus Clinostomum Leidy, 1856. J Parasitol. https://doi.org/10.1645/17-183
Caffara M, Locke SA, Gustinelli A, Marcogliese DJ, Fiovaranti ML (2011) Morphological and molecular differentiation of Clinostomum complanatum and Clinostomum marginatum (Digenea: Clinostomidae) metacercariae and adults. J Parasitol 97:884–891. https://doi.org/10.1645/GE-2781.1
Caffara M, Davidovich N, Falk R, Smirnov M, Ofek T, Cummings D, Gustinelli A, Fioravanti ML (2014) Redescription of Clinostomum phalacrocoracis metacercariae (Digenea: Clinostomidae) in cichlids from Lake Kinneret, Israel. Parasite 21:32. https://doi.org/10.1051/parasite/2014034
Caffara M, Locke SA, Echi PC, Halajian A, Benini D, Luus-Powell WJ, Tavakol S, Fioravanti ML (2017) A morphological and molecular study of Clinostomid metacercariae from African fish with a redescription of Clinostomum tilapiae. Parasitology 144:1519–1529. https://doi.org/10.1017/S0031182017001068
Dzikowski R, Levy MG, Poore MF, Flowers JR, Paperna I (2004) Clinostomum complanatum and Clinostomum marginatum (Rudolphi, 1819) (Digenea: Clinostomidae) are separate species based on differences in ribosomal DNA. J Parasitol 90:413–414. https://doi.org/10.1645/GE-159R
Feizullaev NA, Mirzoeva SS (1983) Revision of the Superfamily Clinostomoidea and analysis of its system. Parazitologiya 17:3–11 (In Russian)
Feizullaev NA, Mirzoeva SS (1986) Keys to taxa of the Super-family Clinostomoidea (Trematoda). Parazitologiya 20:195–201 (In Russian)
Gustinelli A, Caffara M, Florio D, Otachi EO, Wathuta EM, Fiovaranti ML (2010) First description of the adult stage of Clinostomum cutaneum Paperna, 1964 (Digenea: Clinostomidae) from grey herons Ardea cinerea L. and a redescription of the metacercaria from the Nile tilapia Oreochromis niloticus niloticus (L.) in Kenya. Syst Parasitol 76:39–51. https://doi.org/10.1007/s11230-010-9231-5
Kanev I, Radev V, Fried B (2002) Family Clinostomidae Lühe, 1901. In: Gibson DI, Jones A, Bray RA (eds) Keys to the Trematoda, vol I. CAB International, Wallingford, pp 113–120
Locke SA, Caffara M, Marcogliese DJ, Fioravanti ML (2015) A large-scale molecular survey of Clinostomum (Digenea, Clinostomidae). Zool Scr 44:203–217. https://doi.org/10.1111/zsc.12096
Matthews D, Cribb TH (1998) Digenetic trematodes of the genus Clinostomum Leidy, 1856 (Digenea: Clinostomidae) from birds of Queensland, Australia, including C. wilsoni n. sp. from Egretta intermedia. Syst Parasitol 39:199–208
Nadler SA, Pérez-Ponce de León G (2011) Integrating molecular and morphological approaches for characterizing parasite cryptic species: implications for parasitology. Parasitology 138:1688–1709. https://doi.org/10.1017/S003118201000168X
Pante E, Schoelinck C, Puillandre N, Viricel A, Arnaud-Haond S, Aurelle D, Castelin M, Chenul A, Destombe C, Forcioli D, Valeron M, Viard E, Saadi S (2015a) Species are hypotheses: avoid connectivity assessments based on pillars of sand. Mol Ecol 24:525–544. https://doi.org/10.1111/mec.1304
Pante E, Schoelinck C, Puillandre N (2015b) From integrative taxonomy to species description: one step beyond. Syst Biol 64:152–160. https://doi.org/10.1093/sysbio/syu083
Pérez-Ponce de León G, Nadler SA (2010) What we don’t recognize can hurt us: a plea for awareness about cryptic species. J Parasitol 96:453–464. https://doi.org/10.1645/GE-2260.1
Pérez-Ponce de León G, García-Varela M, Pinacho-Pinacho CD, Sereno-Uribe AL, Poulin R (2016) Species delimitation in trematodes using DNA sequences: Middle-American Clinostomum as a case study. Parasitology 143:1773–1789. https://doi.org/10.1017/S0031182016001517
Pinto HA, Caffara M, Fioravanti ML, Melo AL (2015) Experimental and molecular study of cercariae of Clinostomum sp. (Trematoda: Clinostomidae) from Biomphalaria spp. (Mollusca: Planorbidae) in Brazil. J Parasitol 101:108–113. https://doi.org/10.1645/14-497.1
Pleijel F, Jondelius U, Norlinder E, Nygren A, Oxelman B, Schander C, Thollesson M (2008) Phylogenies without roots? A plea for the use of vouchers in molecular phylogenetic studies. Mol Phylogenet Evol 48:369–371. https://doi.org/10.1016/j.ympev.2008.03.024
Rosser TG, Alberson NR, Woodyard ET, Cunningham FL, Pote LM, Griffin MJ (2017) Clinostomum album n. sp. and Clinostomum marginatum (Rudolphi, 1819), parasites of the great egret Ardea alba L. from Mississippi, USA. Syst Parasitol 94:35–49. https://doi.org/10.1007/s11230-016-9686-0
Satler JD, Carstens BC, Hedin M (2013) Multilocus species delimitation in a complex of morphologically conserved Trapdoor Spiders (Mygalomorphae, Antrodiaetidae, Aliatypus). Syst Biol 62:805–823. https://doi.org/10.1093/sysbio/syt041
Sereno-Uribe AL, Pinacho-Pinacho CD, García-Varela M, Pérez-Ponce de León G (2013) Using mitochondrial and ribosomal DNA sequences to test the taxonomic validity of Clinostomum complanatum Rudolphi, 1814 in fish-eating birds and freshwater fishes in Mexico, with the description of a new species. Parasitol Res 112:2855–2870. https://doi.org/10.1007/s00436-013-3457-5
StatSoft Inc. (2005) STATISTICA (data analysis software system), version 7.1. www.statsoft.com
Ukoli FMA (1966) On Clinostomum tilapiae n. sp., and C. phalacrocoracis Dubois, 1931 from Ghana, and a discussion of the systematics of the genus Clinostomum Leidy, 1856. J Helminthol 40:187–214
Acknowledgments
We thank Robert Poulin for the review of an earlier version of the manuscript. We also thank Leopoldo Andrade, Eduardo Hernández, and Jesus Hernández for their help during field work in Mexico and Costa Rica; Luis García for providing specimens for comparison deposited at the Colección Nacional de Helmintos; and Berenit Mendoza for her help processing the samples for the SEM study. Hosts in Mexico were collected under the Cartilla Nacional de Colector Científico (FAUT 0202 and 0057) issued by the Secretaría del Medio Ambiente y Recursos Naturales (SEMARNAT), to MGV and GPPL, respectively. We thank Arturo Angulo for his help to collect fish specimens in Costa Rica under the collecting permit issued by the Costa Rican government.
Funding
This research was supported by grants from the Programa de Apoyo a Proyectos de Investigación e Inovación Tecnológica (PAPIIT-UNAM) I202617 and IN206716 to MGV and GPPL, respectively.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article

Cite this article
Sereno-Uribe, A.L., García-Varela, M., Pinacho-Pinacho, C.D. et al. Three new species of Clinostomum Leidy, 1856 (Trematoda) from Middle American fish-eating birds. Parasitol Res 117, 2171–2185 (2018). https://doi.org/10.1007/s00436-018-5905-8
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00436-018-5905-8