
Abstract
Trematodes of the genus Aphalloides Dollfus, Chabaud & Golvan, 1957 reach maturity in their second intermediate host, small fishes of the family Gobionellidae, genera Pomatoschistus and Knipowitschia. Two morphologically similar species have been described from European waters: Aphalloides coelomicola Dollfus et al., 1957 in the Mediterranean Sea (including the brackish Black Sea region) and Aphalloides timmi Reimer, 1970 in the Baltic Sea. There was no difference in morphology and morphometry for specimens corresponding to A. coelomicola and A. timmi and examination of the 28S rDNA sequence confirmed the similarity. Based on these results, these two species are synonymized and A. coelomicola redescribed.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
The Cryptogonimidae Ward, 1917 is a digenean family, which parasitize a large variety of poikilothermic vertebrates, both in fresh and marine waters (Miller and Cribb 2008b). Trematodes of genus Aphalloides Dollfus, Chabaud & Golvan, 1957 reach maturity in their second intermediate host such as small fishes of Gobionellidae family (see Thacker 2009), genera Pomatoschistus and Knipowitschia (Dollfus et al. 1957; Naydenova 1970, 1974; Maillard 1973; Vaes 1978; Zander et al. 2000; Zander and Reimer 2002; Kvach and Winkler 2011; Stoyanov et al. 2015). These fishes are both definitive and second intermediate hosts of these worms, but the first intermediate host are mud-snails of Hydrobia group (Maillard 1973; Vaes 1978; Reimer 1970). The adults are located in the coelomic cavity of the host, causing the parasitic castration of the fish (Pampoulie et al. 1999, 2004; Sasal and Pampoulie 2000). The goby hosts are annual fishes (Bouchereau and Guelorget 1998; Mazzoldi et al. 2002), thereafter the parasite life cycle has been shown to require host death (Pampoulie et al. 2000). Apart from gobionellids, only one case of incidental infection of the two-spotted goby (Gobiusculus flavescens (Fabricius, 1779) (Actinopterygii: Gobiidae)) has been recorded in the Flensburg Fjord, Germany (Josten et al. 2009).
There are two morphologically very similar parasite species described from the annual gobies from European waters: Aphalloides coelomicola Dollfus et al., 1957 in the Mediterranean Sea (including brackish-water in the Black Sea region) and Aphalloides timmi Reimer, 1970 in the Baltic Sea (Dollfus et al. 1957; Reimer et al. 1996). A. coelomicola was originally described from the Mediterranean fishes, but also occurs in the Black Sea and Sea of Azov (Dollfus et al. 1957; Naydenova 1974). The status of specimens from the North Sea is unclear: in Dievengat, Belgium, it was recorded as A. coelomicola (Vaes 1978), but in Øra area of Skagerrak, Norway, it was mentioned as A. timmi (Bakke 1980).
The Black Sea and the Baltic Sea present some similarities in their water characteristics. For example, both of these water bodies are much desalinated compared to the other seas (average S = 18‰ in the Black Sea, but 6–8‰ in the Baltic Sea) and are both classified as brackish-water basins (Zenkevich 1963; Dethier 1992). The faunas of both seas have the great group of Boreo-Atlantic species, which are relicts of the ancient Tethys Sea (from Upper Miocene) (Zaitsev and Mamaev 1997). Differing from the Baltic Sea, the Black Sea fauna consists of the group of Ponto-Caspian relicts and Mediterranean species (Zaitsev and Mamaev 1997; Zaitsev 1998). The colonization of the Black Sea by Mediterranean species (a process of Mediterranization) was started about 7000 years ago and occurs until now (Miller 1965; Zaitsev 1998).
Because of morphological similarity of the representatives of Aphalloides genus, the aim of our work was to compare the individuals from different geographical regions (Baltic, Black and Mediterranean seas) to clarify the taxonomic status of the Black Sea samples.
Material and methods
The parasites were sampled in different localities in the Mediterranean Sea (St.-Nazaire and Salse-Leucate lagoons), Black Sea (Budaki Lagoon, Sukhyi Estuary) and Baltic Sea (Salzhaff and Unterwarnow) from different species of Gobionellidae: Pomatoschistus marmoratus (Risso, 1810) and Knipowitschia caucasica (Berg, 1916) from the Black Sea, Pomatoschistus microps (Krøyer, 1838) from the Mediterranean and Baltic seas (Table 1). From each infected fish individual, some of the parasites were fixed in pure 96% ethanol for molecular study, and the others were preserved in hot 4% formaldehyde for morphological study (Cribb and Bray 2010). The formaldehyde-preserved worms were then stained with iron acetic carmine, dehydrated in ethanol of increasing concentration and mounted in Canada balsam as permanent slides (Georgiev et al. 1986). Voucher specimens are deposited in the helminthological collection of the Institute of Parasitology of the Academy of Sciences of the Czech Republic, (České Budějovice, Czech Republic).
The DNA from a worm individual from each sample was extracted using the JetQuick kit (Genomed, Germany). The genomic region of 28S rDNA was amplified by KAPA2G Robust HotStart PCR Kit (Kapabiosystems, USA) using primers and annealing temperature detailed in Table 2. Sanger sequencing of PCR products was commercially performed at GATC Biotech (Germany), and sequences were edited and aligned using Geneious 9.0.5 (Kearse et al. 2012). Phylogenetic analysis was based on 28S sequences of 15 newly sequenced individuals. The data of 28S sequences from previous study of Stoyanov et al. (2015)—A. coelomicola, GenBank accession no. KJ162159—were used to compare with 28S sequences from our data.
As out-group, the samples of five other cryptogonimid species from own samples were sequenced (see Table 1). Moreover, we used as out-group the sequences of Brachycladium goliath (KR703279), Acanthostomum burminis (KC489791; Jayawardena et al. 2013), Siphoderina jactus (EU571263) and Siphoderina poulini (EU571267; Miller and Cribb 2008a) available online.
The balsam slides were studied under the light microscope Olympus BX50 equipped with phase contrast, differential interference contrast (DIC) and digital image analysis (Olympus MicroImage™ for Windows 95/98/NT 7.0 (Olympus Optical Co.). The length and width of body, oral and ventral suckers, pharynx, ovary, both testes, seminal receptacle, and egg also length of oesophagus were measured to the nearest micrometre.
Results
Molecular study
Bayesian inference analysis based on sequences of 28S rDNA showed strong support for a monophyletic group encompassing genera Timoniella and Aphalloides (posterior probability PP = 0.99) (Fig. 1). The genus Aphalloides was also monophyletic, included all samples from France, Germany and Ukraine, but the support was only PP = 0.89. Within the genus, only little variation was observed for the 28S rDNA, but it seems that specimens ex. Knipowitschia caucasica (samples from Atanasovsko Lake, Bulgaria, and Sukhyi Estuary, Ukraine) differ from samples ex. Pomatoschistus spp. (the latter form supported monophyletic group, PP = 0.99). Sequences from individuals identified previously as A. cf. timmi from Germany are identical to those identified as A. coelomicola from France.

Phylogenetic trees of Cryptogonimids based on sequences of 28S rDNA
Morphological study
The morphological study of worms from different region did not show differences (Table 3; Fig. 2). The morphometric parameters overlap between worms from different regions and with data from previous authors or original descriptions (see Table 3; Dollfus et al. 1957; Naydenova 1970; Reimer 1970). Based on our observations, we provide a redescription of A. coelomicola as follows:

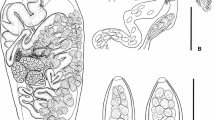
Aphalloides coelomicola. a ex. Pomatoschistus microps, St.-Nazaire Lagoon. b ex. P. microps, Salzhaff. c Photograph of a speciman ex. Knipowitschia caucasica with vitellin ducts visible. Scale bar 500 μm
Family Cryptogonimidae Ward, 1917.
Genus Aphalloides Dollfus, Chabaud & Golvan, 1957.
Aphalloides coelomicola Dollfus, Chabaud & Golvan, 1957 (Fig. 2).
Synonym: Aphalloides timmi Reimer, 1970
Type host: Pomatoschistus microps (Krøyer, 1838) (Actinopterygii, Gobionellidae).
Other hosts: Pomatoschistus minutus (Pallas, 1770), Pomatoschistus marmoratus (Risso, 1810), Knipowitschia caucasica (Berg, 1916), and Knipowitschia longicaudata (Berg, 1916) (Actinopterygii: Gobionellidae); Incidentally: Gobiusculus flavescens (Fabricius, 1779) (Actinopterygii: Gobiidae).
Type locality: Mediterranean Sea near Banyuls-sur-Mer, France.
Other localities: Mediterranean Sea (Camargue, Salse-Leucate Lagoon, St.-Nazaire Lagoon), Black Sea (Atanasovsko Lake, gulfs of Odessa and Yagorlyk, lagoons and estuaries of the North-Western Black Sea, Sea of Azov), Baltic Sea (Rügen Island, lagoons and bights of South-Western Baltic Sea) and North Sea (Dievengat, Øra area of Skagerrak) (see Fig. 3).

Confirmed findings of Aphalloides coelomicola in Europe. Mediterranean Sea: 1 Banyuls-sur-Mer (Dollfus et al. 1957), 2 Salse-Leucate and St.-Nazaire lagoons (this study), 3 Vaccarès and Malagroy lagoons, Camargue (Maillard 1973; Bayssade-Dufour and Maillard 1982; Pampoulie et al. 1999, 2000, 2004; Sasal and Pampoulie 2000); Black Sea: 4 Atanasovsko Lake (Stoyanov et al. 2015), 5 Budaki Lagoon (Kvach 2010; this study), 6 Sukhyi Estuary (Kvach 2010; Krasnovyd et al. 2012; this study), 7 Gulf of Odessa (Chernyshenko and Sventsytskaya 1970), 8 Hryhorivsky and Tyligul estuaries (Chernyshenko and Sventsytskaya 1970; Kvach 2010), 9 Berezan Estuary (Chernyshenko and Sventsytskaya 1970), 10 Gulf of Yagorlyk (Naydenova 1974; Parukhin et al. 1983); Sea of Azov (Naydenova 1974): 11 Gulf of Arabat, 12 Obytichna Split, 13 Biryuchy Island, 14 Taman, 15 Gulf of Yeysk; North Sea: 16 Dievengat Pond (Vaes 1978), 17 Øra area, Glomma River Estuary (Bakke 1980); Baltic Sea: 18 Flensburg Fjord (Josten et al. 2009), 19 Kiel Bight (Zander et al. 2002; Zander 2005), 20 Lübeck Bight (Zander 2003), 21 Salzhaff (Zander et al. 1999, 2000; Zander and Reimer 2002; this study), 22 Unterwarnow (Kvach and Winkler 2011; this study), 23 Rügen Island (Reimer 1970; Reimer et al. 1996)
Site in host: body cavity, gonads.
Voucher material: D-640 (Helminthological collection of the Institute of Parasitology of Academy of Sciences of the Czech Republic, České Budějovice, Czech Republic).
Representative sequence: KY978885, KY978886, KY978887, and KY978888.
Description:
Body large, elongate, variable in size: 350–3750 × 160–1516. Maximum body width in central part, in second quarter or at the level of ovary. Anterior part tapering, posterior part rounded. Tegument slightly rugose transversally, devoid of spines. Two brown eyespots at level of pharynx.
Oral sucker terminal, slightly elongate transversely, different in shape (from pyriform to globular): 37–122 × 37–130. Ventral sucker subspherical in shape, typically smaller than oral sucker, rarely slightly larger, 34–200 × 34–140. Positioned at level of intestinal bifurcation or slightly posterior, at base of tegumental depression.
Prepharynx absent. Pharynx muscular, thick-walled, elongate-oval, 27–95 × 31–59. Oesophagus different in length, thick-walled, 13–323, usually winding, rarely straight. Intestinal bifurcation in anterior quarter of body. Caeca elongate-saccular, without lateral diverticula, contiguous, winding, terminate blindly just anterior to anterior testis or slightly overlapping it. Sinistral caecum sometime slightly longer than dextral. Excretory pore terminal.
Genital pore located immediately anterior to ventral sucker. Gonotyle absent, cirrus and cirrus-sac absent. Excretory vesicle Y-shaped, reaches posterior margin of ventral sucker or slightly posterior to it. Distal part of seminal vesicle covered by prostatic cells. Seminal vesicle tubular, curved, positioned at middle of body.
Testes two, oval, located at same level, close to posterior extremity of body, one anterior to another, sometime slightly overlapping. Anterior testis 66–677 × 55–583, posterior 59–610 × 37–579. Ovary deeply tri-lobed, 46–346 × 59–365, anterior to testes. Ovary positioned sometime immediately at anterior margin of anterior testis, sometime distanced, almost equatorial. Follicular vitellarium in clusters of follicles arranged in groups, positioned from level of posterior end of ventral sucker to level of testes. Vitelline ducts can form ring surrounding seminal receptacle and/or anterior part of ovary. Uterine seminal receptacle anterior to ovary, oval to circular, usually 44–395 × 44–206, sometime gigantic, 500–680 × 282–600, can partially overlap ovary. Space between ventral sucker and testes usually overfill by uterus with eggs. Uterus tubular, strongly developed, forms numerous loops. Metraterm absent. Eggs small, elongate-oval, 24–35 × 12–17, yellowish-brown, not embryonated.
Discussion
In this work, based on both morphological and molecular data, we consider A. coelomicola and A. timmi as a single species in the monotypic genus Aphalloides. The species has a wide geographical range, which includes brackish waters of the Mediterranean region (including the Black Sea) and the Baltic Sea. The host range is limited by the group of annual gobies of Gobionellidae family. The genus was place in Siphoderinae Manter, 1934, a subfamily of Cryptogonimidae (Dollfus et al. 1957). Naydenova (1970) placed this genus to Heterophyidae, which was later disputed by Bayssade-Dufour and Maillard (1982). Miller and Cribb (2008b) do not recognize subfamilies in Cryptogonimidae, confirming Aphalloides inside this family.
The parasite was first described from the common goby (P. microps) from a small brackish-water canal in Banyuls-sur-Mer in Southern France, leading to the creation of a new genus and a new species (Dollfus et al. 1957). Later, this parasite was confirmed in the lagoons of the Rhone delta (Maillard 1973 and more recently Pampoulie et al. 1999). Maillard (1973) showed that the life cycle of A. coelomicola includes two hosts: a snail host (Hydrobia spp.) as first intermediate host with parthenogenetic stages, and the annual gobies, where metacercariae develop into adults. A progenetic life cycle has been characterized for this species, and the worms start to produce eggs at the metacercarial stage (see Lefebvre and Poulin 2005; Stoyanov et al. 2015).
In the Black Sea, the parasite was first recorded in the 1960s from the marbled goby (P. marmoratus) in the Gulf of Odessa and Tyligul Estuary (Chernyshenko and Sventsytskaya 1970). Naydenova (1970, 1974) found both metacercariae and adults in three goby species (P. marmoratus, Knipowitchia longicaudata and K. caucasica) from the northern Sea of Azov. Also, the adults occurred in Pomatoschistus minutus, P. marmoratus and K. caucasica from different parts of the North-Western Black Sea (Parukhin et al. 1983; Kvach 2010; Krasnovyd et al. 2012). Recently, it was recorded in K. caucasica from brackish Atanasovsko Lake in Bulgaria (Stoyanov et al. 2015).
In the Baltic Sea, the distribution of this species is limited to German coastal waters from the Flensburg Fjord (Josten et al. 2009) to Rügen Island (Reimer et al. 1996), where it is common in two gobiids (P. microps, P. minutus) in Kiel Bight (Zander et al. 2002; Zander 2005), Lübeck Bight (Zander 2003), Salzhaff (Zander et al. 1999) and Unterwarnow (Kvach and Winkler 2011).
Describing the new species A. timmi, Reimer (1970) reported differences relative to A. coelomicola as the ratio of suckers (the ventral sucker is larger than the oral sucker in A. timmi), the size of seminal receptaculum (smaller in A. timmi), and vitelline ducts surrounding the seminal receptacle (absent in A. coelomicola). Studying the specimens from the Øra area of Skagerrak, Norway, Bakke (1980) confirmed the absence of tegumental spines. Later, Miller and Cribb (2008b) mentioned this item as one of the characteristic of the genus Aphalloides.
Stoyanov et al. (2015) provided redescription of this species, based on specimens from K. caucasica. They confirmed the relation of Black Sea specimens to A. coelomicola, noting the smaller sizes of seminal receptacle in comparing to previously described (Dollfus et al. 1957; Naydenova 1970). Also, they noted the morphometrical variability in studied specimens. According to Stoyanov et al. (2015), only the presence of vitellin ducts surrounding the seminal receptacle is discriminating between the two species of Aphalloides. Our study confirmed the presence of these ducts, but they were observed not only in specimens from the Baltic Sea, but also in many specimens from France and Ukraine (see Fig. 2c).
Zander and Reimer (2002) considered this species (noted as A. timmi) to be a brackish-water/marine endemic of the Baltic Sea. But, the type-host of this parasite, the common goby (P. microps), is a Boreal-Atlantic fish, widely distributed from Norway in the north to the Canary Islands in the south, including the Mediterranean and Baltic seas (Miller 1986). Another host of this parasite, the sand goby (P. minutus), has similar range, also inhabiting the Black Sea region (Miller 1986). Taking into account the host specificity of Aphalloides, the presence of endemic parasites in widely distributed host seems unlikely.
In the Black Sea (including the Sea of Azov), A. coelomicola was considered as Mediterranean species (Naydenova 1974; Kvach 2010). The marbled goby (P. marmoratus) is a Mediterranean immigrant, entering the Black Sea fauna about 7000–12,000 years ago (Miller 1965; Zaitsev and Mamaev 1997). So, it seems probable that the parasite migrated with its host. But, taking into account the presence of A. coelomicola in the Baltic and North seas, we can consider A. coelomicola as a Boreal-Atlantic species, which is probably part of a relict fauna (from Upper Miocene) in the Black and Baltic seas. The parasite can use different species of mud-snail of Hydrobia group as first intermediate host. So, for the Mediterranean Sea, Ecrobia ventrosa (Montagu, 1803) is recorded as the snail host (Maillard 1973), but Semisalsa stagnorum (Gmelin, 1791) is mentioned for the Baltic Sea (Vaes 1978; Reimer 1970). According to Zander et al. (2002), in the Baltic Sea, both Peringia ulvae (Pennant, 1777) and E. ventrosa are used by Aphalloides as first intermediate host. Two snails of the Hydrobia group, E. ventrosa and Hydrobia acuta (Draparnaud, 1805), are common in the Black Sea fauna (Butenko 2000; Osikowski et al. 2016). These molluscs are related to a group of the European lagoonal species, distribution today in the region of the old Tethys Sea, including eastwards into the Ponto-Caspian basin, in all European coasts, also to the eastern seaboard of North America (Barnes 1989). The wide distribution of snails of Hydrobia group, especially of E. ventrosa as main host, can support the Boreal-Atlantic status of A. coelomicola.
References
Bakke TA (1980) A scanning electron microscope study of the microtopography of Aphalloides timmi Reimer, 1970 (Digenea; Cryptogonimidae). Fauna Norvegica (Series A) 1:38–44
Barnes RSK (1989) What, if anything, is a brackish-water fauna? Trans Royal Soc Edinburgh (Earth Sci) 80:235–240. doi:10.1017/S0263593300028674
Bayssade-Dufour C, Maillard C (1982) Discussion sur la position taxonomique d’Aphalloides coelomicola Dollfus, Chabaud et Golvan, 1957 (Trematoda, Opistorchioidea). Ann Parasitol Hum Comp 57(6):549–553
Bouchereau JL, Guelorget O (1998) Comparison of three Gobiidae (Teleostei) life history strategies over their geographical range. Oceanol Acta 21:503–517. doi:10.1016/S0399-1784(98)80034-0
Butenko OI (2000) Molluscs of coastal shallow waters of the Odessa Bay. Ekologiya Morya 52:44–47 (in Russian with English summary)
Chernyshenko AS, Sventsytskaya GA (1970) Sravnitelno-ekologicheskaya kharakteristika gelmintofauny bychkov (sem. Gobiidae) limanov Severo-Zapadnogo Prichernomorya i Odesskogo zaliva [Comparative ecological characteristic of helminth fauna of gobies (fam. Gobiidae) of lagoons of North-Western Black Sea region and the Gulf of Odessa]. In: Voprosy morskoy parazitologii [Issues of marine parasitology]. Naukova Dumka, Kiev, pp 135–137 (in Russian)
Cribb TH, Bray RA (2010) Gut wash, body soak, blender and heat-fixation: approaches to the effective collection, fixation and preservation of trematodes of fishes. Syst Parasitol 76:1–7. doi:10.1007/s11230-010-9229-z
Dethier MN (1992) Classifying marine and estuarine natural communities: an alternative to the coward in system. J Nat Areas 12:90–100
Dollfus RP, Chabaud AG, Golvan YJ (1957) Helminthes de la région de Banyuls. V. Nouveau Distome Aphalloïdes coelomicola n. gen. n. sp. de la cavité générale d’un Gobius d’eau saumâtre. Annales de Parasitologie 32:29–40
Georgiev B, Biserkov V, Genov T (1986) In toto staining method for cestodes with iron acetocarmine. Helminthologia 23:279–281
Jayawardena UA, Tkach VV, Navaratne AN, Amerasinghe PH, Rajakaruna RS (2013) Malformations and mortality in the Asian common toad induced by exposure to pleurolophocercous cercariae (Trematoda: Cryptogonimidae). Parasitol Int 62(3):246–252. doi:10.1016/j.parint.2013.01.003
Josten N, Detloff KC, Zander CD (2009) Analysis of a parasite supra community from the Flensburg fjord. Parasitol Res 104:449–461. doi:10.1007/s00436-008-1218-7
Krasnovyd V, Kvach Y, Drobiniak O (2012) The parasite fauna of the gobiid fish (Actinopterygii, Gobiidae) in the Sykhyi Lyman, Black Sea. Vestnik Zoologii 46(6):483–490. doi:10.2478/v10058-012-0041-x
Kearse M, Moir R, Wilson A, Stones-Havas S, Cheung M, Sturrock S, Buxton S, Cooper A, Markowitz S, Duran C, Thierer T, Ashton B, Mentjies P, Drummond A (2012) Geneious basic: an integrated and extendable desktop software platform for the organization and analysis of sequence data. Bioinformatics 28(12):1647–1649. doi:10.1093/bioinformatics/bts199
Kvach Y (2010) Helminths of the marbled goby (Pomatoschistus marmoratus), a Mediterranean immigrant in the Black Sea fauna. Vestnik Zoologii 44(6):509–518. doi:10.2478/v10058-010-0034-6
Kvach Y, Winkler HM (2011) The colonization of the invasive round goby Neogobius melanostomus by parasites in new localities in the southwestern Baltic Sea. Parasitol Res 109(3):769–780. doi:10.1007/s00436-011-2321-8
Lefebvre F, Poulin R (2005) Progenesis in digenean trematodes: a taxonomic and synthetic overview of species reproducing in their second intermediate hosts. Parasitology 130:587–605. doi:10.1017/S0031182004007103
Littlewood DTJ, Rohde K, Clough KA (1997) Parasite speciation within or between host species?—Phylogenetic evidence from site-specific polystome monogeneans. Int J Parasitol 27:1289–1297. doi:10.1016/S0020-7519(97)00086-6
Littlewood DTJ (1994) Molecular phylogenetics of cupped oysters based on partial 28S rRNA gene sequences. Mol Phylogenet Evol 3(3):221–229. doi:10.1006/mpev.1994.1024
Maillard C (1973) Mise en évidence du cycle évolutif abrégé d'Aphalloïdes coelomicola Dollfus, Chabaud et Golvan, 1957 (trematoda). Notion d'hôte historique. CR Acad Scie Paris (Sér D) 277:317–320
Mazzoldi C, Poltronieri C, Rasotto MB (2002) Egg size variability and mating system in the marbled goby Pomatoschistus marmoratus (Pisces: Gobiidae). Mar Ecol Prog Ser 233:231–239. doi:10.3354/meps233231
Miller PJ (1965) Relictogobius kryzhanovskii and the penetration of Mediterranean gobies into the Black Sea. Nature 208:474–475. doi:10.1038/208474a0
Miller PJ (1986) Gobiidae. In: Whitehead PJP, Bauchot M-L, Hureau J-C, Nielsen J, Tortonese E (eds) Fishes of the North-eastern Atlantic and the Mediterranean, volume 3. UNESCO, Paris, pp 1019–1085
Miller TL, Cribb TH (2008a) Eight new species of Siphoderina Manter, 1934 (Digenea, Cryptogonimidae) infecting Lutjanidae and Haemulidae (Perciformes) off Australia. Acta Parasitol 53(4):344–364. doi:10.2478/s11686-008-0053-4
Miller TL, Cribb TH (2008b) Family Cryptogonimidae Ward, 1917. In: Bray RA, Gibson DI, Jones A (eds) Keys to the Trematoda, Vol. 3. CAB International, Wallingford, pp 51–112
Naydenova NN (1970) K sistematicheskomu polozheniju Aphalloides coelomicola Dollfus, Chabaud et Golvan, 1958 [to systematic position of Aphalloides coelomicola Dollfus, Chabaud et Golvan, 1958]. Biologiya Morya (Kiev) 20:74–84 (in Russian)
Naydenova NN (1974) Parazitofauna ryb semeystva Gobiidae Chernogo i Azovskogo morey [the parasite fauna of fishes of Gobiidae Family in the Black Sea and the sea of Azov]. Naukova Dumka, Kiev, p 182 (in Russian)
Nunn G (1992) Nematode molecular evolution. An investigation of evolutionary patterns among nematodes based upon DNA sequences. PhD dissertation, University of Nottingham, UK
Osikowski A, Hofman S, Georgiev D, Kalcheva S, Falniowski A (2016) Aquatic snails Ecrobia maritima (Milaschewitsch, 1916) and E. ventrosa (Montagu, 1803) (Caenogastropoda: Hydrobiidae) in the East Mediterranean and Black Sea. Annales Zoologici 66:477–486. doi:10.3161/00034541ANZ2016.66.3.012
Pampoulie C, Lambert A, Rosecchi E, Crivelli AJ, Bouchereaut J-L, Morand S (2000) Host death: a necessary condition for the transmission of Aphalloides coelomicola Dollfus, Chabaud, and Golvan, 1957 (Digenea, Cryptogonimidae)? J Parasitol 86(2):416–417. doi:10.1645/0022-3395(2000)086[0416:HDANCF]2.0.CO;2
Pampoulie C, Morand S, Lambert A, Rosecchi E, Bouchereaut J-L, Crivelli AJ (1999) Influence of the trematode Aphalloïdes cœlomicola Dollfus, Chabaud & Golvan, 1957 on the fecundity and survival of Pomatoschistus microps (Krøyer, 1838) (Teleostei: Gobiidae). Parasitology 119:61–67. doi:10.1017/S0031182099004412
Pampoulie C, Rosecchi E, Bouchereaut J-L, Crivelli AJ (2004) Do environmental changes influence the occurrence and effect of parasites? J Negat Results: Ecol Evol Biol 1:8–15
Parukhin AM, Machkevskiy VK, Ilchenko SV (1983) Osobennosti gelmintofauny ryb Yagorlytskogo zaliva [the characteristics of the helminth fauna of fish of the Gulf of Yahorlyk]. Vestnik Zoologii 1:33–37 (in Russian)
Reimer L (1970) Digene Trematoden und Cestoden der Ostseefische als natürliche Fischmarken. Parasitologische SchriftenReiche 20:1–144
Reimer LW, Hildebrand A, Scharberth D, Walter U (1996) Trematodes of the brackish waters of the Baltic Sea and their distribution together with that of related species in other European areas. Appl Parasitol 37:177–185
Sasal P, Pampoulie C (2000) Asymmetry, reproductive success and parasitism of Pomatoschistus microps in a French lagoon. J Fish Biol 57:382–390. doi:10.1111/j.1095-8649.2000.tb02178.x
Stoyanov B, Neov B, Pankov P, Radoslavov G, Hristov P, Georgiev BB (2015) Redescription of Aphalloides coelomicola Dollfus, Chabaud & Golvan, 1957 (Digenea, Opisthorchioidea) based on specimens from Knipowitschia caucasica (Berg) (Actinopterygii, Gobionellidae) from a Black Sea lagoon, with comments on the systematic position of the genus. Syst Parasitol 91:1–12. doi:10.1007/s11230-015-9559-y
Thacker CE (2009) Phylogeny of Gobioidei and placement within Acanthomorpha, with a new classification and investigation of diversification and character evolution. Copeia 2009(1):93–104. doi:10.1643/CI-08-004
Vaes M (1978) Infection of the common goby, Pomatoschistus microps, with Aphalloides coelomicola (Trematoda Digenea). Vlaams Diergeneeskundig Tijdschrift 47(3):274–278
Zaitsev Y, Mamaev V (1997) Biological diversity in the Black Sea: a study of change and decline. UN Publication, New York, p 208
Zaitsev YP (1998) The most blue sea in the world. UN Publication, New York, p 142 (in Russian)
Zander CD (2003) Four-year monitoring of parasite communities in gobiid fishes of the south-western Baltic. I. Guild and component community. Parasitol Res 90:502–511. doi:10.1007/s00436-003-0887-5
Zander CD (2005) Comparative studies on goby (Teleostei) parasite communities from the North and Baltic Sea. Parasitol Res 96:62–68. doi:10.1007/s00436-005-1327-5
Zander CD, Koçoglu O, Skroblies M, Strohbach U (2002) Parasite populations and communities from the shallow littoral of the Orther Bight (Fehmarn, SW Baltic Sea). Parasitol Res 88:734–744. doi:10.1007/s00436-002-0652-1
Zander CD, Reimer LW (2002) Parasitism at the ecosystem level in the Baltic Sea. Parasitology 124:S119–S135. doi:10.1017/S0031182002001567
Zander CD, Reimer LW, Barz K (1999) Parasite communities of the Salzhaff (Northwest Mecklenburg, Baltic Sea). I. Structure and dynamics of communities of littoral fish, especially small-sized fish. Parasitol Res 85:356–372. doi:10.1007/s004360050562
Zander CD, Reimer LW, Barz K, Dietel G, Strohbach U (2000) Parasite communities of the Salzhaff (Northwest Mecklenburg, Baltic Sea). II. Guild communities, with special regard to snails, benthic crustaceans, and small-sized fish. Parasitol Res 86:359–372. doi:10.1007/s004360050681
Zenkevich LA (1963) Biologiya morey SSSR [Biology of seas of USSR]. Izdatelsto AN SSSR, Moskva, p 302 (in Russian)
Acknowledgements
The study received financial support from the European Centre of Ichthyoparasitology–Centre of Excellence, Grant Agency of the Czech Republic Project No. P505/12/G112. We thank Kevin Roche for English correction.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article

Cite this article
Kvach, Y., Bryjová, A., Sasal, P. et al. A revision of the genus Aphalloides (Digenea: Cryptogonimidae), parasites of European brackish water fishes. Parasitol Res 116, 1973–1980 (2017). https://doi.org/10.1007/s00436-017-5480-4
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00436-017-5480-4