
Abstract
Freshly emerged virgin female Sarcophaga ruficornis were topically treated with different doses of pyriproxyfen to test the efficacy on reproduction and subsequent F1 progeny. The results included mortality of the treated adults, significant reduction in fecundity, more than 90 % inhibition in larvae production, mortality in F1 generation during larval instars, reduction in pupariation, and adult emergence and production of deformed adults. There was a dose-dependent response showing high degree of correlation in the doses administered and deformities observed. The effects in F1 generation show that an intraovarial transfer of pyriproxyfen was responsible for the various deformations observed, showing the potency of juvenoid pyriproxyfen for longer durations across generations. This is the first study that clearly demonstrates the efficacy of pyriproxyfen against reproduction in S. ruficornis and its potential for the management of this notorious pest of medico-veterinary importance.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Sarcophaga ruficornis, the flesh fly, is a larviparous cyclorrhaphous diptera causing myiasis in animals and humans. Myiasis is the invasion of tissues and organs in humans and animals by larvae of fly that feed on the host tissues, body fluids, or ingested food or live as parasites in the skin, subcutaneous tissues, mouth, stomach, eyes, nose, ears, intestines, urinogenital system, and other soft tissues of the body (Kumarsinghe et al. 2000). In humans, cases of urinogenital (Dincer et al. 1995), ophthalmic (Tuncer et al. 1996), cutaneous (Merino et al. 2000), oral (Yazar et al. 2005), and nasal myiasis (Turk et al. 2006) have been reported to be caused by Sarcophaga species. The larvae also infest meat and fish and are a menace to the meat processing industry all around the globe. Adults act as carriers and vectors of many disease causing microbes, pathogens, parasites, eggs, and cysts causing various diseases. It is suspected carrier of leprosy bacilli and also poses threat of spreading many diseases (Sukontason et al. 2007). It is also associated with pathogenic strains of Escherichia coli, causative agent of gastrointestinal diseases, and food-borne sickness (Maike et al. 2007) apart from being a source of annoyance.
Chemical control methods pose several problems to the environment causing biomagnifictaion, induction of resistance and also pose threat to nontarget and beneficial insects. The search for safe and effective control measures led to the use of insect growth regulators—compounds that mimic the action of insect hormones and hamper the growth, development, metamorphosis, reproduction, and finally killing the insect pest.
Pyriproxyfen, a stable aromatic, nonterpenoidal juvenoid has been effectively used for the control of pests like whiteflies, mealworms, scales, thrips, and cutworms that are insensitive or resistant to chemical/conventional insecticides (Ishaaya and Horowitz 1992; Oouchi and Langley 2005; Aribi et al. 2006). It is a potent endocrine disruptor and acts by overloading the insect hormonal system being toxic in their embryonic, larval, and reproductive stages.
Pyriproxyfen is photo-stable, selective in action, easily biodegradable, compatible with biological control, and integrated pest management. It possesses minimal threat of resistance and is safe for environment besides being nontoxic to beneficial insects like bees and bumble bees (De Wael et al. 1995), a range of predatory species (Peleg 1988; Delbeke et al. 1997; Liu and Stanley 1997), and vertebrates and natural enemies of target pests (Meola et al. 2000; Medina et al. 2003; Wang et al. 2005; Ishaaya and Horowitz 2007).
Pyriproxyfen has been extensively used for the control of Blatella germanica (Koehler and Patterson 1991), Culex sp. (Chavasse et al. 1995), many ticks and fleas (Strey et al. 2001), California red scale, Aonidiella aurantii (Eliahu et al. 2007), and Musca domestica (Sulaiman et al. 2008), but there is a dearth of studies pertaining to use of pyriproxyfen in control of cyclorrhaphous diptera which prompted the authors to study the effects of pyriproxyfen on reproduction of S. ruficornis considering the medical and veterinary importance. The present study intends to study the effects of administration of JH analog pyriproxyfen to the adults and subsequent F1 progeny of myiasis causing medico-veterinary pest S. ruficornis.
Materials and methods
Test insect
Sarcophaga ruficornis was obtained from a colony reared in the Department of Zoology, University of Allahabad, Allahabad (25° 27′ N 81° 44′ E), UP, India, since 2005. Adults were also captured from wild and introduced to maintain vigor of the colony. The adult flies were reared at 28 ± 2 °C and 75 ± 5 % relative humidity and 10 L/14D photoperiod in 30 × 30 × 30 cm cages. Powdered sugar, 10 % honey solution, and water were provided for feeding. Fresh pieces of goat’s liver were provided for larviposition and also constituted a standard protein source. The larvae laid in a single laying were transferred to 15 × 10-cm glass troughs capped with muslin cloth and provided with fresh pieces of the liver and kidney for feeding. Once the larvae stopped feeding, they were shifted to glass troughs provided with sawdust for pupation. Pupae formed were shifted to 500-ml jars until emergence.
Test compound
Pyriproxyfen, 2-[1-methyl-2-(4-phenoxyphenoxy)ethoxy]pyridine (Vetranal®) A.I. 99.9 % was purchased from Sigma–Aldrich Chemicals Co. USA. Known quantities of pyriproxyfen was weighed and dissolved in 1 ml of acetone to get the desired concentration of doses. Each time fresh doses were prepared to avoid any differences in concentration due to evaporation.
Experimental procedure
Freshly emerged virgin adults were anesthetized using few drops of diethyl ether. Males and females were separated and divided into batches of 10 each. The females were treated with 50 and 100 μg/5 μl/adult of pyriproxyfen using 10-μl Hamilton syringe (Sigma–Aldrich Chemicals Co. USA) on the ventral surface of the abdomen. The controls were treated with diluents acetone only. After the treatment, the females were paired with untreated males in the ratio of 1:2 and kept in cages with similar conditions mentioned above. Fresh pieces of the liver were provided for larviposition in both the treated and control batches. The larvae were collected for 10 days after treatment or up to when treated adults died whichever was earlier. Number of larvae laid were counted for each laying separately under stereo zoom SMZ1000 microscope (Nikon Corp., Japan) and reared as mentioned earlier. Once the adults from F1 generation puparia of controls emerged, the treated batches were checked for emergence and puparia from which adults did not emerge on their own were dissected to check the growth of the imago. The p-a mosaics, pharate adults, and deformed adults formed were fixed in Bouin’s fluid and preserved in 70 % ethyl alcohol for further microscopic studies and photography.
Data analysis
Reduction in fecundity was calculated by the formula:
where Nt = no. of larvae in treated batch.
Nc = no. of larvae in control batch.
All the data collected was subjected to statistical analysis using Biostat 5.8.0 professional 2009 software (Analyst Softech, USA). Correlation coefficient was calculated to find out correlation between doses applied and deformities observed. Photography was done using Nikon stereo zoom SMZ1000 microscope (Nikon Corp., Japan).
Results
The treatment of freshly emerged virgin female S. ruficornis with 50 and 100 μg/5 μl/adult dose of pyriproxyfen resulted into mortality of the adults as well as reduced fecundity in both the treated groups. Several latent effects were also observed in the F1 progeny resulting from treated females. These included mortality in larval stage, reduced pupariation, formation of p-a mosaics, reduced adult emergence, and formation of deformed adults with malformed mouthparts, wings, legs and abdomen along with deformity in sclerotization.
Adult mortality was 20 % in both the treated groups, i.e., at 50- and 100-μg doses as compared to controls where no mortality was observed (r = 0.87; p ≤ 0.05) showing a “knockdown” effect as these treated adults died within 24 h after treatment (Table 1).
Fecundity was drastically reduced in a dose-dependent manner, more being at the higher dose of 100 μg. Over a period of 10 days from the day of treatment or till the death of treated females, only 6.2 and 5.1 % larvae were laid down at 50- and 100-μg doses as compared to 100 % in control group with inhibition rate (IR %) being 92.1 and 93.6 % at 50- and 100-μg doses, respectively (r = −0.87; p ≤ 0.05), showing a negative correlation between the doses applied and fecundity (Table 1). Not only the inhibition rate was high in treated groups, the larviposition was not regular as observed in control batch. An intermittent laying was observed in both the treated groups.
Larval mortality in F1 generation was also observed in 1st (L1) and 2nd (L2) larval instars, both in controls and treated groups. The larval mortality in controls were very low being only 0.7 and 0.1 % in L1 and L2 instars as compared to 12.9 and 10.5 % at 50 μg and 17.0 and 4.9 % at 100 μg in L1 and L2 instars [r = 0.96 (L1) and r = 0.46 (L2), (p ≤ 0.05) respectively]. No mortality was observed during third instar at any dose. The young larvae were more susceptible to pyriproxyfen (Table 1).
Normal puparia formation was reduced in both the treated groups as compared to the controls. The percentage normal puparia formed reduced from 99.1 % in controls to 76.5 and 78.0 % at 50- and 100-μg doses, respectively, with coefficient of correlation r = −0.83 (p ≤ 0.05) (Table 1).
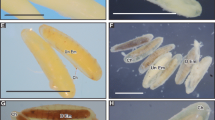
Pupal–adult mosaics were also formed and the percentage being 0.1 % in controls which increased to 10.0 at 50 μg and 36.0 % at 100-μg dose. The correlation coefficient was r = 0.96 (p ≤ 0.05) showing a positive and dose-dependent correlation between doses applied and p-a mosaics produced (Table 1). The p-a mosaics formed were defined on the basis of degree of development and presence of pupal and adult characteristics (Plate 1a) (Table 2).

Types of deformities produced in the F1 progeny after topical administration of pyriproxyfen to female adult flies of Sarcophaga ruficornis. a Pupal–adult mosaic, 50 μg. b Pharate adult, 100 μg. c–d Deformed adults, 50 μg. e–f Defects in genitalia, 50 μg. g Half eclosed defective adult, 100 μg. h–j Deformed adults, 100 μg (Abn Ad = abnormal adult; Def W = defective wings; E = eyes; Ev G = everted genitalia; G = genitalia; PM = pupal mass; Pu Abd = pupal abdomen; Pu = puparia; Rd Abd = reduced abdomen; Scl = Sclerite; Th = thorax) (bar = 1 mm)
Adult emergence was reduced in a dose-dependent manner from 99.0 % in control batch to 90.0 and 63.3 % at 50- and 100-μg doses, respectively, with a correlation coefficient r = −0.96 (p ≤ 0.05). No deformities were observed in emerged F1 adults in control batch while 23.0 % deformed adults at 50 μg and 22.7 % deformed adults at 100 μg was observed (r = 0.86; p ≤ 0.05). The deformities were mainly in the mouthparts, legs, wings, and abdomen (Plate 1c–j) (Table 2). The percentage of normal adults produced declined gradually in a dose-dependent manner with correlation coefficient r = −0.99 (p ≤ 0.05) showing a negative correlation between the doses applied and normal adults produced (Table 1).
Discussion
Treatment of freshly emerged virgin flesh fly, S. ruficornis, with 50 and 100 μg/adult doses resulted in toxicity into adult flies as well as reduction in fecundity. Apart from these effects on adults, several latent or delayed effects were seen in the F1 generation produced from the treated flies. These included mortality during larval instars, reduced pupariation, decreased adult emergence, formation of pupal–adult mosaics, and production of deformed adults having abnormalities in the mouthparts, wings, legs, and abdomen.
Adult mortality as a result of pyriproxyfen was observed at both the doses. These flies died within 24 h of treatment showing their sensitivity toward pyriproxyfen. “Knockdown” effect may be due to lethal effects of IGR affecting the vital mechanisms and physiology of the treated insects. Death of adults due to IGR/JH analogs has been previously reported in Blatella germanica (Ross and Cochran 1990, 1991), Anopheles punctatus (Okazawa et al. 1991), Myzus persicae (Hatakoshi et al. 1991), Musca domestica and B. germanica (Kwada et al. 1992), Aphis gossypi (Wood and Godfrey 1998), Tribolium castaneum and Sitophilus oryzae (Kostyukovsky et al. 2000), Lipaphis erysimi (Chen and Liu 2002), Thrips tabaci (Liu 2003), Hyposoter didymator (Schneider et al. 2004), Aedes aegypti (Darriet and Corbel 2006; Alejandro et al. 2009; Emilia et al. 2014), Diaphorina citri (Boina et al. 2009), and Eurygaster integriceps (Mojaver and Bandani 2010).
Reduced fecundity was the most prominent effect observed as a result of pyriproxyfen treatment. Reduced fecundity and suppressed oviposition by pyriproxyfen in diamond back moth, Plutella xylostella, was observed by Oouchi (2005). Langley et al. (1990) used pyriproxyfen as a sterilant for the successful control of tse tse flies, Glossina moritans moritans as congruent with the present study. Ross and Cochran (1990, 1991) while studying the effects of 3 IGRs–pyriproxyfen, fenoxycarb, and diflubenzuron found that these IGRs not only caused mortality but also induced female sterility, reduced productive matings, and caused deleterious effects on ovaries producing unfertilized eggs in German cockroach B. germanica.
Kwada et al. (1992) found that pyriproxyfen treatment not only reduced the number of eggs but also reduced the hatchability of eggs laid by treated females of M. domestica and B. germanica. Reduced fecundity due to pyriproxyfen was also observed in T. tabaci (Liu 2003). Boina et al. (2009) observed that in D. citri, pyriproxyfen treatment of newly emerged adults did not produce acute lethal effects as observed in present study but resulted into reduced fecundity and egg viability as found in the present study. Varloud and Hodgkins (2015) studied the effects of combined formulation of dinotefuron, pyriproxyfen, and permethrin (DPP) against cat flea Ctenocephalides felis and brown dog ticks Rhiphicephalus sanguineus and demonstrated that DPP was highly effective against adult ticks and fleas. Also, DPP reduced more than 90 % egg hatch and adult emergence in cat fleas congruent with the present study.
JHAs cause inhibitory effect on reproduction by affecting the physiology and hormonal homeostasis when adults are treated. Hatakoshi and Hirano (1990) proposed two principal roles of JH mimics in adults—(a) inhibition of “oviposition stimulating haemolymph factor” to cause decrease in number of eggs and (b) direct effects on ovaries.
Reproductive cycle in adults proceeds in different steps—(a) vitellogenin (yolk protein) synthesis by fat body, (b) separation of new follicle from the germarium, (c) previtellogenic growth of the oocyte, and (d) vitellogenesis (yolk protein uptake by oocyte). All these processes are orchestrated by ecdysteroids, juvenile hormone, and various neuroendocrine hormones (Nijhout 1998) and can be used as tools or control points for regulation of reproduction. Any exogenous substance that disturbs or interferes with intrinsic hormonal levels can affect the various events associated with reproduction. In diptera, the vitellogenin synthesis is stimulated by ecdysteroids (Nijhout 1998), and JH regulates growth of ovaries and behavior and mating. These two hormones along with ovarian ecdysteroidogenic hormone (OEH) or egg development neurosecretory hormones (EDNH) secreted from neurosecretory cells (NSCs) of the brain regulate the reproductive cycle (Nijhout 1998).
JHA pyriproxyfen, when administered to the freshly emerged adults in the present study, appears to have exerted its detrimental effects on one or more steps during reproductive cycle/maturity as a consequence of its presence in large quantities in the insect system at an inappropriate time. The high titers of JHA might have suppressed the ecdysteroid titers which are normally high in the absence of or low JH levels, and thus disrupting the vitellogenin synthesis and consequently leading to reduced fecundity. The exact mode of JHA pyriproxyfen action on various steps involving reproduction is unknown and warrants molecular level investigations. It seems that in the present study, the JHA pyriproxyfen exerts its effects on the actions of all the three hormones—the JH, ecdysteroids, and the EDNH, resulting in the disruption of not only the maturation of ovaries but also on the synthesis and uptake of vitellogenin and egg development as evident by extremely low fecundity as compared to that of controls.
Pyriproxyfen also exerted several latent effects that were expressed in the F1 generation progeny of the treated females. This shows that the JHA pyriproxyfen has been transovarially passed on to the developing embryos/larvae and caused several deformities and malfunctions as observed in the larvae that were laid. Larval mortality was observed both in first and second instar stage of the treated groups in a dose-dependent manner showing the sensitivity of the young larvae to pyriproxyfen. Larvicidal properties of pyriproxyfen is well documented in literature against a wide range of pests including mosquitoes and citrus butterfly (Invest and Lucas 2008; Alejandro et al. 2009; Boina et al. 2009; Mojaver and Bandani 2010; Singh and Kumar 2011; Emilia et al. 2014).
Pyriproxyfen treatment also caused reduction in pupariation and adult emergence in the F1 generation in a dose-dependent manner. Several previous studies show the effects of IGRs on various insect species congruent with the present study (Aribi et al. 2006; Andrighetti et al. 2008; Boina et al. 2009).
Moreover, various grades of pupal–adult mosaics and deformed adults having several malformations were also produced in the F1 progeny due to pyriproxyfen treatment of parent females. Insect metamorphosis is mainly regulated by JH and ecdysone titers, and exogenous application of JHA disrupts the normal hormonal levels derailing the normal developmental process. In the present study, pyriproxyfen induces delayed effects during the development and metamorphosis of F1 progeny of treated females resulting into formation of mosaics and deformed adults with defects in wings, mouthparts, and appendages. Pyriproxyfen being highly stable and biologically active continues to persist in the system for a longer time to induce detrimental effects or to suppress the intrinsic ecdysone levels along with its effects on various other neurohormones like eclosion hormone (EH) affecting the adult emergence and bursicon affecting the sclerotization of the deformed adults. Any exogenous substance that acts as endocrine disruptor produces various deformities in development and is always lethal as seen in the present study. Many previous studies have shown the formation of mosaics as well as deformed adults as a result of IGR/JHA treatments (Koehler and Patterson 1991; Eto 1990; Koçak and Kilinçer 1997; Richardson and Lagos 2007).
Juvenoid pyriproxyfen was found to be very potent in inducing inhibitory effects on reproduction of S. ruficornis significantly reducing fecundity and also causing various detrimental effects in the F1 progeny produced from the treated females. Pyriproxyfen being a pest specific, stable, and nontoxic JHA has promising prospects in fulfilling the lacuna in control of pest of medical and veterinary importance S. ruficornis.
References
Alejandro L, Harburguer L, Licastro S, Zerba E, Masuh H (2009) Efficacy of new combined larvicidal-adulticidal ultralow volume formulation against Aedes aegypti (Diptera: Culicidae), vector of dengue. Parasitol Res 104:1101–1107
Andrighetti MTM, Cerone F, Rigueti M, Galvani KC, Macoris MLG (2008) Effect of pyriproxyfen in Aedes aegypti population with different levels of susceptibility to the organophosphate temophos. Dengue Bull 32:186–198
Aribi N, Smagghe G, Lakbar S, Soltani-Mazouni N, Soltani N (2006) Effects of pyriproxyfen, a juvenile hormone analog, on development of the mealworm, Tenebrio molitor. Pestic Biochem Physiol 84:55–62
Boina DR, Rogers ME, Wang N, Stelinski LL (2009) Effect of pyriproxyfen, a juvenile hormone mimic, on egg hatch, nymph development, adult emergence and reproduction of the Asian citrus psyllid, Diaphorina citri Kuwayama. Pest Manag Sci 66:349–357
Chavasse DC, Lines JD, Ichimori K, Majala AR, Minjas JN, Marijani J (1995) Mosquito-control in Dar-es-Salaam 2. Impact of expanded polystyrene beads and pyriproxyfen treatment of breeding sites on Culex quinquefasciatus densities. Med Vet Entomol 9:147–154
Chen TY, Liu TX (2002) Susceptibility of immature stages of Chrysoperla rufilabris (Neurop., Chrysopidae) to pyriproxyfen, a juvenile hormone analog. J Appl Entomol 126:125–129
Darriet F, Corbel V (2006) Laboratory evaluation of pyriproxyfen and spinosad, alone and in combination against Aedes aegypti larvae. J Med Entomol 43(6):1190–1194
De Wael L, DeGreef M, VanLacre O (1995) Toxicity of pyriproxyfen and fenoxycarb to bumble bee brood using a new method for testing insect growth regulators. J Agric Res 34:3–8
Delbeke F, Vercrysse P, Tirryl DeCleraq P, Deghele D (1997) Toxicity of diflubenzuron, pyriproxyfen, imidacloprid and diafenthiuron tothe predatory bug orius laevigatus. (Het: Anthocoridae). Entomophaga 72:349–358
Dincer S, Tanyukesel M, Kuçuk T (1995) Two cases of human urinogenital myiasis caused by Psycoda spp. (Diptera: Nematocera) and Sarcophaga spp. (Diptera: Cyclorrhapha) in Ankara. Turk Parazitol Derg 19(3):402–408
Eliahu M, Blumberg D, Horowitz AR, Ishaaya I (2007) Effect of pyriproxyfen on developing stages and embryogenesis of California red scale (CRS), Aonidiella aurantii. Pest Manag Sci 63(8):743–746
Emilia S, Laura J, Eduardo Z, Licastro S (2014) Aedes aegypti (Diptera:Culicidae): evaluation of natural long-lasting materials containing pyriproxyfen to improve control strategies. Parasitol Res 113:3355–3360
Eto M (1990) Biochemical mechanism of insecticidal activities. In: Hang G, Hoffman H (eds) Chemistry of plant protection, Springer Verlag 6, p 65-107
Hatakoshi M, Hirano M (1990) Effects of S-71639 on reproduction of Spodoptera litura. In: Hoshi M, Yamashita O (eds) Advances in invertebrate reproduction, Elsevier Science Publishers, Amsterdam, p 429-434
Hatakoshi M, Shono Y, Yamamoto H, Hirano M (1991) Effects of juvenile hormone analog pyriproxyfen on Myzus persicae and Unaspis yononeusis. Appl Entomol Zool 26:412–414
Invest JF, Lucas JR (2008) Pyriproxyfen as a mosquito larvicide. In: Robinson H, Daniel B (eds) Proceedings of Sixth International Conference on Urban Pests. William Oak Press, Hungary
Ishaaya I, Horowitz AR (1992) Novel phenoxy hormone analog (pyriproxyfen) supresses embryogenesis and adult emergence of sweet potato whitefly. J Econ Entomol 85:2113–2117
Ishaaya I, Horowitz AR (2007) In focus: IPM using novel insecticides and other approaches. Pest Manag Sci 63:729
Koçak E, Kilinçer N (1997) Juvenil hormone analoğu methopreń in pamuk yaprak kurdu Spodoptera littoralis Boist. (Lep: Noctuidae)' na etkileri: 1. pupa ve yumurta etkiler. Bitki kor Bült 37:163–172
Koehler PG, Patterson RJ (1991) Incorporation of pyriproxyfen in a german cockroach (Dictyoptera: Blatellidae) management program. J Econ Entomol 84:917–921
Kostyukovsky M, Chen B, Atmsi S, Shaaya E (2000) Biological activity of two juvenoids and two ecdysteroids against three stored product insects. Insect Biochem Mol Biol 30:819–897
Kumarsinghe SPW, Karunaveera ND, Ihalamulla RL (2000) A study of cutaneous myiasis in Sri Lanka. Int J Dermatol 39:689–694
Kwada H, Senbo S, Abe Y (1992) Effects of pyriproxyfen on the reproduction of the housefly, Musca domestica, and the german cockroach, Blattella germanica. Jpn J Sanit Zool 43:169–175
Langley PA, Felton T, Stafford K, Oouchi H (1990) Formulation of pyriproxyfen, a juvenile hormone mimic for tse-tse control. Med Vet Entomol 4:127–133
Liu TX (2003) Effects of a juvenile hormone analog, pyriproxyfen, on Thrips tabaci (Thysanoptera: Thripidae). Pest Manag Sci 59:904–912
Liu TX, Stanley PA (1997) Effects of pyriproxyfen on three species of Encarsia (Hymemoptera: Aphelinidae), endoparasite of Bemisia argentifolii (Homotera: Alegrodidae). J Econ Entomol 90:404–411
Maike F, Klimpel S, Melhorn H, Sievert K, Messler S, Pfeffer K (2007) Pilot study on synanthropic flies (e.g. Musca, Sarcophaga, Calliphora, fannia, Lucilia, Stomoxys) as vectors of pathogenic microorganisms. Parasitol Res 101:243–246
Medina P, Budia FEP, Viñuela E (2003) Effects of three modern insecticides, pyriproxyfen, spinosad and tebufenozide on survival and reproduction of Chrysoperla carnea adults. Ann Appl Biol 142:55–61
Meola R, Meier K, Dean S, Bhaskaran G (2000) Effects of pyriproxyfen in the blood diet of catfleas on adult survival, egg development and larval development. J Med Entomol 37:503–506
Merino FJ, Campos A, Nebreda T, Canovas C, Cueza F (2000) Cutaneous myiasis by Sarcophaga sp. Enferm Infecc Microbiol Clin 18(1):19–21
Mojaver M, Bandani AR (2010) Effects of the insect growth regulator pyriproxyfen on immature stages of Sunn pest, Eurygaster integriceps Puton (Heteroptera: Scutelleridae). Mun Ent Zool 5(1):187–197
Nijhout HF (1998) Insect Hormones. Princeton University Press, Princeton
Okazawa T, Bakote’e B, Suzuki H, Kwada H, Kere N (1991) Field evaluation of an insect growth regulator, pyriproxyfen, against Anopheles punctatus on north Guadalcanal, Solomon Islands. J Am Mosq Control Assoc 7(4):604–607
Oouchi H (2005) Insecticidal properties of a juvenoid pyriproxyfen, on all life stages of the diamond back moth, Plutella xyllostella (Lepidoptera: Yponomeutidae). Appl Entomol Zool 40:145–149
Oouchi H, Langley P (2005) Control of greenhouse whitefly (Trialeurodes vaporariorum) using visually attractive targets impregnated with pyriproxyfen. J Pestic Sci 30(1):50–52
Peleg BA (1988) Effects of a new phenoxy juvenile hormone analog on California red scale (Homoptera: Coccidae), Florida wax scale (Homptera: Coccidae) and the ectoparasite Aphytias holoxanthus DeBach (Hymemoptera: Aphylinidae). J Econ Entomol 81:88–92
Richardson ML, Lagos DM (2007) Effects of a juvenile hormone analog, pyriproxyfen, on apterous form of soybean aphid (Aphis glycines). J Appl Entomol 131:297–302
Ross MH, Cochran DG (1990) Response of late-instar Blatella germanica (Dictyoptera: Blatellidae) to dietary insect growth regulators. J Econ Entomol 83(6):2295–2305
Ross MH, Cochran DG (1991) Effects on German cockroach nymphs of contact exposure to IGRs, singly and in combination. Entomol Exp Appl 61(2):117–122
Schneider MI, Smagghe G, Pineda S, Vinuela E (2004) Action of insect growth regulator insecticides and spinosad on life history parameters and absorption in third instar larvae of the endoparasite Hyposoter didymator. Biol Control 31:189–198
Singh S, Kumar K (2011) Effect of the juvenile hormone agonist pyriproxyfen on larval and pupal development of the citrus swallowtail Papilio demoleus (Lepidoptera: Papilionidae). Int J Trop Insect Sci 31(3):192–198
Strey OF, Teel PD, Longnecker MT (2001) Effects of pyriproxyfen on off-host water balance and survival of adult lone star ticks (Acari: Ixodidae). J Med Entomol 38:589–595
Sukontason KL, Bunchoo M, Khantawa B, Piangjai S, Rongsiyam Y, Sukontason K (2007) Comparison between Musca domestica and Chrysomya megacephala as carriers of bacteria in northern Thailand. Southeast Asian J Trop Med Public Health 31(1):38–44
Sulaiman S, Mustafa SHMS, Jeffery J (2008) Effect of Triflumuron and Pyriproxyfen on Musca domestica L. larval stages in laboratory. Iran J Arthropod-Borne Dis 2(1):1–6
Tuncer I, Dik B, Guçlu F, Baysal B, Ozkan F (1996) Ophthalmomyiasis caused by Sarcophaga larvae in child. Acta Parasitol Turc 20(2):229–231
Turk M, Afaşar I, Özbel Y, Sener AG, Üner A, Türker M (2006) A case of nasomyiasis whose agent was Sarcophaga sp. Acta Parasitol Turc 30(4):330–332
Varloud M, Hodgkins E (2015) Five month comparative efficacy evaluation of three ectoparasiticides against adult cal fleas (Ctenocephalides felis), flea egg hatch and emergence, and adult brown dog ticks (Rhiphicephalus sanguineus sensu lato) on dogs housed outdoors. Parasitol Res 114:965–973
Wang HY, Olmstead AW, Li H, LeBlanc GA (2005) The screening of chemicals for juvenoid-related endocrine activity using the water flea Daphnia magna. Aquat Toxicol 74(3):193–204
Wood JP, Godfrey LD (1998) Effects of whitefly insect growth regulators Knack and Applaud on cotton aphid reproduction and survival. In: Dugger P, Ritcher D (eds) Proceedings of Beltwide Cotton Conference, San Diego, CA. National Cotton Council, Memphis, TN, pp 1278–1281
Yazar S, Dik B, Yalcin S, Demirtas F, Yaman O, Ozturk M, Sahin I (2005) Nasocomial oral myiasis by Sarcophaga sp. in Turkey. Yonsei Med J 46(3):431–433
Acknowledgments
The authoress expresses her gratitude to the Council of Scientific and Industrial Research (CSIR), New Delhi, for providing financial assistance (Ref. No. 09/001/(0376)/2013/EMR-I) and University Grants Commission (UGC) New Delhi for funds to purchase juvenoids under Major research Project [F.No. 33-354/2007 (SR)] sanctioned to Prof. Krishna Kumar.
Conflicts of interest
The authors declare that they have no conflict of interest.
Compliance with Ethical Standards
All procedures performed in the study were in accordance with the Guidelines of Institutional Animal Ethical Committee (IAEC) of the Department of Zoology, University of Allahabad.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article

Cite this article
Singh, S., Kumar, K. Effects of juvenoid Pyriproxyfen on reproduction and F1 progeny in myiasis causing flesh fly Sarcophaga ruficornis L. (Sarcophagidae: Diptera). Parasitol Res 114, 2325–2331 (2015). https://doi.org/10.1007/s00436-015-4428-9
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00436-015-4428-9