
Abstract
Knowledge of natural Anguillicola infections of Short-finned eels (Anguilla australis) in New Zealand is very limited. So far, no study contains data on all life cycle stages of Anguillicola novaezelandiae in naturally infected eels. In order to study the frequency of A. novaezelandiae in New Zealand Short-finned eels, we examined eels of the North and the South Island for the presence of the swim bladder parasite. The results show that A. novaezelandiae is a common parasite of the Short-finned eel. The parasite was present in both regions. Eels from both localities show differences in their infection status with respect to prevalence, abundance and intensity. While eels of the South Island were only infected with larval stages, adult and preadult stages could be detected in eels of the North Island. Nevertheless, infrapopulations at both sites were dominated by larval parasite stages. This unique composition of infrapopulations has never been described for any Anguillicola species before and suggests a seasonal occurrence as a possible reason. Export of live eels should be handled cautiously to prevent the spread of A. novaezelandiae throughout other eel populations.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Within the genus of freshwater eels (Anguilla), four species are of major importance for worldwide trade and food production (FAO 2012): the European eel (Anguilla anguilla), the Japanese eel (Anguilla japonica), the American eel (Anguilla rostrata) and the Short-finned eel (Anguilla australis). Adult eels are traded as processed dish, smoked, frozen and also alive. With this export and import of live eels, not only alien fish species travel around the world but with them are a variety of parasites. As the spread of the gill parasites Pseudodactylogyrus bini and Pseudodactylogyrus anguilla and the swim bladder nematode Anguillicola crassus has shown, this trade can lead to the introduction of alien parasite species. A. crassus has spread quickly throughout the European eel population and can cause damage to the eels' swim bladder (Jakob et al. 2009; Kirk 2003; Køie 1991). This swim bladder damage could disable the eel in reaching its spawning ground in the Sargasso Sea (Palstra et al. 2007) and could thus be one factor contributing to the decline of the European eel population (Sures and Knopf 2004).
The family Anguillicolidae consists of five species, which are all adapted to different Pacific eel species. In 2006, Moravec split the genus Anguillicola into the two genera Anguillicola and Anguillicoloides due to morphologic differences. The phylogenetic study of Laetsch et al. (2012) on Anguillicolidae found no support for the maintenance of these two genera; therefore, all species of the family are referred to as Anguillicola in the following.
Whilst the European eel is not an original host to any Anguillicola species, the Japanese eel is the natural host of the two species A. crassus and Anguillicola globiceps; A. novaezelandiae is a parasite of the Short-finned eel. Anguillicola australiensis is parasitic in the Long-finned eel (Anguilla reinhardtii) and Anguillicola papernai is found in Anguilla mossambica (Moravec and Taraschewski 1988). Two of these Anguillicola species have shown their invasive potential and were able to infect alien eel species. Besides A. crassus, which has been in focus of science since live eel trade started in the 1960s (Egusa 1979; Kennedy 2007), A. novaezelandiae has also been able to infect European eels and built a stable population in an Italian lake (Moravec et al. 1994; Paggi et al. 1982). However, the latter species disappeared from the lake after the arrival of A. crassus, probably due to hybridization between A. crassus and A. novaezelandiae (Grabner et al. 2012). Apart from eels which are used as definitive hosts, all Anguillicola species use mainly copepods as obligate intermediate hosts. In the eel host, larval stages are found in the swim bladder wall as third stage (L3) and fourth stage larvae (L4) while preadult stages migrate into the swim bladder lumen, feed on the host's blood and moult to the adult stage (Moravec 2006).
Anguillicola novaezelandiae is native to Australia and New Zealand, where its host, the Short-finned eel (A. australis), occurs. So far, there is only limited data on the distribution of the parasite in Short-finned eel populations (Hine 1978; Lefebvre et al. 2004; Rid 1973). The latest work of Lefebvre et al. (2004) concentrates solely on adult parasites in the lumen of the swim bladder. Older studies mainly gave prevalence data or only report if eels were infected or not. So far, no study has presented data on all stages of the parasite. In order to evaluate the potential risk of spreading A. novaezelandiae with the trade of live Short-finned eels, its occurrence in the original host (including all larval stages within the swim bladder wall) has to be analysed. Therefore, eels from two sampling sites, located on each of the New Zealand main islands, have been checked for the presence of A. novaezelandiae in the present study. In addition, we compared the demographic structure of A. novaezelandiae in the present study with that reported in other Anguillicola–Anguilla systems, in order to infer potential differences in seasonal patterns and/or population dynamics.
Material and methods
Eel sampling
Wild Short-finned eels (A. australis) were obtained from professional fishermen in November and December 2007 and transported alive to the lab. Sampling sites are located on each of the New Zealand islands. The Hydro Lakes of the Waikato River were sampled on the North Island. On the South Island, eels were sampled from Lake Ellesmere (Table 1).
The eels were kept in large oxygenated tanks, killed by decapitation within 2 weeks after capture and examined immediately. The length and weight of the eels were measured, whereupon the swim bladder was removed and examined for the presence of A. novaezelandiae. The condition factor (C factor) was calculated as described by Schäperclaus (1990), with C as the ratio of the fish somatic mass × 100 × total length−3.
Parasitological examination of eel swim bladders
All parasites were removed from the swim bladder lumen. They were identified and their number and sex were recorded. Species identification was performed morphologically according to the key of Moravec (2006) and subsequently verified by molecular analyses. Preadult and adult parasites were stored in 70 % alcohol. After removal of adult worms, the swim bladder was examined between two plexiglass plates with a stereomicroscope (magnification ×8 to ×50) for larval stages (L3 and L4). Since the differentiation between L3 and L4 stage is not always possible, all larvae longer than 1.5 mm were considered as L4, as described in Blanc et al. (1992). Length and width of preadult and adult parasites were measured using the stereomicroscope (magnification ×8 to ×50). Prevalence (P) and mean intensity of parasites in the eels were calculated as described in Bush et al. (1997). For the description of the host–parasite relation, a frequency distribution was used. In order to check for an aggregated pattern of the parasite's population, the variance-to-mean ratio was calculated as described in Shaw and Dobson (1995; k = s 2 / MA; where MA is mean abundance). Overdispersion (negative binominal distribution) is characterised by k > 1, while a random or Poisson distribution is characterised by k ≈ 1 (Shaw and Dobson 1995).
Molecular identification of A. novaezelandiae
For genetic verification of the morphologically identified species, a subsample of 11 nematodes was analysed genetically. DNA was extracted from six randomly chosen parasites and the five largest specimens using the JETQUICK Genomic DNA Purification Kit (Genomed) based on the method of Bowtell (1987). A part of the coxI gene was amplified using the primers HCO2198 and LCO1490 (Folmer et al. 1994) and PCR conditions as described in Wielgoss et al. (2008). PCR products were purified with a JETQUICK PCR Product Purification Kit (Genomed) and sent for sequencing (GATC) with HCO2198 and LCO1490 primers. A BLAST search, implemented in the homepage of the National Center for Biotechnology Information (http://blast.ncbi.nlm.nih.gov/Blast.cgi), was conducted with the sequences obtained to check for homologies in the database.
Statistical treatment of data
For comparisons of eel length, weight and C factor, the unpaired Student's t test was used with a significance level of p < 0.05 and p < 0.001, respectively. Dependence of infection status or number of parasites on the C factor was examined by one-way analysis of variance. All statistical tests were conducted with STATISTICA 10.
Results
Eel data
Data on the Short-finned eels, their origin, length, weight and condition factor are summarised in Table 1. Eels from the North Island were longer and heavier than eels from the South Island. The weight of both eel samples shows a highly significant difference, while the length shows no significant difference. Infection status or number of parasites had no influence on the C factor (p < 0.05).
Infection patterns of A. novaezelandiae
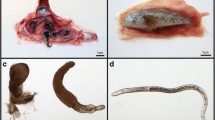
The 105 eels from the North Island were infected with 145 Anguillicola individuals, whereas the 101 South Island eels harboured 675 nematodes. All nematodes detected in the swim bladder were identified as A. novaezelandiae according to the key of Moravec (2006). The nematode infrapopulation in eels from South Island was dominated by L3 (79.7 %), while the infrapopulation in eels from North Island was dominated by L4 (53.1 %; Fig. 1). Eels of the South Island harboured only larval stages with a prevalence of 89.1 %. The total prevalence in North Island eels was 51.4 % of which 21.0 % of the eels were found to be infected with preadult and adult stages (Table 2).

Infrapopulation of A. novaezelandiae developmental stages. Eels from the North Island were infected with all stages of A. novaezelandiae, while eels originating from the South Island were only infected with larval parasite stages in the swim bladder wall
Only two eels of the North Island were infected with mature parasites. While one eel was infected with a single male parasite, the other eel contained three male and two female parasites. The swim bladder of the latter eel also contained second stage larvae (L2). Only one encapsulated larva could be observed in the swim bladder wall. No swim bladder showed signs of pathological changes due to the infection with Anguillicola. Frequency scale of infection showed negative binominal distribution for parasites from North Island eels. Of the eels from the South Island, 33.6 % were infected with more than seven parasites (Fig. 2). The variance-to-mean ratio for these data was computed for both sampling sites (s 2/MA). Both were >1, indicating overdispersion (Table 3).

Frequency distribution of A. novaezelandiae. Data on the North Island (dark) show a negative binominal distribution. On the South Island (light), 33.6 % of the eels were infected with more than seven parasites
Morphometric measurements of preadult and adult stages show two size classes (Fig. 3). The first group (n = 25) had a mean length of 4.58 ± 0.85 mm, while the second group (n = 4) was 28.69 ± 12.63 mm long. The longest parasite was a 40.16-mm-long male with a width of 1.992 mm. The two female parasites of the same eel could not be measured as they were opened immediately after removal from the swim bladder to obtain eggs for experimental infections described elsewhere (Grabner et al. 2012).

Size classes of parasites in swim bladder lumen. Preadult parasites show a very small size range (4.58 ± 0.85 mm), while the length of adult parasites varied between 12.0 and 40.2 mm (mean length 28.69 ± 12.63 mm)
Molecular species identification
The sequences obtained from the 11 specimens used for molecular analysis (length 597 bp) were identical except for one polymorphic position at 119 bp. New sequence information was deposited in GenBank under accession nos. JX868555 and JX868556. A comparison to the sequences of A. novaezelandiae from Laetsch et al. (2012) (originating from Tasmania, Australia) using BLAST shows only 89.1 % identity.
Greatest consensus showed the sequence of A. australiensis with 90.2 % identity. A. crassus showed an identity of 89.3 %.
Discussion
According to morphological identification, all parasites found in the swim bladder belong to the species A. novaezelandiae. The cox1 sequences identified showed genetic difference to all other Anguillicola cox1 genes. As Laetsch et al. (2012) mentioned, the genetic difference of the cox1 gene between A. novaezelandiae from New Zealand and Tasmania suggests the hypothesis of the existence of two sibling species. The geographic distance between New Zealand and Australia could have lead to isolation of both A. novaezelandiae populations and thus to the genesis of sibling species. This seems plausible since the taxonomic status of the host species A. australis is still discussed and it is currently uncertain if the species may consist of the two subspecies A. australis schmidtii (New Zealand) and A. australis australis (Australia and Tasmania). The latest studies found both morphological (Watanabe et al. 2006) and genetic differences (Shen and Tzeng 2007) comparing A. australis from New Zealand and Australia. Therefore, it is possible that both eel subspecies are infected by different A. novaezelandiae sibling species. Further studies combining morphological and genetic approaches should be performed to verify the taxonomic status of the two parasite populations.
The finding of only one Anguillicola species is in accordance with several findings of A. novaezelandiae in the 1970s (Hine 1978; Moravec and Taraschewski 1988; Rid 1973) as well as with the latest study of Lefebvre et al. (2004). So far, A. novaezelandiae is the only known species of Anguillicola in New Zealand as well as in A. australis. However, three of the four male adult specimens in our study were larger than all A. novaezelandiae specimens described so far (see Table 4). This might reflect the lack of data on A. novaezelandiae in literature (Lefebvre et al. 2004; Moravec and Rohde 1992; Moravec and Taraschewski 1988; Paggi et al. 1982). Further studies are needed to gain more knowledge of the species in terms of their morphology, life cycle and distribution.
This study is the first to cover all stages of A. novaezelandiae in eel. By including larval stages in the swim bladder wall, the parasite could also be found in eels of the South Island. A study only covering stages in the swim bladder lumen would have detected no parasites, even though 89 % of the eels were infected. Having this in mind, a lack of A. novaezelandiae findings in the literature (as described for some locations by Hine (1978) and Lefebvre et al. (2004)) must be viewed with caution. A possible explanation for the differences in infrapopulation composition of the two sampling sites could be seasonality. While eels from the South Island harboured only larval stages, 14 % of the eels from the North Island harboured preadults. The preadult stages were all about of the same size (4.6 ± 0.9 mm). Only two eels from the North Island were infected with adult stages, one with a single worm and the other eel with adult stages exceeding all so far known size data for A. novaezelandiae. A possible explanation is that these adults overwintered in the eel, while all other eels were freshly infected in spring 2007. The hypothesis of seasonality is supported by the fact that 20 % of South Island A. novaezelandiae are in L4 stage and 23 % of North Island parasites are in the preadult stage. Parasites of the North Island seem to be a step ahead in their life cycle. An explanation is the about 2 °C warmer temperature on the northern sampling site compared to the South Island sampling site (NIWA 2012). Prevalence data of preadult and adult stages of A. novaezelandiae by Lefebvre et al. (2004) also support this hypothesis. Prevalence in October was lower than in December and eels from the North Island were infected with a higher prevalence at the same sampling date than those from the South Island. So far, indications for seasonality have not been described for any other Anguillicola–Anguilla system. Compared to data from Australia (A. australiensis from A. reinhardtii; Kennedy 1994) and Taiwan (A. crassus from A. japonica; Münderle et al. 2006), clear differences become obvious for A. novaezelandiae from New Zealand. While only one dead larva was found during our study, a large number of dead larvae were described in Kennedy (1994) and Münderle et al. (2006). Furthermore, the dominance of larval stages in the eel A. novaezelandiae infrapopulation in both locations of the present study is a unique finding in an Anguillicola–Anguilla system (see Fig. 4); as in Australia and Taiwan, the infrapopulations were all dominated by worms in the swim bladder lumen (Kennedy 1994; Münderle et al. 2006). For A. crassus, lower prevalences in wild and cultured Japanese eel populations were described in winter (Egusa et al. 1969; Han et al. 2008; Kim et al. 1989; Münderle et al. 2006; Nagasawa et al. 1994), but a lack of adult stages in spring was never observed in neither the natural nor the newly invaded distribution areas. All these data support the hypothesis of seasonal occurrence of A. novaezelandiae in New Zealand; nevertheless, more eel sampling should be carried out to verify this theory as other factors like different water temperature regimes of the sampling sites may also be important.

Comparison of Anguillicola infrapopulations in native hosts. While about 30 % of the Anguillicola infrapopulations in Australia and Taiwan consist of adult parasites, only eels of one location in New Zealand were infected with adult parasites (4 %), whereas the second sampling site lacks any adult A. novaezelandiae. Only living stages were taken into account to allow a comparison between the different localities; all data are of spring samples. Australia: data from the sampling on 1 December 1993 at Bundaberg were used as an example (details see Kennedy 1994; Kennedy personal communication 2012); Taiwan: data from Kao-Ping River sampled in March 2003 were used as an example (worms < 10 mm were counted as preadult; for details, see Münderle 2005 and Münderle et al. 2006). Numbers indicate the number of infected eels per sampled eel
The frequency distribution of all stages and both sampling sites exhibited an aggregated dispersal. This is in accordance with other natural Anguillicola–Anguilla systems (Han et al. 2008; Kennedy 1994; Kim et al. 1989; Münderle et al. 2006). The observed overdispersion is a characteristic of macroparasites in wildlife populations and arises from heterogeneities in exposure of host populations or under infection pressure (Shaw and Dobson 1995). Eels of the South Island showed a more aggregated infection pattern than eels of the North Island. The rate of infection with parasites in the swim bladder lumen in this study is comparable to literature data of Rid (1973), Hine (1978) and Lefebvre et al. (2004). Prevalence of A. novaezelandiae in the swim bladder lumen in the natural habitat never exceeded 12 % (Boustead (1982) mentioned that eel in Hine (1978) originating from Rangitaiki River, Bay of Plenty, with P of 72.7 % was of an eel farm; also, Long-finned and Short-finned eel infection is combined in the paper). The infection of Short-finned eel with A. novaezelandiae is similar to infections of other Anguillicolidae in their natural eel host in wild habitats. Prevalence of adult A. papernai in A. mossambica varies between 8 and 63 % (Taraschewski et al. 2005). Kennedy (1994) found 27–78 % of A. reinhardtii infected with adult and preadult A. australiensis. Japanese eel populations showed a maximum prevalence of around 60 % for A. crassus (all stages; Han et al. 2008; Münderle et al. 2006), while A. globiceps infected the same eel species with a prevalence of 6 to 60 % (all stages; Nagasawa et al. 1994; Suyehiro 1957; Wang and Zhao 1980; Wu 1956). Infection of non-native eel species is more severe. A. crassus was introduced to Europe in the 1980s and to North America in the 1990s. The alien parasite infects its new host, the European eel with a prevalence of up to 100 % (overview in Jakob et al. 2009). In Germany, a well-studied country, prevalence is around 80 % and mean intensity varies between 1 and 14 parasites per eel (Jakob et al. 2009; Sures and Streit 2001; Sures et al. 1999; Würtz et al. 1998).
The findings in the present study seem to be comparable with other natural Anguillicola–Anguilla systems. No swim bladder damage could be observed. The invasive potential, shown by the introduction of the species to Europe in the 1970s, should be reason enough for further studies of the species in its natural habitat. As the populations of the most important eel species have fallen to catastrophic lows (Stone 2003), Short-finned eels may be of growing importance in eel production worldwide. There is no glass eel fishery for this species, so live eels are exported from New Zealand as yellow eels with their parasite load. Wang et al. (2006) showed the import of a number of parasite species via live trade of Short-finned eels to Taiwan. In the future, more attention should be paid to the risk of this trade. In order to give a risk assessment, the distribution of A. novaezelandiae throughout the Short-finned eel population should be studied including larval stages. In order to study the seasonal occurrence, some eel populations should be monitored regularly throughout a year. The knowledge of a distribution pattern and the seasonal occurrence of the species are essential to give advice to eel traders and prevent the parasite from spreading throughout alien eel species.
References
Blanc G, Bonneau S, Biaganti S, Petter AJ (1992) Description of the larval stages of Anguillicola crassus (Nematoda, Dracunculoidea) using light and scanning electron microscopy. Aquat Living Resour 5:307–318
Boustead NC (1982) Fish diseases recorded in New Zealand, with a discussion on potential sources and certification procedures. In: Fisheries Research Division occasional publication, 34th ed. New Zealand Ministry of Agriculture and Fisheries, Wellington, pp. 18
Bowtell DDL (1987) Rapid isolation of eukaryotic DNA. Anal Biochem 162:463–465
Bush AO, Lafferty KD, Lotz JM, Shostak AW (1997) Parasitology meets ecology on its own terms: Margolis et al. revisited. J Parasitol 83:575–583
Egusa S (1979) Notes on the culture of European eel (Anguilla anguilla) in Japanese eel farming ponds. Rapp P-V Réun Cons Int Explor Mer 174:51–58
Egusa S, Kira K, Wakabayashi H (1969) On the occurrence of Anguillicola globiceps Yamaguti, a swimbladder roundworm, in pond-cultured eels. Fish Pathol 4:52–58
FAO (Food and Agriculture Organization of the United Nations) (2012) Total fish production 1950–2010. FishStat Plus—universal software for fishery statistical time series: http://www.fao.org/fishery/statistics/software/fishstat/en. Accessed May 2012
Folmer O, Black M, Hoeh W, Lutz R, Vrijenhoek R (1994) DNA primers for amplification of mitochondrial cytochrome c oxidase subunit I from diverse metazoan invertebrates. Mol Mar Biol Biotechnol 3:294–299
Grabner DS, Dangel KC, Sures B (2012) Merging species? Evidence for hybridization between the eel parasites Anguillicola crassus and A. novaezelandiae (Nematoda, Anguillicoloidea). Parasit Vectors 5:244
Han YS, Chang YT, Taraschewski H, Chang SL, Chen CC, Tzeng WN (2008) The swimbladder parasite Anguillicola crassus in native Japanese eels and exotic American eels in Taiwan. Zool Stud 47:667–675
Hine PM (1978) Distribution of some parasites of freshwater eels in New Zealand. New Zeal J Mar Fresh 12:179–187
Jakob E, Walter T, Hanel R (2009) A checklist of the protozoan and metazoan parasites of European eel (Anguilla anguilla): checklist of Anguilla anguilla parasites. J Appl Ichthyol. doi:10.1111/j.1439-0426.2009.01345.x
Kennedy CR (1994) The distribution and abundance of the nematode Anguillicola australiensis in eels Anguilla reinhardtii in Queensland, Australia. Folia Parasitol 41:279–285
Kennedy CR (2007) The pathogenic helminth parasites of eels. J Fish Dis 30:319–334
Kim Y-G, Kim E-B, Kim J-Y, Chun S-K (1989) Studies on a nematode, Anguillicola crassa parasitic in the air bladder of the eel. J Fish Pathol 2:1–18
Kirk RS (2003) The impact of Anguillicola crassus on European eels. Fisheries Manag Ecol 10:385–394
Køie M (1991) Swimbladder nematodes (Anguillicola spp.) and gill monogeneans (Pseudodactylogyrus spp.) parasitic on the European eel (Anguilla anguilla). ICES J Mar Sci 47:391–398
Laetsch DR, Heitlinger EG, Taraschewski H, Nadler SA, Blaxter ML (2012) The phylogenetics of Anguillicolidae (Nematoda: Anguillicolidea), swimbladder parasites of eels. BMC Evol Biol 12:60
Lefebvre F, Schuster T, Münderle M, Hine M, Poulin R (2004) Anguillicolosis in the short-finned eel Anguilla australis: epidemiology and pathogenicity. New Zeal J Mar Fresh 38:577–583
Moravec F (2006) Dracunculoid and Anguillicoloid nematodes parasitic in vertebrates, 1st edn. Academia, Praha, 636
Moravec F, Rohde K (1992) Three species of nematodes of the superfamily Dracunculoidea from Australian fishes. Acta Soc Zool Bohemoslov 56:187–195
Moravec F, Taraschewski H (1988) Revision of the genus Anguillicola Yamaguti, 1935 (Nematoda: Anguillicolidae) of the swimbladder of eels, including descriptions of two new species, A. novaezelandiae sp. n. and A. papernai sp. n. Folia Parasitol 35:125–146
Moravec F, Di Cave D, Orecchia P, Paggi L (1994) Present occurrence of Anguillicola novaezelandiae (Nematoda, Dracunculoidea) in Europe and its development in the intermediate host. Folia Parasitol 41:203–208
Münderle M (2005) Ökologische, morphometrische und genetische Untersuchungen an Populationen des invasiven Schwimmblasen-Nematoden Anguillicola crassus aus Europa und Taiwan. Dissertation, Fakultät für Chemie und Biowissenschaften der Universität Karlsruhe (TH)
Münderle M, Taraschewski H, Klar B, Chang CW, Shiao JC, Shen KN, He JT, Lin SH, Tzeng WN (2006) Occurrence of Anguillicola crassus (Nematoda: Dracunculoidea) in Japanese eels Anguilla japonica from a river and an aquaculture unit in SW Taiwan. Dis Aquat Org 71:101–108
Nagasawa K, Kim YG, Hirose H (1994) Anguillicola crassus and A. globiceps (Nematoda, Dracunculoidea) parasitic in the swimbladder of eels (Anguilla japonica and A. anguilla) in East Asia—a review. Folia Parasitol 41:127–137
NIWA (National Institute of Water and Atmospheric Research) (2012) The National Climate Database. http://cliflo.niwa.co.nz/. Accessed 26 Apr 2012
Paggi L, Orecchia P, Minervini R, Mattiucci S (1982) Appearance of Anguillicola australiensis Johnston and Mawson, 1940 (Dracuncoloidea: Anguillicolidae) in Anguilla anguilla of Lake Bracciano. Parassitologia 24:139–144
Palstra AP, Heppener DFM, van Ginneken VJT, Székely C, van Den Thillart GEEJ (2007) Swimming performance of silver eels is severely impaired by the swim-bladder parasite Anguillicola crassus. J Exp Mar Biol Ecol 352:244–256
Rid LE (1973) Helminth parasites of the long-finned eel, Anguilla dieffenbachii, and the short-finned eel, A. australis. Mauri Ora 1:99–106
Schäperclaus W (1990) Fischkrankheiten. Akademie Verlag, Berlin
Shaw DJ, Dobson AP (1995) Patterns of macroparasite abundance and aggregation in wildlife populations: a quantitative review. Parasitology 111(Suppl):S111–27
Shen KN, Tzeng WN (2007) Genetic differentiation among populations of the shortfinned eel Anguilla australis from East Australia and New Zealand. J Fish Biol 70:177–190. doi:10.1111/j.1095-8649.2007.01399.x
Stone R (2003) Freshwater eels are slip-sliding away. Science 302:221–222
Sures B, Knopf K (2004) Parasites as a threat to freshwater eels? Science 304:208–209
Sures B, Streit B (2001) Eel parasite diversity and intermediate host abundance in the River Rhine, Germany. Parasitology 123:185–191
Sures B, Knopf K, Würtz J, Hirt J (1999) Richness and diversity of parasite communities in European eels Anguilla anguilla of the River Rhine, Germany, with special reference to helminth parasites. Parasitology 119:323–330
Suyehiro Y (1957) On the ecology of parasites of Japanese eel. In: Suyehiro Y, Oshima Y, Hiyama Y (eds) Suisangaku Shusei. Tokyo University Press, Tokyo, pp 415–418
Taraschewski H, Boomker J, Knopf K, Moravec F (2005) Anguillicola papernai (Nematoda: Anguillicolidae) and other helminths parasitizing the African longfin eel Anguilla mossambica. Dis Aquat Org 63:185–195
Wang P, Zhao Y (1980) Observations on the life history of Anguillicola globiceps (Nematoda: Anguillicolidae). Acta Zoologica Sinica 26:243–249
Wang WS, Hung S-W, Lin Y-H, Shih C-W, Tu C-Y, Tung K-C, Chen B-R, Ooi H-K (2006) Parasitosis of Short-finned eel, Anguilla australis schmidti, in Taiwan. Bull Eur Ass Fish Pathol 26:106–111
Watanabe S, Aoyama J, Tsukamoto K (2006) Confirmation of morphological differences between Anguilla australis australis and A. australis schmidtii. New Zeal J Mar Fresh 40:325–331
Wielgoss S, Taraschewski H, Meyer A, Wirth T (2008) Population structure of the parasitic nematode Anguillicola crassus, an invader of declining North Atlantic eel stocks. Mol Ecol 17:3478–3495. doi:10.1111/j.1365-294X.2008.03855.x
Wu H-S (1956) Studies on the parasitic nematodes of freshwater fishes in China 1. Acta Hydrobiologica Sinica 1:99–106
Würtz J, Knopf K, Taraschewski H (1998) Distribution and prevalence of Anguillicola crassus (Nematoda) in eels Anguilla anguilla of the rivers Rhine and Naab, Germany. Dis Aquat Org 32:137–143
Acknowledgments
We are grateful to R. Poulin (University of Otago) for providing eels and laboratory space and to K. Herrmann for laboratory assistance. We would like to thank C. Kennedy (University of Exeter) for providing detailed data on A. australiensis in A. reinhardtii. For their help with molecular identification of parasites, we would like to thank S. Mattiucci (Sapienza, University of Rome), M. Paoletti (Tuscia University) and D. Grabner (University of Duisburg-Essen). This work was supported by the German Academic Exchange Service DAAD (K.C.D., grant number D/07/48024) and by Deutsche Forschungsgemeinschaft via grant SU217/12-1.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Dangel, K.C., Sures, B. Natural Anguillicola novaezelandiae infection—is there seasonality in New Zealand?. Parasitol Res 112, 1623–1630 (2013). https://doi.org/10.1007/s00436-013-3317-3
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00436-013-3317-3