
Abstract
The immune responses of Angiostrongylus cantonensis infection are closely relevant to the host’s self-protection and the nematode’s pathogenesis. In the present study, BALB/c mice were randomly divided into uninfected control group, infection group 1, and infection group 2. The infection group 1 and infection group 2 were infected with 20 and 40 third-stage larvae of A. cantonensis per mouse, respectively. The splenocytes from the mice were collected and cultured on the 19th and 25th days post-infection; the subtypes of T cells in splenocytes were detected by flow cytometry with fluorescence staining method, and the cytokines in cultured supernatants of splenocytes were assayed by the method of ELISA. The specific IgG and IgE antibodies in sera of the mice were periodically detected by ELISA. The results showed that the percentages of CD4+ and CD4+ IL-4+ T cells in splenocytes of infected mice were much higher (P < 0.05) than those in control mice; however, the percentages of CD4+ IL-17+ and CD4+ IFN-γ+ T cell were much lower(P < 0.01) after the infection. The levels of CD8+ T cells in infected mice also rose, but differences between control mice and infected mice were not significant. In comparison with control mice, the concentration of IL-4 in the cultured supernatants of splenocytes in infected mice increased significantly (P < 0.05), but that of IL-17 decreased significantly (P < 0.01). In addition, the number of larvae infected and days after infection may influence levels of the T cell subtypes and the cytokines in spleen, too (P > 0.05). On humoral immunity, the levels of specific IgG antibodies in sera rose a bit at the fifth day post-infection, and reached a peak at the 20th day post-infection; the specific IgE antibodies gradually heightened during first 10 days post-infection; then, it showed a downward trend during the 15th to 25th days post-infection. It is evident that the percentages of CD4+ T lymphocytes of spleen in the mice infected with A. cantonensis markedly increase and polarize to Th2 phenotypes, and the function of Th17 cells is inhibited. In addition, the elevation of specific IgG antibodies in sera of the infected mice is more significant than that of specific IgE antibodies.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Angiostrongylus cantonensis (A. cantonensis) is the pathogen of angiostrongyliasis, an emerging infection disease, which mainly invades human central nervous system and cause eosinophilic meningoencephalitis (Hüttemann et al. 2007). In recent years, several outbreaks of human angiostrongyliasis have been reported in China and other countries (Wang et al. 2008; Qu et al. 2007; Lindo et al. 2002; Slom et al. 2002; Ko et al. 1987), and its importance has been recognized. Previous studies (Sugaya et al. 1997; Perez et al. 1989) showed that the immune responses of the hosts infected with A. cantonensis were very important for the host’s resistance to the infection and pathogenesis of the parasitic nematode (OuYang et al. 2012; Wei et al. 2012). Therefore, the alteration of T cell subtypes in spleen and antibodies of serum in mice infected by A. cantonensis were observed in this study.
Materials and methods
Mice
Female BALB/c mice aged 6 weeks, SPF grade, were purchased from Experimental Animal Center, Guangdong Province.
Reagents
PMA, Saponing, Ionomycin, and Brefeldin A (BFA) were produced by Sigma Company, USA. PerCP anti-mouse CD4, FITC anti-mouse CD8, APC anti-mouse IL-4, APC anti-mouse IFN-gamma, and PE anti-mouse IL-17 were purchased from BD Biosciences Inc. USA. IL-4 (DY404) and IL-17 (DY421) ELISA assay kit were purchased from R&D Systems Inc., USA.
Instruments
Flow cytometry (Calibur) from BD Biosciences Inc., EL × 800 microplate reader from BioTeK Inc., 1267P CO2 incubator manufactured by Yi Liang medical company, Shanghai, China.
Methods
Animal infection
The third-stage larvae of A. cantonensis were obtained from snail Achatina fulica (Chen et al. 2011). Thirty BALB/c mice were randomly divided into control group, infection group 1, and infection group 2; each group has ten mice. The infection group 1 was infected with 20 larvae per mouse by peritoneal injection, infection group 2 infected with 40 larvae, and control group received only water.
Confirmation of infection
The activities of infected mice were observed during the experiment. The larvae of A. cantonensis in the brain of infected mice were examined by microscopic or visual study. The pathological changes of the brain tissues were detected by paraffin section and HE staining.
Collection of sera
The blood was periodically collected from the caudal vein or eyeball of the infected mice at pre-infection and post-infection, and the sera were stored at −20 °C.
Collection of splenic lymphocytes
Five mice in each group were sacrificed at the 19th and 25th days post-infection. Splenic lymphocytes of the mice were collected as described in the report (Anukumar and Shahir 2011). The cells were resuspended in 4 mL RPMI Medium 1640 (1×) liquid, stained by 0.4 % trypan blue and counted under microscope.
Detection of T cell subtypes in splenocytes
According to the methods reported by other authors (Anukumar and Shahir 2011; Yang et al. 2006), spleen lymphocytes(2 × 106 per milliliters) were incubated with PMA 10 ng/mL and Ionomycin 1 μg/mL in 37 °C, 5 % CO2 incubator for 1 h and then incubated with BFA for 4 h. The cells were fixed by 4 % formaldehyde and added buffer (including Saponin) at 4 °C overnight. The mAbs labeled fluorescein were mixed with the cells; the subtypes of T cells in splenocytes were detected by flow cytometry.
Detection of cytokines in culture supernatants of splenocytes
The splenocytes were incubated with PMA and Ionomycin in the cell culture plates for 72 h in 37 °C, 5 % CO2 incubator; each splenocyte suspension was simultaneously cultured with three well. The concentrations of IL-4 and IL-17 in culture supernatants of splenocytes were assayed by ELISA according to the protocol in kit. The optical density (OD) values were taken the average detected value of the three well; then, OD values were converted into concentration of the cytokine in accordance with the standard curve.
Preparation of the adult antigens
The preparation of adult A. cantonensis antigens for ELISA is referred to the report of Jin (Jin et al. 2006).
Assay of specific IgG antibodies in sera
Microtiter plates were coated with the adult A. cantonensis antigens (10 μg/mL, 100 μL per well). The dilution of the mouse sera tested was 1:25, and that of the anti-mouse IgG-HRP antibody (Jackson ImmunoResearch Inc., USA) was 1:5,000 in ELISA assay. The optical density (OD) value was read at 450 nm with a microplate reader.
Assay of specific IgE antibodies in sera
The dilution of anti-mouse IgE-HRP antibody (Alpha Diagnoestic International Inc., USA) was 1:4,000, and the other methods were similar to the detection of specific IgG antibodies.
Statistical analysis
All data were expressed as mean ± SD. The comparisons between infection groups and control group were made with SPSS, version 19.0 (SPSS Inc., Chicago, IL, USA), and statistical methods used in data analysis include analysis of variance, t test, and Wilcoxon rank sum test.
Results
Flow cytometry of spleen T cells
The percentages of spleen T cells from the control group and infection groups in flow cytometry were shown in Table 1.
CD4+ and CD8+ T cells
The changes of CD4+ or CD8+ T cell ratios after infection were shown in Fig. 1.

Percentage of CD4+ T and CD8+ T cells in spleen. (*P < 0.05)
T cell subtypes in splenocytes
The changes of T cell subtype ratios after infection were shown in Fig. 2.

Percentage of IL-4+ T, IL-17+ T and IFN-γ+ T cells in spleen. (*P < 0.05; **P < 0.01)
Cytokines in culture supernatants of splenocytes
The detective results of cytokines IL-4 and IL-17 in culture supernatants of splenocytes from the control mice and infected mice were shown in Table 2 and Fig. 3.

Cytokines IL-4 and IL-17 in culture supernatants of splenocytes
Alteration of T cells and cytokines in different times after infection
Flow cytometry analysis
As shown in Table 3, the percentages of CD4+ T, CD4+ IL-4+ T, CD4+ IL-17+ T, and CD4+ IFN-γ+ T cells in the 19th and 25th days post-infection (P > 0.05).
Detection by ELISA
As shown in Table 3, the levels of IL-4 and IL-17 in cultured supernatants of splenocytes in the 19th and 25th days post-infection (P > 0.05).
Dynamics of specific antibodies in serum
IgG antibody
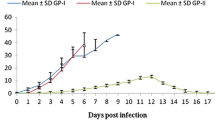
The results were shown in Fig. 4. The levels of specific IgG antibodies in sera of infected mice rose a bit at the fifth day post-infection, the levels of the antibodies reached a peak at the 20th day post-infection.

Detection of the specific IgG antibodies in sera
IgE antibody
The results were shown in Fig. 5. The level of specific IgE antibodies in sera of infected mice arrived at a peak during the 10th to 15th days post-infection; then, it showed a downward trend during 15th to 25th days post-infection.

Detection of the specific IgE antibodies in sera
Confirmation of infection
Some symptoms being relative to central nervous system were damaged, e.g., spin, paralysis had begun to appear in the infected mice since 16th day post-infection. The lesions of hyperemia, hemorrhage, and tissue adhesion on surface of brain tissue could be seen in the infected mice. The pathological changes of the brain tissue were shown in Fig. 6. The larvae of A. cantonensis could be obtained in the brain of all infected mice.

Pathological section of brain tissue from the infected mice (arrows indicate cross section of A. cantonensis)
Discussion
T cells (T lymphocytes) are divided into CD4+ and CD8+ T cells. The biological function of CD4+ T cells is to stimulate and modulate the host immune response, and CD8+ T cells can specifically kill target cells. In this study, the results of flow cytometry showed that percentages of CD4+ T cell in splenocytes of infected mice were significantly higher than those in control mice. In addition, the levels of CD8+ T cells in infected mice increased, but differences between control mice and infected mice were not significant. It indicates that CD4+ T cells may play a major role in immune response of the early-phase infection. The results are consistent with other reports (Lee et al. 1996; Aoki et al. 1998).
CD4+ T cells (or helper T cells, Th) mainly consist of Th1 and Th2 cells, both usually secrete different cytokines to have mutual antagonism in immune response. Therefore, the cytokines from dominant Th cell type will determine the development and outcome of infection (Mosmann et al. 1986; Abbas et al. 1996; Chen 1998; Chen and Wang 2009). IL-4 is produced by CD4+ IL-4+ T cells (belong to Th2); its main biological functions are to proliferate the B cell, to induce antibody types transition to IgE, and to promote the proliferation and function of Th2 cells by autocrine effect. The results of flow cytometry showed that percentages of CD4+ IL-4+ T cells of infected mice were significantly higher than those of the control mice, and levels of IL-4 in cultured supernatants of splenocytes were also significantly elevated after the infection. It is approximately consistent with results of the previous research (Sugaya et al. 1997).
Cytokine IL-17 is mainly generated by Th17 cell belonging to CD4+ T cell (Chen 1998). IL-17, involved in inflammation and vascular generation, was paid more attention in recent years (Kolls and Linden 2004), but its changes in A. cantonensis infection were not known yet. In this study, the percentage of spleen CD4+ IL-17+ T cells and concentration of IL-17 in cultured supernatants of splenocytes from the infected mice were significantly lower than those of the control mice. Another research (Harrington et al. 2005) showed that IL-4, which was high expression after schistosome infection, could inhibit the differentiation of Th17 cells and IL-17 secretion by the PKC-θ signal pathways. Whether IL-4, which was also high expression after A. cantonensis infection, restricts the mature of Th17 and production of IL-17 of the infected mice requires further investigation.
IFN-γ, produced by CD4+ IFN-γ+ T cell being Th1, chiefly takes effects on killing intracellular parasitic protozoan in the infection of parasites (Kushawaha et al. 2011; Langermans et al. 1992; Su and Stevenson 2000). The results of this study showed that the ratio of CD4+ IFN-γ+ T cells in the infected mice was significantly lower than that of the control mice. The reason may be due to significant increasing of CD4+ IL-4+ T cells and IL-4 levels in the mice infected with A. cantonensis, because Lee’s study (Lee et al. 1996) showed that IL-4 could suppress the differentiation and proliferation of Th1 cells by inhibiting the expression of STAT1.
The experimental results showed that levels of the T cell subtypes and cytokines in spleen between infection group 1 and infection group 2 were different, and there were also differences between 19th and 25th days post-infection, but the differences were not statistically significant. It suggests that number of larvae given and infective time might finitely affect the immune responses of the infected mice.
IgG, which is the primary antibodies in host’s immune response, plays a key role in the process of anti-infection immunity. At present, the information on dynamics of specific IgG antibodies in serum after A. cantonensis infection are mostly from the studies of infected rat as animal model (Jin et al. 2006; Huang et al. 2001; Pan et al. 2000). Rats and mice are discrepant in compatibility to A. cantonensis, but human being and mice seem to be same in this aspect. Therefore, the immunological observation to the infected mice as animal model may be more valuable in medicine. However, in our research, the dynamics of specific IgG antibodies in the infected mice was basically similar to the results in rats.
IgE (its Fc fragment) binding the FcεR I receptor in surface of eosinophil will lead to eosinophil degranulation and cause a series of biological effects. Some clinical studies (Dorta-Contreras et al. 2005; Padilla-Docal et al. 2008) found that the cerebrospinal fluids and sera of meningoencephalitis patients infected by A. cantonensis contained a high-level IgE. Meanwhile, the researches about Schistosoma mansoni (Gounni et al. 1994; Joseph et al. 1983; Dunne et al. 1992; Pinot de Moira et al. 2010) and Trichinella spiralis (Watanabe et al. 2005; Gurish et al. 2004) showed that high levels of IgE in the hosts might promote the elimination of the worms and reduce the chances of host’s re-infection. In the present study, the levels of specific IgE antibody in sera of the mice at post-infection were higher than at pre-infection. If the phenomenon is associated with high levels of IL-4 in the infected mice and if the specific IgE can help the hosts in the immunity against A. cantonensis, it is interesting.
In summary, the ratio of spleen CD4+ T cells of mice infected with A. cantonensis increases significantly, Th2 polarization appears in the immune responses, and the function of Th17 cells is inhibited. On humoral immunity, the levels of specific IgG and IgE antibodies in sera of the infected mice all increase significantly.
References
Abbas AK, Murphy KM, Sher A (1996) Functional diversity of helper T lymphocytes. Nature 383(6603):787–793
Anukumar B, Shahir P (2011) Immune regulation in Chandipura virus infection: characterization of CD4+ T regulatory cells from infected mice. Virol J 8(259):1–11
Aoki M, Sugaya H, Ishida K et al (1998) The role of CD4+ and CD8+ T-cells in host morbidity and innate resistance to Angiostrongylus cantonensis in the mouse. Parasitol Res 84(2):91–99
Chen DX, Zhang Y, Shen HX et al (2011) Epidemiological survey of Angiostrongylus cantonensis in the west-central region of Guangdong Province, China. Parasitol Res 109:305–314
Chen P, Wang WL (2009) Th1/Th2 Cytokines and worm infection. J of Pathog Biol 4(4):318–320
Chen ZL (1998) Parasite infection and cytokine. Foreign Med Sci Parasit Dis 25(5):223–227
Dorta-Contreras AJ, Noris-Garcia E, Escobar-Perez X et al (2005) IgG1, IgG2 and IgE intrathecal synthesis in Angiostrongylus cantonensis meningoencephalitis. Neurol Sci 238(1–2):65–70
Dunne DW, Butterworth AE, Fulford AJ et al (1992) Human IgE responses to Schistosoma mansoni and resistance to reinfection. Mem Inst Oswaldo Cruz 87:99–103
Gounni AS, Lamkhioued B, Ochiai K et al (1994) High-affinity IgE receptor on eosinophils is involved in defence against parasites. Nature 367(6459):183–186
Gurish MF, Bryce PJ, Tao H et al (2004) IgE enhances parasite clearance and regulates mast cell responses in mice infected with Trichinella spiralis. J Immunol 172(2):1139–1145
Harrington LE, Hatton RD, Mangan PR et al (2005) Interleukin 17-producing CD4+ effector T cells develop via a lineage distinct from the T helper type 1 and 2 lineages. Nat Immunol 6(11):1123–1132
Huang XH, Yang FZ, Zhang YZ et al (2001) Determination of antibodies in sera of rats infected with Angiostrongylus cantonensis. Strait J Prev Med 7(2):41–42
Hüttemann M, Schmahl G, Mehlhorn H (2007) Light and electron microscopic studies on two nematodes, Angiostrongylus cantonensis and Trichuris muris, differing in their mode of nutrition. Parasitol Res 101(Suppl 2):S225–S232
Jin H, He X, Liu DX et al (2006) Dynamic observation on the serum IgG level in rats infected with Angiostrongylus cantonensis. China Trop Med 6(1):21–22
Joseph M, Auriault C, Capron A et al (1983) A new function for platelets: IgE dependent killing of schistosomes. Nature 303(5920):810–812
Kolls JK, Linden A (2004) Interleukin-17 family members and inflammation. Immunity 21(4):467–476
Ko RC, Chan SW, Chan KW et al (1987) Four documented cases of eosinophilic meningoencephalitis due to Angiostrongylus cantonensis in Hong Kong. Trans R Soc Trop Med Hyg 81(5):807–810
Kushawaha PK, Gupta R, Sundar S et al (2011) Elongation factor-2, a Th1 stimulatory protein of Leishmania donovani, generates strong IFN-γ and IL-12 response in cured Leishmania-infected patients/hamsters and protects hamsters against Leishmania challenge. J Immunol 187(12):6417–6427
Langermans AM, Van der Hulst ME, Nibbering PH et al (1992) IFN-gamma-induced L-arginine-dependent toxoplasmastatic activity in murine peritoneal macrophages is mediated by endogenous tumor necrosis factor-alpha. J Immunol 148(2):568–574
Lee JD, Wang JJ, Chang JH et al (1996) Role of T cell subpopulations in mice infected with Angiostrongylus cantonensis. J Helminthol 70(3):211–214
Lindo JF, Waugh C, Hall J et al (2002) Enzootic Angiostrongylus cantonensis in rats and snails after an outbreak of human eosinophilic meningitis, Jamaica. Emerg Infect Dis 8(3):324–326
Mosmann TR, Cherwinski H, Bond MW et al (1986) Two types of murine helper T cell clone I. Definition according to profiles of lymphokine activities and secreted proteins. J Immunol 136(7):2348–2357
OuYang L, Wei J, Wu ZD et al (2012) Differences of larval development and pathological changes in permissive and nonpermissive rodent hosts for Angiostrongylus cantonensis infection. Parasitol Res 111(4):1547–1557
Padilla-Docal B, Dorta-Contreras AJ, Bu-Coifiu-Fanego R et al (2008) Intrathecal synthesis of IgE in children with eosinophilic meningoencephalitis caused by Angiostrongylus cantonensis. Cerebrospinal Fluid Res 5:18
Pan CW, Ling HB, Liang SH et al (2000) Detection of antibodies in rats infected with Angiostrongylus cantonensis by ELISA. Chin J Zoonoses 16(1):79–80
Perez O, Capron M, Lastre M et al (1989) Angiostrongylus cantonensis: role of eosinophils in the neurotoxic syndrome (Gordon-like phenomenon). Exp Parasitol 68(4):403–413
Pinot de Moira A, Fulford AJ, Kabatereine NB et al (2010) Analysis of complex patterns of human exposure and immunity to Schistosomiasis mansoni: the influence of age, sex, ethnicity and IgE. PLoS Negl Trop Dis 4(9):1–11
Qu ZY, Zhan XM, He A (2007) The analysis on epidemic situation of angiostrongyliasis in China. Int J Med Parasit Dis 34(5):279–283
Slom TJ, Cortese MM, Gerber SI et al (2002) An outbreak of eosinophilic meningitis caused by Angiostrongylus cantonensis in travelers returning from the Caribbean. N Engl J Med 346:668–675
Sugaya H, Aoki M, Abe T et al (1997) Cytokine responses in mice infected with Angiostrongylus cantonensis. Parasitol Res 83(1):10–15
Su Z, Stevenson MM (2000) Central role of endogenous gamma interferon in protective immunity against blood-stage Plasmodium chabaudi AS infection. Infect Immun 68(8):4399–4406
Wang QP, Lai DH, Zhu XQ et al (2008) Human angiostrongyliasis. Lancet Infect Dis 8(10):621–630
Watanabe N, Bruschi F, Korenaga M (2005) IgE: a question of protective immunity in Trichinella spiralis infection. Trends Parasito 21(4):175–178
Wei J, Wu F, Sun X, Zeng X, Liang JY, Zheng HQ, Yu XB, Zhang KX, Wu ZD (2012) Differences in microglia activation between rats-derived cell and mice-derived cell after stimulating by soluble antigen of IV larva from Angiostrongylus cantonensis in vitro. Parasitol Res [Epub ahead of print]
Yang BY, Shen EX, Wu CY (2006) Phenotype of mouse CD4+CD25+ regulatory T cells in mouse splenocytes. Curr Immunol 26(3):208–212
Acknowledgments
This work was supported by the grants from the National Natural Science Foundation of P.R. China (30972577) and Natural Science Foundation of Guangdong Province (s2011040003741).
Author information
Authors and Affiliations
Corresponding author
Additional information
Haijuan Liu and Xiaodong Luo contributed equally to this article.
Rights and permissions
About this article
Cite this article
Liu, H., Luo, X., Shen, E. et al. Alteration of T cell subtypes in spleen and antibodies of serum in mice infected with Angiostrongylus cantonensis . Parasitol Res 112, 1255–1260 (2013). https://doi.org/10.1007/s00436-012-3259-1
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00436-012-3259-1