
Abstract
Due to the difficulty of DNA extraction for Demodex, few studies dealt with the identification and the phyletic evolution of Demodex at molecular level. In this study, we amplified, sequenced, and analyzed a complete (Demodex folliculorum) and an almost complete (D12 missing) (Demodex brevis) ribosomal DNA (rDNA) sequence and also analyzed the primary sequences of divergent domains in small-subunit ribosomal RNA (rRNA) of 51 species and in large-subunit rRNA of 43 species from four superfamilies in Acari (Cheyletoidea, Tetranychoidea, Analgoidea, and Ixodoidea). The results revealed that 18S rDNA sequence was relatively conserved in rDNA-coding regions and was not evolving as rapidly as 28S rDNA sequence. The evolutionary rates of transcribed spacer regions were much higher than those of the coding regions. The maximum parsimony trees of 18S and 28S rDNA appeared to be almost identical, consistent with their morphological classification. Based on the fact that the resolution capability of sequence length and the divergence of the 13 segments (D1–D6, D7a, D7b, and D8–D12) of 28S rDNA were stronger than that of the nine variable regions (V1–V9) of 18S rDNA, we were able to identify Demodex (Cheyletoidea) by the indels occurring in D2, D6, and D8.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Eukaryotic ribosomal DNA (rDNA), the gene encoding the ribosomal RNA (rRNA) in cell nucleus, comprises a big multi-gene family in the form of tandem multi-copy. Each copy is composed of structural genes including large-subunit (LSU) 28S rDNA, 5.8S rDNA, and small-subunit (SSU) 18S rDNA and nonstructural genes including non-transcribed and transcribed spacer regions. With variable evolution rates, different rDNA portions can be used for phylogenetic study at different taxonomic category levels. Compared with the internal transcribed spacers 1 and 2 (ITS-1 and ITS-2), 18S and 28S rDNA share lower evolution speeds and are made up of conserved regions and divergent domains (variable regions in 18S rDNA and expansion segments in 28S rDNA). Both 18S and 28S rDNA have been the extraordinarily helpful molecular markers for phylogenetic analysis until now (Baverstock and Johnson 1990).
Demodex mites belong to the class Arachnida, Acariformes, Cheyletoidea, and Demodicidae. Since Simon's report on Demodex in 1842, there have been 140 Demodex species or subspecies identified from 11 orders of mammals (dog, sheep, cat, pig, mouse, etc.), residing in hair follicles, sebaceous glands, meibomian glands, and internal organs. Demodex folliculorum and Demodex brevis are two types of human Demodex (Desch and Nutting 1972). For a long time, the classification and phylogenetic study of Demodex have been based on the hosts and phenotype features of the mites, which may lead to phylogenetic indeterminacy and difficulties. Therefore, it is necessary to study Demodex at the gene level.
However, there are only a few studies on Demodex at a molecular level (Toops et al. 2010; Zhao and Wu 2011, 2012; De Rojas et al. 2012), and there is no report on the molecular data of its rDNA. The reasons are threefold. First, it is hard to obtain standardized mites for experiments. Demodex mites are species-specific to their hosts. Other than a few reports about cross-infection between human and animals (Morsy et al. 1995; Wang et al. 1998), the mites have not been maintained in vitro successfully (Zhao et al. 2009a, 2010). Second, Demodex DNA extraction is difficult due to the tiny body and thick chitin of the mites (Zhao et al. 2009b). Third, it is hard to get sufficient number of Demodex mites from the same host for genomic DNA extraction. A large number of mites would be needed for the gDNA extraction at a time, i.e., 1,500 mites/host for D. folliculorum and 2,000 mites/host for D. brevis (Zhao and Cheng 2009). Despite these difficulties, we successfully amplified and sequenced the complete rDNA sequences (18S, ITS-1, 5.8S, ITS-2, and 28S) of the two human Demodex mites, which were further analyzed, for the first time in this study. And the primary sequence analysis of nine variable regions in SSU rRNA and the 13 expansion segments in LSU rRNA for four superfamilies (Cheyletoidea, Tetranychoidea and Analgoidea in Acariformes, and Ixodoidea in Parasitiformes) in Acari were conducted, in terms of sequence lengths, evolution rates, and divergences. The findings in the present study have provided molecular data for Demodex study at the gene level.
Materials and methods
Demodex collection and DNA extraction
In this study, ethical permission was not required because the mite collection method was a noninvasive sampling technique routinely used in pathogen. All the sufferers were from Xi'an, China. Cellophane tape method was applied to sample the isolates of D. folliculorum and D. brevis from the sufferers' faces. Mite samples were collected using a self-made needle under a 10 × 4 microscope and preserved in Eppendorf tubes at −20 °C. The mites were then put into liquid nitrogen for 1 min of freeze thawing, followed by a 10-min milling, and this procedure was repeated four times. After that, DNA extraction was conducted using a DNA extraction kit (Omega) obeying the manufacturer's protocol (Zhao et al. 2009b).
Primer design
Specific primers were designed respectively for the two segments of 18S rDNA, ITS region, and the eight segments of 28S rDNA according to the corresponding conserved regions identified in the alignments of referred sequences from GenBank (Table 1). The primers were synthesized in Beijing AuGCT Biotechnology Co., Ltd. The model species and the designed primers are listed in Table 1.
PCR amplification, cloning, and sequencing
All PCR reactions were conducted on AB Applied Biosystems 2720 Thermal Cycler. Each 12.5-μL reaction volume contained 1 μL of DNA template, 1 μL of each primer (1 μmol/L), 6 μL of 2× Premix Taq buffer (pH 8.3, 0.05 U/μL Taq DNA polymerase, 3 mmol/L MgCl2, and 0.4 mmol/L each dNTP), and 3.5 μL of double-distilled water. The PCR conditions were as follows: an initial degeneration of DNA for 5 min at 94 °C, followed by 40 cycles of 1-min degeneration at 94 °C, annealing for 1 min at proper temperature (Table 1) and extension for 2 min at 72 °C, and finally extension for 10 min at 72 °C. The PCR products were separated by electrophoresis in 2 % agarose gels and visualized under UV light using 100 bp DNA ladder (TaKaRa) as the marker.
The PCR products were purified with Omega Gel Extraction Kit, recombined with pMD18-T vector (TaKaRa), and then cloned into Escherichia coli (DH5α), which was followed by ampicillin selection. After the plasmids were extracted, PCR amplified, and identified, the bacterial liquid with positive plasmids was chosen for sequencing in Beijing AuGCT Biotechnology Co., Ltd.
Sequence alignments and analysis
After being aligned in DNAssist version 1.0, the amplified sequence segments (Table 1) were combined to form complete rDNA sequences for D. folliculorum and D. brevis separately. Apis mellifica and Drosophila melanogaster (Insecta: Endopterygota) were taken as outgroups. Species from four Acari superfamilies (Cheyletoidea, Tetranychoidea, Analgoidea, and Ixodoidea), whose 18S rDNA and 28S rDNA were available in GenBank, were chosen to reconstruct 18S and 28S rDNA maximum parsimony (MP) phylogenetic trees under Maximum Composite Likelihood model in MEGA 4.0 (Tamura et al. 2007). Next, based on the alignment results obtained from ClustalX (version 1.8; Thompson et al. 1997), the five regions of rDNA sequences (Table 2) as well as the divergent domains of SSU rDNA and LSU rDNA were identified in line with that of A. mellifica (Insecta: Hymenoptera) (Gillespie et al. 2006), with small adjustments according to the predicted secondary structures (see our follow-up research paper). Finally, the lengths of the divergent domains and their divergences compared to D. folliculorum were calculated.
Results and discussion
Complete rDNA sequence
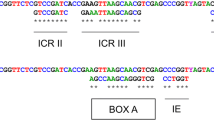
In this study, 11 gDNA samples were extracted successfully from 11 isolates of three Demodex species (five D. folliculorum, three D. brevis, and three Demodex canis). However, only one complete D. folliculorum rDNA sequence (6,397 bp) and one almost complete D. brevis sequence (5,837 bp, lacking 28S terminal with D12) were obtained. For D. canis, only the complete 18S rDNA sequence and a large part of 28S rDNA sequence were obtained, but no sequence of the ITS regions was obtained. Therefore, only one D. folliculorum isolate and one D. brevis isolate were included in Demodex sequence alignments. The alignment results and the division of divergent domains can be found in Supplementary Material 1.
We also showed that, for D. folliculorum, V1–V9 covered 38.5 % (697/1809) of its 18S rDNA, and D1–D12 covered 43.0 % (1,743/4,050) of its 28S rDNA. For D. brevis, V1–V9 accounted for 38.9 (710/1824) of the 18S rDNA, and D1–D11 took 34.0 % (1,135/3,340) of the 28S rDNA (D12 lacked; Table 2).
Table 2 shows that the complete 18S rDNA sequences of D. folliculorum and D. brevis were the most conserved, with the lowest divergence and most approximate GC contents, and they were followed by less conserved 5.8S rDNA and then 28S rDNA. ITS-1 and ITS-2 shared the fastest evolution rates. It was indicated that different portions of rDNA might be used in phylogeny analysis at different taxonomic category levels due to discrepant sequence variances.
Since the complete 28S rDNA sequences were limited to just four superfamilies in Acari, only the 18S rDNA of 51 species from the same taxa was chosen to contrast the resolution capacity of 28S sequenced with that of the 18S rDNA sequences. It was found that the MP trees derived from the 18S rDNA and the 28S rDNA (Figs. 1 and 2) were identical at superfamily level and above. The species of each superfamily would form a cluster first, then the taxa of Acariformes and Parasitiformes would form two distinct branches and gathered later, forming the Acari branch that would eventually gathered with A. mellifica and D. melanogaster, the outgroup species. These results were in line with the traditional classification based on phenotype characteristics, demonstrating an important role of both 18S rDNA and 28S rDNA primary sequences in phylogeny across Acari.

MP tree based on 18S rDNA complete or almost complete sequences from 51 Acari species and two outgroups and lengths and divergences between D. folliculorum and any other species for each variable region (V1–V9)

MP tree based on 28S rDNA complete or almost complete sequences from 43 Acari species and two outgroups and lengths and divergences between D. folliculorum and any other species for each expansion segment (D1–D12)
V1–V9 and D1–D12 regions
Compared to complete rDNA sequences, the divergent domains shared higher evolution rates, suggesting a greater significance of these domains in phylogenetic study. In this study, in 18S rDNA, in terms of sequence length, in the inter- and intra-superfamilies in Acari, except the obvious insertions in V2, V4, and V7 in Tetranychoidea, slight indels (usually less than 10 bp) were found in V1, V3, V5, V8, and V9 and none in V6 in all the studied species including the outgroups A. mellifica and D. melanogaster. In terms of divergences, the pairwise sequence divergences between D. folliculorum and any other species from the four Acari superfamilies were in accordance with the phylogenetic relationship shown in the MP trees. However, the V3 divergences between Cheyletoidea and Ixodoidea (Parasitiformes) were smaller than those between Cheyletoidea and Analgoidea (Acariformes). For the inter-superfamilies, the divergence between Demodicidae (Cheyletoidea) and others reached the highest in V9 (35.2–74.1 %) and V8 (32.3–44.6 %) and was lowest in V6 (3.9–11.8 %) and V3 (8.1–24.3 %). Further investigation showed that V2, V4, and V7 were the most valuable regions for discrimination of 18 rDNA. The divergences of D. folliculorum (Cheyletoidea) compared with the species in the other three superfamilies were all within the range of 11.0 to 34.7 % (Fig. 1), except for the larger divergences compared with Tetranychoidea (48.0–82.0 %) due to insertions.
Much more differences in sequence length and divergence were identified in 28S rDNA compared with 18S rDNA. A deletion of about 200 bp occurred in Demodex in D2, and an insertion of about 20–30 bp occurred in D6 in Demodex as well as in A. mellifica. In D8, D. folliculorum and D. canis had an insertion of about 200 bp, and the divergences between D. folliculorum or D. canis and other superfamilies were high. There was no insertion in D. brevis. The D12, which was absent in some species, was much longer in D. folliculorum. It was worth noticing that D3, D5, D9, and D10 were expanded to various extents in Tetranychoidea, and D3 and D7a of Tetranychus pacificus, Raoiella indica, and Petrobia harti were more significantly expanded. Between Demodicidae and any other taxon, the most different segments were D9 (57.6–89.8 %) and D10 (63.4–81.0 %), followed by D6, D5, D2, D8, D12, D1, D4, D11, D3, and D7b in descending order. D7a was the segment with the smallest divergences (17.7–30.6 %), except in T. pacificus, R. indica, and P. harti of Tetranychoidea due to insertions.
Conclusions
In this study, we have amplified, cloned, sequenced, and analyzed a complete and an almost complete rDNA sequence of 18S, ITS-1, 5.8S, ITS-2, and 28S of the two human Demodex, D. folliculorum and D. brevis. Then, we analyzed the sequences of all the divergent regions in the four Acari superfamilies whose 18S rDNA and 28S rDNA were both available in GenBank. It can be inferred that 28S rDNA is more valuable than 18S rDNA in molecular discrimination at the gene level.
References
Baverstock PR, Johnson AM (1990) Ribosomal RNA nucleotide sequence: a comparison of newer methods used for its determination, and its use in phylogenetic analysis. Aust Syst Bot 3:101–110
De Rojas M, Riazzo C, Callejón R, Guevara D, Cutillas C (2012) Morphobiometrical and molecular study of two populations of Demodex folliculorum from humans. Parasitol Res 110:227–233
Desch CE, Nutting WB (1972) Demodex folliculorum (Simon) and D. brevis (Akbulatova) of man: redescription and revaluation. J Parasitol 58:167–177
Gillespie JJ, Johnston JS, Cannone JJ, Gutell RR (2006) Characteristics of the nuclear (18S, 58S, 28S and 5S) and mitochondrial (12S and 16S) rRNA genes of Apis mellifera (Insecta: Hymenoptera): structure, organization, and retrotransposable elements. Insect Mol Biol 5:657–686
Morsy TA, el Okbi MM, el-Said AM, Arafa MA, Sabry AH (1995) Demodex (follicular mite) infesting a boy and his pet dog. J Egypt Soc Parasitol 25:509–512
Tamura K, Dudley J, Nei M, Kumar S (2007) MEGA4: molecular evolutionary genetics analysis (MEGA) software version 4.0. Mol Biol Evol 24:1596–1599
Thompson JD, Gibson TJ, Plewniak F, Jeanmougin F, Higgins DG (1997) The CLUSTAL_X windows interface: flexible strategies for multiple sequence alignment aided by quality analysis tools. Nucleic Acids Res 25:4876–4882
Toops E, Blagburn B, Lenaghan S, Kennis R, MacDonald J, Christine D (2010) Extraction and characterization of DNA from Demodex canis. Intern J Appl Res Vet Med 8:31–43
Wang YP, Li P, Bing GQ (1998) A case report of human dermatitis caused by canine Demodex. J Norman Bethune Univ Med Sci 24:265 (In Chinese)
Zhao YE, Cheng H (2009) RAPD analysis and sequence alignment of genomic DNA of hair follicle mites Demodex folliculorum and D. brevis (Acari: Demodicidae). Acta Entomol Sin 52:1273–1279 (In Chinese)
Zhao YE, Wu LP (2011) RAPD-SCAR marker and genetic relationship analysis of three Demodex species (Acari: Demodicidae). Parasitol Res. doi:101007/s00436-011-2778-5
Zhao YE, Wu LP (2012) Phylogenetic relationships in Demodex mites (Acari: Demodicidae) based on mitochondrial 16S rDNA partial sequences. Parasitol Res. doi:10.1007/s00436-012-2941-7
Zhao YE, Guo N, Wu LP (2009a) The effect of temperature on the viability of Demodex folliculorum and Demodex brevis. Parasitol Res 105:1623–1628
Zhao YE, Cheng H, Xun M, Wu LP (2009b) Extraction and random primer PCR detection of genomic DNA of parasitic mites Demodex folliculorum and Demodex brevis (Acari: Demodicidae). Acta Entomol Sin 52:929–933 (In Chinese)
Zhao YE, Guo N, Wu LP (2010) The influence of temperature and medium on viability of Demodex folliculorum and Demodex brevis (Acari: Demodicidae). Exp Appl Acarol 54:421–425
Author information
Authors and Affiliations
Corresponding author
Electronic supplementary material
Below is the link to the electronic supplementary material.
Supplementary Material 1
The alignment of a complete nuclear rDNA sequence of Demodex folliculorum (18S, ITS-1, 5.8S, ITS-2 and 28S) and an almost complete sequence of Demodex brevis. ITS-1 and ITS-2 are noted by dark shades, and the 9 variable regions of SSU rDNA and 13 expansion segments of LSU rDNA are noted by light shades. (JPEG 2.30 mb)
Rights and permissions
About this article
Cite this article
Zhao, YE., Wu, LP., Hu, L. et al. Sequencing for complete rDNA sequences (18S, ITS1, 5.8S, ITS2, and 28S rDNA) of Demodex and phylogenetic analysis of Acari based on 18S and 28S rDNA. Parasitol Res 111, 2109–2114 (2012). https://doi.org/10.1007/s00436-012-3058-8
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00436-012-3058-8