
Abstract
In this study, 51 piglets originating from five different sows were included in the investigations. The animal source of all sows had a history of Clostridium perfringens type A (β2) infection. The piglets of three sows (n = 31) were experimentally infected with Isospora suis within the first 4 h after birth and were randomly assigned to the treatment group or the sham-dosing group. The piglets of the two remaining sows (n = 20) served as I. suis-uninfected controls. Twelve hours post-infection, the animals in the treatment group (n = 15) were treated with toltrazuril (20 mg/kg BW, Baycox® 5% suspension). During an observation period of 14 days faecal consistency, faecal oocyst counts, faecal germ counts, mortality, body weight development and clinical status were recorded. One piglet per study group and litter was necropsied, and intestinal tissue samples were taken for histopathological investigations and in situ hybridisation on study days (SDs) 3 and 14. I. suis-infected but untreated piglets showed clinical disease resulting in liquefaction of faeces and decreased body weight development. In 59.2% of the observations, I. suis-infected but untreated piglets showed abnormal faecal consistencies whereas only 12.0% or respectively 4.4% of the faecal samples had a pasty consistency in the I. suis-infected–treated or in the control animals. The mean body weight at the end of the study was 3.37 kg in the I. suis-infected but untreated piglets while the average body weight in the I. suis-infected–treated animals was calculated as 4.42 kg and the control animal’s mean body weight was 4.45 kg. Moreover, mortality, occurring between SDs 8 and 14, in this study group was 38.5% (n = 5), with 30.8% (n = 4) died from necrotic enteritis. In contrast, no piglets died in the I. suis-uninfected control group or in the toltrazuril-treated study group. The results of this study corroborate the hypothesis that simultaneous infection with I. suis and C. perfringens type A soon after birth leads to distinct interactions between the two pathogens and result in an increase in clinical disease, mortality and metabolically active C. perfringens type A.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Toltrazuril has been shown to be an effective anticoccidial compound when used in poultry, pigs, calves, lambs and other species exposed to experimental or natural infections with coccidia. In piglets, Isospora suis is generally regarded as a primary causative agent of enteritis in young piglets, and metaphylactic treatment with toltrazuril 3 to 5 days after birth is a common control measure practised in many countries. On the other hand, other infectious organisms may occur simultaneously and contribute to the clinical course of infection, or even trigger the disease in the presence of per se subclinical exposure to I. suis (Westphal et al. 2007). Clostridium perfringens is a ubiquitous agent and part of the physiological gut flora of neonatal piglets. Piglets are often typically infected with I. suis and with clostridia, e.g. C. perfringens, and show clinical disease associated with the infections (McDonel 1986; Songer 1996; Songer and Anderson 2006; Songer and Uzal 2005). A correlation between necrotic enteritis due to clostridial infection and coccidiosis has been clearly demonstrated in chickens (Baba et al. 1997; Collier et al. 2008; Williams et al. 2003). However, such data are virtually lacking for mammalian coccidiosis although natural disease related to co-infection appears probable according to observations in the field (Westphal et al. 2007). Histopathological studies have shown that the intestinal mucosa is severely altered where I. suis reproduces, opening the mucosal barrier for other potentially hazardous pathogens (Bach et al. 2003). The present study explores the clinical response of newborn piglets exposed to natural C. perfringens type A infection to low-dose experimental infection with I. suis.
Materials and methods
The study was designed as a randomised, single-blinded, double-negative controlled clinical study. The sows (commercial breed) originated from a farm with known history health problems due to clostridial infection and were thus expected to excrete clostridia with their faeces. The current bacteriological status of the sows (excretion of clostridia) was estimated by examination of three individual faecal samples during the periparturient period, i.e. one sample antepartum, one on the day of littering and one on study day (SD) 3. The calculated day of farrowing was within 2 weeks after housing in the experimental farrowing unit. Because piglets meant to serve as uninfected controls are at high risk of attracting accidental infection when kept in the same unit as piglets experimentally infected with I. suis, two separate but equally equipped stables were used (infection stable, control stable). Two sows (nos. 3 and 5) were randomly assigned to the control stable and three (nos. 1, 2, 4) were kept in the infection stable. The litters of sows nos. 3 and 5 comprised 9 and 11 piglets; the litters of sows nos. 1, 2 and 4 contained 8, 10 and 13 piglets, respectively. Healthy piglets weighing at least 0.8 kg at birth were included in the study. They were individually marked with ear tattoos within 4 h after the completion of farrowing including at least a first intake of colostrum. In total, 51 piglets participated in the study.
The groups were denominated on the basis of experimental Isospora infection (I+, I−) and toltrazuril treatment (T+, T−). Piglets destined to be experimentally infected with I. suis oocysts (strain LE-11-I. suis-98/X+8) (I+) were randomly allocated immediately after birth across litters nos. 1, 2 and 4 to either the treatment group (I+T+, n = 15) or to the sham-treated group (I+T−, n = 16). All the piglets in litter no. 3, i.e. nine animals, and five piglets from litter no. 5 formed group I−T− (uninfected, sham-treated, n = 14) and the remaining six piglets from litter no. 5 represented group I−T+ (uninfected, toltrazuril treatment).
The sows were fed with commercial feed for farrowing sows, and water was freely available. Piglets received milk from the sow and, from the age of 10 days, were additionally fed with probiotic-free piglet starter feed. All piglets received iron supplementation within the 3 days post natum to prevent iron deficiency.
Day 0 of the study was defined as the day of birth, i.e. the time point of the experimental infection (SD 0).
The experimental infection took place within 4 h after conclusion of the individual sow’s littering. All piglets in groups I+T+ and I+T− were individually inoculated with 1,000 sporulated oocysts of I. suis suspended in 2 ml of tap water by a single-use flexible plastic pipette deep into the throat. The piglets in groups I+T+ and I−T+ received a single oral drench of Baycox® 5% suspension (20 mg toltrazuril/kg BW) 12 h after artificial infection with I. suis oocysts, while 0.5 ml tap water was similarly administered to those in groups I+T− and I−T−.
The following parameters were recorded: semiquantitative oocyst counts, germ counts (e.g. C. perfringens, Clostridium difficile, total germ count), mortality, weight gain, pathological and clinical observations (faecal consistency, behaviour, skin turgidity, conjunctiva, body temperature, coughing and other noticeable observations). The health status of the individual piglets was recorded immediately after birth and daily until the termination of the experiment on day 14 (SD 14) of the study.
During the study period, 18 piglets (one piglet per litter and study group each on SDs 3 and 14) were euthanised for necropsy.
Oocyst excretion
For oocyst quantification and evaluation of faecal consistency, samples were collected individually on SDs 3, 7, 10 and 14 from the rectum by stimulation of the anus with a lightly greased cotton swab until at least 1 g of faeces was released spontaneously into an appropriately labelled tube.
For estimation of oocyst excretion, drops of pasty to liquid faeces were smeared on microscopic glass slides (two slides per sample) with a single-use pipette. Dry, formed faeces were homogenised in a few drops of distilled water with a wooden spatula, and the resulting suspensions were smeared. Oocysts were identified by epifluorescence microscopy (Daugschies et al. 2001; Kuhnert et al. 2006) and counted semiquantitatively at ×100 magnification in five separate randomly chosen views per slide. Each view was scored individually, and the average score was calculated for the sample.
The scoring system is described in Table 1.
Faecal consistency
Faecal consistency was judged immediately after faecal sampling and recorded using a scoring system representing four different qualities (see Table 2).
Determination of faecal Clostridia counts
Germ counts for C. perfringens and C. difficile were determined after cultivation in specific culture media in faecal samples collected on SD 1, 2, 3 and 14 (Table 3). In addition, one faecal sample per sow collected before littering, on the day of littering and 3 days postpartum was examined bacteriologically.
Examination of intestinal tissue samples
On study days 3 and 14, one piglet per study group and per litter underwent histopathological and micromorphological examinations. These study animals were sacrificed and intestinal tissue samples were taken at necropsy from the duodenum, jejunum, ileum, caecum and colon. Three samples at a time were taken from each intestinal site and stored in Carnoy medium at a temperature of +4°C to +8°C until examination.
Fluorescence in situ hybridisation was performed according to the method as established and described by Swidsinski et al. (2005).
Six different probes, including Non-338 nonsense probe for specific binding verification, were used for in situ hybridisation. Results shown and discussed in this paper belong to the investigations with Eub338, Chis150 and Clis135 probes. The Eub338 probe (Amann et al. 1990) was used to detect eubacteria. For differentiation of Clostridium spp., the Chis150 probe was used for the Clostridium histolyticum group (the organism to be expected in the samples collected in this trial was C. perfringens), and the Clis135 probe was used for the Clostridium lituseburense group (which includes C. difficile) (Franks et al. 1998). The Chis150 probe is specific for a number of species within Clostridium clusters I and II, i.e. C. histolyticum, Clostridium beijerinckii, Clostridium butyricum, C. perfringens, Clostridium putrificum and Clostridium cadaveris. Clis135 probe is specific for some species of the C. lituseburense group which represents a part of Clostridium cluster XI. Target species of Clis135 are Eubacterium tenue, Clostridium ghoni, Clostridium sordellii, Clostridium bifermentans, C. lituseburense, C. difficile and Clostridium mangenotii.
Body weight development
Individual body weights of the piglets were investigated after conclusion of the according sow’s littering and on the last day of the study, i.e. on study day 14.
Statistics
All the raw data from the study were transferred to spreadsheets. The statistical programme Testimate Vers. 6.4 from IDV Gauting was used for analysis, and the recorded parameters were tested for normal distribution and analysed using the two-sided Wilcoxon–Mann–Whitney U test, Fisher’s exact test and descriptive statistics.
Results
Clinical observations (excluding faecal score)
No adverse effects on piglet health of administering toltrazuril were observed; neither was a positive effect observed in uninfected piglets. Also, the bacteriological examinations as total aerobe germ counts, total anaerobe germ counts, enterobacteria or lactobacillus proved no statistical significant differences between the I. suis-uninfected toltrazuril-treated or I. suis-uninfected toltrazuril-untreated animals with exception of the total aerobe germ counts on SD 14 and the enterobacteria counts on SD 3. The piglets in groups I−T+ and I−T− were therefore considered to be identical in status and were combined to form control group I− for data analysis.
Oocyst excretion
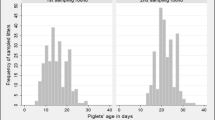
Oocysts were first detected on SD 7 in the faeces of nine out of 13 piglets in group I+T− (Fig. 1). Two of the four piglets that were negative on this occasion became positive on SDs 10 and 14, respectively, while the remaining two piglets showed no oocyst excretion at any of the faecal oocyst examinations during the study. Most piglets continued to excrete oocysts on the following days, and six out of ten piglets were still positive on SD 14.

Oocyst excretion
Of the 12 piglets infected with I. suis and additionally treated with toltrazuril (I+T+), only one excreted very low numbers of oocysts on one occasion (SD 10, oocyst score 1).
No oocysts were detected in the faeces of the control animals (I−T−, I−T+). Statistically, oocyst excretion in group I+T+ differed significantly (P = 0.0056 to P = 0.0005) from the values in group I+T− but not from those of the uninfected controls (I−).
Clostridia excretion and in situ hybridisation
Clostridia excretion—sows
C. perfringens type A was detected in faecal samples from all sows, and the highest numbers of excreted germs were detected in faecal samples antepartum. No C. difficile was isolated from sows.
The number of shed clostridia decreased distinctly in all sows after littering and started to increase again at the last sampling (SD 3).
The highest numbers of C. perfringens were excreted by the sow of litter 4 with 7.18 × log10 cfU/g at the antepartum sampling, followed by the sow of litter 5 (6.92 × log 10 cfU/g) whose piglets belonged to groups I−T+ and I−T−. The third and fourth highest numbers of excreted C. perfringens type A were found in the samples from the sows of litter 2 (6.65 × log10 cfU/g; I+T− and I+T+) and litter 3 (6.62 × log10 cfU/g; I−T−). The lowest numbers of C. perfringens type A were found in the faeces of the sow of litter 1, which shed 6.2 × log10 cfU/g antepartum.
Clostridia excretion—piglets
C. perfringens type A (β2) was the most prominent clostridia to be found in the faecal samples within the study period. C. perfringens type C was detected only in some piglets of the I. suis-infected litters. C. difficile was not detected in any faecal sample of the piglets. C. perfringens type A was found in faecal samples from all study groups. The mean faecal colony-forming units per sample were higher for animals in study group I+T− (4.94 × log10 to 6.68 × log10 cfu/g) than in study group I+T+ (2.65 × log10 to 4.70 × log10 cfu/g) until the last faecal sampling on SD 14. This difference was statistically significant for faecal examinations on SDs 3 and 14 (P = 0.049 and P = 0.029). The differences in faecal Clostridia excretion on SDs 1 and 2 could not be proofed statistically (P = 0.052 and P = 0.298). On SD 14, significantly higher numbers were counted for the I. suis-infected, toltrazuril-treated study group. The groups not infected with I. suis (I−) showed an independent dynamic of microflora composition that differed clearly from both artificially infected study groups (I+T− and I+T+; Table 4).
In situ hybridisation
The results of in situ hybridisation examinations of the jejunum using Eub338, Chis150 and Clit135 probes (Amann et al. 1990; Franks et al. 1998) show considerable differences between study animals belonging to study groups I+T− and I+T+ (see Fig. 2a, b; Table 5). Animals in study group I+T− showed distinctly higher numbers of Chis150-detectable clostridia in the small intestine than piglets belonging to the infected study group treated with toltrazuril (I+T+). This difference was investigated in samples taken on SD 3, while on SD 14 this distinct difference was no longer detectable. The average number of Chis150-detectable clostridia in the jejunum of animals in study group I+T− was 3.0 ± 2.7 × 109/ml on SD 3, while for study group I+T+, a mean of 0.007 ± 0.01 × 109/ml was determined and in study group I− no Chis150-detectable clostridia could be found.
Further data collected from microbiological investigations, including PCR typing of C. perfringens toxin genes, and serological examinations are discussed by Krüger et al. (2010).

a Colonisation with clostridia—jejunum (piglet no. 2/4, group I+T+, SD 3), Chis150 Cy5-Probe, 1,000-fold. b Colonisation with clostridia—jejunum (piglet no. 4/4, group I+T−, SD 3), Chis150 Cy5-Probe, 1,000-fold (Swidsinski 2007)
Faecal consistency
Faecal scores—SD 1
On SD 1, increased faecal score values were observed in all study groups (I+T−, I+T+, I−). This liquefaction of the faecal consistency was found in six animals out of a total of 14 in study group I+T− (two of faecal score of 1, four of faecal score of 2), seven out of 15 piglets in study group I+T+ (four of faecal score of 1, three of faecal score of 2) and in the control group (I−) 14 animals out of 20 were noted as having a faecal score >0 (three of faecal score of 1, 11 of faecal score of 2; Fig. 3). As this first faecal sampling was done between approximately 9 and 20 h after treatment (21 to 32 h after the artificial infection) and taking the mode of action of toltrazuril (e.g. sensitive parasite stages) into consideration, the observed alterations of faecal consistency cannot be explained by infection or treatment but were obviously due to stress particularly in newborn piglets.
Therefore, SD 1 was not considered in the further analysis and the following explanations refer to the period SDs 3 to 14.

Distribution of faecal consistencies
Faecal scores—SDs 3 to 14
Study group I+T− showed distinctly higher average faecal scores than the other groups from SD 3 onwards (Fig. 4). Faecal scores >0 were documented for 29 samples in piglets in group I+T− (59.2%) during the experimental study period. Of these, almost 30% (eight samples) were liquid to watery (score 2) while the others were pasty to semiliquid (score 1). A faecal score of 2 was observed exclusively in animals in group I+T−.

Median faecal scores per study group per study day
In contrast, most of the piglets in group I+T+ showed no diarrhoea. Pasty to semiliquid faeces (score 1) were occasionally recorded (six out of 50 samples, i.e. 12.0%) for piglets between SDs 3 and 14.
In the uninfected control group (I−), only three out of 20 animals (three out of 68 samples, i.e. 4.4%) had the maximum faecal score of 1 on SD 3 or later, with the highest average value of 0.10 being calculated for SD 3.
Statistically, group I+T+ piglets showed clearly less severe clinical disease compared to I+T− piglets over the whole study period (P = 0.000), although average scores were actually slightly higher in group I+T+ than in the two other groups on SD 3. No statistical difference in faecal scores was evident between group I+T+ and I−.
Body weight
The average body weight in the study groups was 1.28 kg (I+T−) or 1.29 kg (I+T+, I−) on SD 0. At birth, the lightest piglet weighed 0.86 kg (I+T−, I+T+) or 0.90 kg (I−) and the heaviest 1.70 kg (I+T−), 1.68 kg (I+T+) and 1.63 (I−). Therefore, the groups are considered to be very homogeneous in terms of weight at birth.
When the experiment was concluded on SD 14, the piglets in group I+T− had a mean body weight of 3.37 kg with a range of 1.66 to 3.55 kg. The average weight in group I+T+ was considerably higher at 4.42 kg (range, 2.54 to 5.54 kg), and these values were almost identical to those in group I− (average, 4.45 kg; range, 2.60 to 5.29 kg; see Fig. 5). When the weight gains of piglets in groups I+T+ and I− were compared with those in group I+T−, the difference was statistically different (P = 0.0002), whereas no such difference was present between groups I+T+ and the controls in group I− (P > 0.05).

Body weight development
Mortality
Unforeseen deaths occurred in five of the study animals during the study period. Natural mortality occurred between SDs 8 and 14. The numbers of piglets per study group within this study period, i.e. after the first experimental necropsies, were 13 piglets in study group I+T−, 12 piglets in study group I+T+ and 17 piglets served as uninfected controls (I−). The piglets showing natural mortality belonged to litter no. 2 (one piglet dead on SD 8) and litter no. 4 (one piglet on SD 8, one piglet on SD 11, two piglets on SD 14), and all of them were infected with I. suis but not treated (group I+T−).
One piglet in litter no. 4 was diagnosed as probably having died as a result of meningitis and massive ulceration of the whole non-glandular part of its stomach, and death was thus obviously not related to the experimental conditions.
According to the necropsy reports, four piglets died from enteritis, i.e. one animal with catarrhal enteritis, and all other piglets were found to suffer from necrotic enteritis (see Fig. 6). In addition to Isospora stages, moderate to massive intestinal colonisation with microscopically ‘bacillary bacteria’/‘clostridia-like bacteria’ was recorded in all cases. Thus, almost 38.5% of the piglets in group I+T− died, 30.8% obviously from necrotic enteritis. No such observations were made in any of the other groups.

Necropsy results—piglet group I+T−, SD 3: necrotic enteritis
Discussion
It is generally acknowledged that intestinal coccidiosis caused by I. suis is one of the major drawbacks in piglet production (Maes et al. 2007; Mavromatis et al. 2004). Toltrazuril is widely administered as a metaphylactic drug to avoid diarrhoea due to coccidiosis in suckling piglets and to prevent the related economic losses. Although the prevalence of I. suis is very high, particularly during the first weeks of life, this is not always associated with clinical disease (Niestrath et al. 2002). Experimental single infections with I. suis induce typical diarrhoea and have been shown to result in an average of 5 to 7 days of oocyst shedding, mild loss of body weight (gains) and histopathological villous necrosis or atrophy, mainly in the anterior part of the jejunum. This starts within the first 5 days p.i. and decreases in intensity until 14 days p.i. (Mundt et al. 2003, 2006). The presence of a second pathogen is not necessary to induce clinical disease (Harleman and Meyer 1984, 1985; Vítovec and Koudela 1990).
In addition to its role as a primary pathogen, I. suis has been assumed to also participate in multifactorial diarrhoea and may in particular be associated with necrotic enteritis due to clostridial infection. This may occur soon after birth, whereas typical isosporosis is generally seen later (second and third weeks of life).
C. perfringens type A agents are parts of the physiological flora of the large bowel in swine herds, but under certain circumstances, they can cause severe diseases. We hypothesised that I. suis infection of newborn piglets in concert with exposure to C. perfringens type A might lead to enteritis at an early age that depends on the presence of both pathogens (Krüger et al. 2010). This hypothesis has been corroborated by the results of this trial.
The study was performed simultaneously for all 51 study animals, although the study schedule was adapted individually for each litter on the basis of farrowing dates. To model neonatal necrotic enteritis due to co-infection, newborn piglets naturally exposed to C. perfringens type A were additionally artificially infected with 1,000 oocysts of I. suis, a dose that is below the standard experimental dose for I. suis (Mundt et al. 2006, 2007). Even lower oocyst numbers may induce clinical disease when inoculated into young piglets (Mundt et al. 2003); however, the bacteriological status of these animals was not recorded.
Although the piglets in the I. suis-infected litters were randomised per litter, the piglets in study group I+T− excreted significantly higher numbers of C. perfringens in comparison with both the animals in group I+T+ and the controls that were not infected with I. suis (I−) until SD 14. However, faecal microbiological investigations of clostridia do not show the effective amount of microflora, i.e. the number of intestinally active germs, and moreover depend on the faecal consistency as the results are calculated on basis of moist mass. The intestines of slaughtered animals therefore underwent examination by in situ hybridisation to investigate the metabolic activity of the clostridia in the intestinal tissue samples. Fluorescence in situ hybridization (FISH) response to the used 16S rRNA probes depends on the amount of ribosome per cell which corresponds to growth rates of the agents (Amann et al. 1995; Poulsen et al. 1993). This influence is most prominent in samples of study group I− which had the highest numbers of colonies per gram of faeces on SD 3 but showed no detectable response at the FISH investigation of the same study day’s tissue samples. A lack of metabolic activity of the C. perfringens in these samples may be concluded. Therefore, the distinctly higher response in samples of study group I+T− in comparison to the infected and toltrazuril-treated group (I+T+) shows also an increased metabolic activity of the intestinal clostridia in the animals of this group on SD 3. On SD 14, there were no longer differences in results of the in situ hybridisation detectable. The fact that differences in the number of intestinally metabolic active clostridia were only detectable in the jejunum, the section of the intestines mainly affected by I. suis infections (Niestrath et al. 2002; Robinson et al. 1983; Stuart et al. 1980) supports the assumption of interaction between the two pathogens. Consequently, mortality was observed in 38.5% of piglets exposed to isosporosis and clostridiosis (I+T−), whereas no such observations were recorded in the absence of Isospora infection or when infected piglets were appropriately treated with Baycox® 5% suspension (toltrazuril).
Our results show that piglets with high faecal C. perfringens type A counts but not infected with coccidia do not necessarily develop necrotic enteritis. This was evident in the piglets in litter no. 3. These piglets showed very high antibody titres to C. perfringens alpha toxin but no considerable clinical disease (data not shown). We assume that I. suis paves the way for extensive development of C. perfringens by inducing massive intestinal production of mucus which serves as a growing substrate for clostridia (Collier et al. 2008).
Conclusions
The present study clearly underpins the hypothesis that simultaneous infections with I. suis and C. perfringens type A induce strong interactions which may lead to a distinct increase in the severity of clinical disease in piglets, i.e. necrotic enteritis. Both I. suis and clostridia are widely distributed in piglet breeding units, and thus, co-infection appears to be a likely event in conventional herds. Anticoccidial treatment may thus not only protect piglets from parasite-induced lesions but may also help to prevent secondary necrotic enteritis.
References
Amann RI, Binder BJ, Olson RJ, Chisholm SW, Devereux RStahl DA (1990) Combination of 16s rRNA-targeted oligonucleotide probes with flow cytometry for analyzing mixed microbial populations. Appl Environ Microbiol 56:1919-1925
Amann RI, Ludwig W, Schleifer K-H (1995) Phylogenetic identification and in situ detection of individual microbial cells without cultivation. Microbiol Rev 59:143–169
Baba E, Ikemoto T, Fukata T, Sasai K, Arakawa A, McDougald LR (1997) Clostridial population and the intestinal lesions in chickens infected with Clostridium perfringens and Eimeria necatrix. Vet Microbiol 54:301–308
Bach U, Kalthoff V, Mundt HC, Popp A, Rinke M, Daugschies A, Luttge B (2003) Parasitological and morphological findings in porcine isosporosis after treatment with symmetrical triazintriones. Parasitol Res 91:27–33
Collier CT, Hofacre CL, Payne AM, Anderson DB, Kaiser P, Mackie RI, Gaskins HR (2008) Coccidia-induced mucogenesis promotes the onset of necrotic enteritis by supporting Clostridium perfringens growth. Vet Immunol Immunopath 122:104–115
Daugschies A, Bialek R, Joachim A, Mundt HC (2001) Autofluorescence microscopy for detection of nematode eggs and protozoa, in particular Isospora suis, in swine feces. Parasitol Res 87:409–412
Franks AH, Harmsen HJ, Raangs GC, Jansen GJ, Schut F, Welling GW (1998) Variations of bacterial populations in human feces measured by fluorescent in situ hybridization with group specific 16s rRNA-targeted oligonucleotide probes. Appl Environ Microbiol 64:3336-3345
Harleman JH, Meyer RC (1984) Life cycle of Isospora suis in gnotobiotic and conventionalized piglets. Vet Parasitol 17(1):27–39
Harleman JH, Meyer RC (1985) Pathogenicity of Isospora suis in gnotobiotic and conventionalised piglets. Vet Rec 116(21):561–565
Krüger M, Schroedl W, Krüger M, Schwarz S, Mengel H, Daugschies A, Swidsinski A, Mundt HC, Westphal B (2010) Prakt Tierarzt 91:774–784
Kuhnert Y, Schmäschke R, Daugschies A (2006) Comparison of different methods for examining the feces of suckling piglets for Isospora suis. Berl Munch Tierarztl Wochenschr 119(7–8):282–286
Maes D, Vyt P, Rabaeys P, Gevaert D (2007) Effects of toltrazuril on the growth of piglets in herds without clinical isosporosis. Vet J 173:197–199
Mavromatis I, Kyriakis CS, Alexopoulos C, Kritas SK, Tzika ED, Kyriakis SC. Efficacy and cost benefit study on the use of toltrazuril for the control of neonatal coccidiosis in pigs due to Isospora suis under field conditions. In: Proceedings of the 18th Congress of the International Pig Veterinary Society, Hamburg, Germany, 2004, 323
McDonel JL (1986) Toxins of Clostridium perfringens type A, B, C, D and E. In: Dorner F, Drews H (eds) Pharmacology of bacterial toxins. Pergamon, Oxford, pp 477–517
Mundt HC, Joachim A, Daugschies A, Zimmermann M (2003) Population biology studies on Isospora suis in piglets. Parasitol Res 90:158–159
Mundt HC, Joachim A, Becka M, Daugschies A (2006) Isospora suis: an experimental model for mammalian intestinal coccidiosis. Parasitol Res 98:167–175
Mundt HC, Mundt-Wüstenberg S, Daugschies A, Joachim A (2007) Efficacy of various anticoccidials against experimental porcine neonatal isosporosis. Parasitol Res 100:401–411
Niestrath M, Takla M, Joachim A, Daugschies A (2002) The role of Isospora suis as a pathogen in conventional piglet production in Germany. J Vet Med B Infect Dis Vet Public Health 49(4):176–180
Poulsen LK, Ballard G, Stahl DA (1993) Use of rRNA fluorescence in situ hybridization for measuring the activity of single cells in young and established biofilms. Appl Environ Microbiol 59:1354–1360
Robinson Y, Morin M, Girard C, Higgins R (1983) Experimental transmission of intestinal coccidiosis to piglets: clinical, parasitological and pathological findings. Can J Comp Med 47:401–407
Songer JG (1996) Clostridial enteric disease of domestic animals. Clin Microbiol Rev 9:216–234
Songer JG, Anderson MA (2006) Clostridium difficile: an important pathogen of food animals. Anaerobe 12(1):1–4
Songer JG, Uzal FA (2005) Clostridial enteric infections in pigs. J Vet Diagn Invest 17(6):528–536
Stuart BP, Lindsay DS, Ernst JV, Gosser HS (1980) Isospora suis enteritis in piglets. Vet Pathol 17(1):84–93
Swidsinski A, Loening-Baucke V, Lochs H, Hale LP (2005) Spatial organization of bacterial flora in normal and inflamed intestine: a fluorescence in situ hybridization study in mice. World J Gastroenterol 11:1131–1140
Vítovec J, Koudela B (1990) Double alteration of the small intestine in conventional and gnotobiotic piglets experimentally infected with the coccidium Isospora suis (Apicomplexa, Eimeriidae). Folia Parasitol (Praha) 37(1):21–33
Westphal B, Bernemann U, Kathmann L (2007) Isospora suis and Clostridium perfringens as mixed infection in suckling piglets just after post partum? Tierärztl Umschau 62:682
Williams RB, Marshall RN, La Regione RM, Catchpole J (2003) A new method for the experimental production of necrotic enteritis and its use for studies on the relationship between necrotic enteritis, coccidiosis and anticoccidial vaccination of chickens. Parasitol Res 90:19–26
Acknowledgements
Special thanks to Yvonne Kuhnert for the detection of excreted I. suis oocysts via epifluorescence microscopy.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Mengel, H., Kruger, M., Kruger, M.U. et al. Necrotic enteritis due to simultaneous infection with Isospora suis and clostridia in newborn piglets and its prevention by early treatment with toltrazuril. Parasitol Res 110, 1347–1355 (2012). https://doi.org/10.1007/s00436-011-2633-8
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00436-011-2633-8