
Abstract
This article presents morphological descriptions of larvae and adults of Hypopygiopsis tumrasvini Kurahashi (Diptera: Calliphoridae), a blow fly species that may be of forensic importance. Both second and third instar larvae are illustrated, with emphasis placed on important characteristic features used for larval identification, such as the cephalopharyngeal skeleton, anterior and posterior spiracles, and dorsal spines between the prothorax and mesothorax. Eight to 11 papillae were found on each anterior spiracle arranged in a single row. The dorsal spines between the prothorax and mesothorax were arranged in sets of posteriorly projecting acuminate spines with darkly pigmented tips. The posterior spiracles each bear three prominent and separated long, slender spiracular slits encircled by a dark, thick peritreme that is complete ventromedially around a button. Prominent inner projection of peritreme is seen between the middle and lower spiracular slits. A previously published key for differentiating third instar larvae of flies of possible forensic importance in Thailand is updated to include this additional species. Some characteristic features of males and females of the species are also provided.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Blow flies are the most common insect group found associated with human corpses; thus, they play an important role in forensic entomology throughout the world. In Southeast Asia, the primary blow fly genera known to contain species of forensic importance include Chrysomya, Achoetandrus, Lucilia, and Hemipyrellia (Lee et al. 2004; Sukontason et al. 2007); however, a blow fly species of the genus Hypopygiopsis has been recently reported. Both second and third instar larvae of Hypopygiopsis violacea Macquart were collected from a corpse found lying on the ground at a palm oil estate in Malaysia (Ahmad Firdaus et al. 2010). Therefore, Hypopygiopsis has now been added to the list of blow fly genera of forensic importance in Southeast Asia.
In the Oriental region, five species of Hypopygiopsis have been cataloged (Verves 2005). The documented species include: Hypopygiopsis fumipennis Walker, Hypopygiopsis infumata (Bigot), Hypopygiopsis tumrasvini Kurahashi, Hypopygiopsis robusta Malloch, and H. violacea. Among them, the first three species have been reported from Thailand (Kurahashi 1977; Verves 2005). During an ongoing survey for blow flies of possible forensic importance in Thailand from 2007–2010, in which day-old, tainted pork viscera was used as bait, H. tumrasvini and H. infumata were both collected along with several other blow fly species (unpublished data of authors). Because of the significance of possible forensic importance of these other species of Hypopygiopsis in the Oriental region, their biology should be investigated in order to provide baseline data in advance for anticipated use in forensic entomology. Due to the availability of specimens of the larvae and adults of H. tumrasvini, we therefore describe their morphology using light microscopy.
Materials and methods
The second and third instar larvae of H. tumrasvini used in this study were obtained from the F1 progeny of field-captured adult males and females from surveys using day-old, tainted pork viscera as bait in 2010. Specimens were removed from the rearing box and processed for morphological investigation. Two cuts were made on each specimen using a sharp blade so that parts could be positioned for optimal examination of important taxonomic characters. The first cut was made at the middle of the mesothorax to remove the anterior portion of the body to allow more suitable examination of the cephalopharyngeal skeleton, anterior spiracles, and dorsal spines between the prothorax and mesothorax. The second cut was made at the end of the eighth abdominal segment in order to remove the posterior portion of the body for optimal positioning to examine the posterior spiracles. The two removed parts were transferred into a well containing 10% KOH for 5 min, then transferred onto a clean glass slide into a few drops of Neo-Shigaral Permount® (Tokyo, Japan) and covered with a coverslip. Examination of morphological characters was performed under a light microscope, equipped with a calibrated eyepiece micrometer, using ×10 and ×40 magnifications (Olympus, Japan). Images were recorded with a digital camera (Nikon, Japan).
For examination of adult males and females, specimens were obtained from field collections. Specimens in the best condition were selected for taking photographs using the digital camera (Nikon, Japan). A few specimens of each sex were processed for illustration of their genitalia by cutting the posterior end (tergite 5 and sternite 5) from the rest of the abdomen under a dissection microscope. Each posterior end was placed in 70% alcohol for a few minutes and then transferred to 10% KOH for clearing (24 h for males and 12 h for females). Specimens were then placed on a glass depression slide containing 0.85% normal saline solution for dissecting. Dissected parts were subsequently transferred onto a clean glass slide with a few drops of Neo-Shigaral Permount® (Tokyo, Japan) and covered with a coverslip. Slide-mounted specimens were then examined under the light microscope (Olympus, Japan), and illustrations of male and female genitalia (ovipositor) were drawn from magnified images.
Results
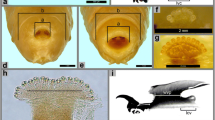
The larvae of this species exhibit the classical superficial appearance of most calliphorid larvae with a smooth, creamy white vermiform body composed of a head region lacking a head capsule, three thoracic segments, and eight abdominal segments (Figs. 1 and 2). Upon examination of the second instar under the light microscope, the cephalopharyngeal skeleton was found to lack an accessory sclerite ventral to the pair of mouth hooks (Fig. 1b). Eight to 11 papillae (n = 9) were found on each anterior spiracle arranged in a single row (Fig. 1c). Dorsal spines between the prothorax and mesothorax were found to be arranged in irregular rows with the tips of the spines greatly darkened (Fig. 1d). The posterior spiracles of second instar larvae each possess a light brown peritreme that was incomplete and only partially encircled the two completely separated, straight spiracular slits (Fig. 1e). A spiracular button was also absent in this instar.

Second instar larva of Hypopygiopsis tumrasvini. a Dorsal view of whole body, anterior end facing left. b Cephalopharyngeal skeleton. c Anterior spiracle showing 11 papillae. d Arrangement and shape of dorsal spines between prothorax and mesothorax. e Posterior spiracles each displaying incomplete peritreme encircling two completely separated, straight spiracular slits. Spiracular buttons are absent

Third instar larva of Hypopygiopsis tumrasvini. a Dorsal view of whole body, anterior end facing left. b Cephalopharyngeal skeleton with lack of accessory sclerite in position ventral to mouth hooks (arrow). c Anterior spiracle showing eight papillae. d Arrangement and shape of dorsal spines between prothorax and mesothorax revealing irregular rows of sets of posteriorly projecting acuminate spines with pigmented tips. e Posterior spiracles each displaying complete peritreme encircling three long, straight spiracular slits and ventromedially located button. Arrow indicates prominent inner projection of peritreme between middle and lower slits
The third instar larvae (Fig. 2a) were slightly larger with a median body length of 1.37 cm (range, 0.87–1.58 cm; n = 7). As was seen in the second instar larva, the cephalopharyngeal skeleton lacks an accessory sclerite ventral to the mouth hooks (Fig. 2b). The number of papillae on each anterior spiracle was similar with that of the second instar, with eight being displayed in Fig. 2c. The dorsal spines between the prothorax and mesothorax also appear in irregular rows, but in the third instar, these rows are arranged in sets of posteriorly projecting acuminate spines with darkly pigmented tips (Fig. 2d). The posterior spiracles each bear three prominent and separated long, slender spiracular slits encircled by a dark, thick peritreme that is complete ventromedially around a button (Fig. 2e). In addition, a prominent inner projection of peritreme is seen between the middle and lower spiracular slits (Fig. 2e, arrow).
A key for identification of third instar larvae of fly species of potential forensic importance in Thailand was published (Sukontason et al. 2004). The following key updates that key to include H. tumrasvini:
-
1.
Hairy body (with large tubercles along the body)—2
Non-hairy body (without large tubercles along the body)—3
-
2.
Small spines only at tips of tubercles; 9–12 papillae on each anterior spiracle—Achoetandrus rufifacies (Macquart)
Small spines along entire tubercles; 13–15 papillae on each anterior spiracle—Achoetandrus villeneuvi (Patton)
-
3.
Posterior spiracle with incomplete peritreme ventromedially—4
Posterior spiracle with complete peritreme ventromedially—5
-
4.
With large dark brown patch on dorsal surface; end of upper peritreme slightly expanded—Ceylonomyia nigripes (Aubertin)
Without large dark brown patch on dorsal surface; end of upper peritreme not expanded—Chrysomya megacephala (Fabricius) or Chrysomya pinguis (Walker)
-
5.
Sinuous spiracular slits; peritreme “D-shaped,” dark, and heavily sclerotized—6
Straight spiracular slits; peritreme not D-shaped, dark brown—7
-
6.
Sinuous spiracular slits “S-shaped”—Synthesiomyia nudiseta van der Wulp*
Sinuous spiracular slits “M-shaped”—Musca domestica L.*
-
7.
Slender larval shape; posterior spiracles close together; peritreme pale—Hydrotaea spinigera Stein*
Stout larval shape; posterior spiracles far apart; peritreme dark brown—8
-
8.
Spines between prothorax and mesothorax large; inner projection of peritreme between middle and lower spiracular slits not prominent; 4–7 papillae on each anterior spiracle—Lucilia cuprina (Wiedemann)
-
9.
Spines between prothorax and mesothorax small; inner projection of peritreme between middle and lower spiracular slits prominent; 5–11 papillae on each anterior spiracle—10
-
10.
Cephalopharyngeal skeleton with accessory sclerite; posterior spiracles far apart; 5–11 papillae on each anterior spiracle—Hemipyrellia ligurriens (Wiedemann)
Cephalopharyngeal skeleton without accessory sclerite; posterior spiracles close together; 8–11 papillae on each anterior spiracle—H. tumrasvini Kurahashi
*Muscidae
The adults of this fly are quite large, measuring ~9–15 mm in length. Important identifying characteristics of both sexes include the shining metallic bluish purple body with the distal portion shaded light brown (Fig. 3a, b) and the orange antennae, palpi, and parafacial region of the head (Fig. 3c, d). For comparative purposes, the ovipositor of the female is illustrated in Fig. 4a, while the male external genitalia of the species is shown in both caudal view (Fig. 4b) and lateral view (Fig. 4c). A lateral view of the aedeagus is also shown to compare to other species (Fig. 4d).

Adult stages of Hypopygiopsis tumrasvini. a Dorsal view of female. b Dorsal view of male. c Frontal view of female. d Frontal view of male

Genitalia of Hypopygiopsis tumrasvini. a Ovipositor. b Caudal view of male genitalia. c Lateral view of male genitalia. d Lateral view of aedeagus
Discussion
Proper identification of insect specimens found in human corpses is a necessary prerequisite before they can serve as useful entomological evidence in forensic investigations. Not only can adults be identified to species but third instar larvae of certain fly species can sometimes be identified on the basis of morphological characters when taxonomic keys are available. This is important in forensic investigations since this is often the most collected and abundant stage of flies in human corpses. Problems may sometimes arise during the rearing of larvae. For example, pupae may fail to produce adults in the laboratory due to damage by parasitoids. In this study, we found that the third instar larva of H. tumrasvini is very similar in morphology to that of H. violacea which was collected from a human corpse in Malaysia (Ahmad Firdaus et al. 2010). The accessory sclerite of the cephalopharyngeal skeleton was the only primary character found to differentiate between them, with this sclerite being absent in H. tumrasvini but present in H. violacea (Table 1). Although the spines between the prothorax and mesothorax of H. tumrasvini seem to be slightly different from the “intersegmental” spines described from H. violacea, we cannot draw conclusions from this comparison since the location of the specific site of the intersegmental spines was not designated in the description of the latter species (Ahmad Firdaus et al. 2010). The number of papillae on each anterior spiracle can occasionally serve to distinguish the two species (Table 1). Taxonomically, Hypopygiopsis is placed together with Hemipyrellia and Lucilia in the tribe Luciliini (Kurahashi and Chowanadisai 2001). Results of the present study indicate that the third instar larva of H. tumrasvini is similar in general morphology to the closely related blow fly, H. ligurriens (Wiedemann), a fly of forensic importance (Sukontason et al. 2007), but can be distinguished based on the key provided. Spines between the prothorax and mesothorax of H. tumrasvini were too similar to be distinguished from those previously reported from H. ligurriens (Sukontason et al. 2010). It can be inferred from previous results (Sukontason et al. 2004, 2010) and the present study that these spines between the prothorax and mesothorax of larvae of members of the tribe Luciliini are probably a unique feature to the group. However, the third instar larva of H. tumrasvini was more morphologically similar to H. ligurriens than to L. cuprina, of which the morphological distinction of these two latter species has already been published (Sukontason et al. 2010).
Only two characters were found to be useful in distinguishing between third instar larvae of H. tumrasvini and H. ligurriens: the accessory sclerite, which is absent in the former versus present in the latter, and the proximity of the pair of posterior spiracles to each other. However, if larvae are reared to adults, the two species can be easily identified based on adult morphological characters.
The original description of H. tumrasvini was based on adults and was first reported by Kurahashi (1977) who discovered it to be a new species from Thailand. The male external genitalia were also illustrated in this original description. Our study adds to the illustration of adult morphology of this fly species by providing figures of the female ovipositor. Moreover, color plates of adult males and females are provided to assist in confirmation of final identification of adults that have been reared from third instar larvae.
The ecological distribution of H. tumrasvini has been reported to be similar to H. violacea, with both species prevailing in dense tropical forests (Kurahashi 1977), although analyses from our own fly collections in combination with previous reports from Thailand (Tumrasvin et al. 1978), Malaysia, Singapore (Kurahashi et al. 1997), Cambodia, Laos, Vietnam (Kurahashi and Chowanadisai 2001), and the Philippines (Kurahashi and Magpayo 2000) reveal that the former species prevails in generally higher altitudes (349–1,700 m) than the latter species (30–1,100 m). Currently, H. violacea has not been recorded from Thailand (Kurahashi et al. 1997).
Conclusion
This present study highlights some important morphological features of the second and third instar larvae of H. tumrasvini, as well as illustrates genitalia and provides pictures of both adult males and females. A key to differentiate third instar larvae of fly species of possible forensic importance in Thailand was updated to include H. tumrasvini. This information should be useful to investigators in future forensic cases in Thailand that involve this species of blow fly.
References
Ahmad Firdaus MS, Marwi MA, Syamsa RA, Zuha RM, Ikhwan Z, Omar B (2010) Morphological descriptions of second and third instar larvae of Hypopygiopsis violacea Macquart (Diptera: Calliphoridae), a forensically important fly in Malaysia. Trop Biomed 27:134–137
Kurahashi H (1977) The tribe Luciliini from Australian and Oriental regions I. Genus Hypopygiopsis Townsend (Diptera, Calliphoridae). Kontyu 45:553–562
Kurahashi H, Chowanadisai L (2001) Blow flies (Insecta: Diptera: Calliphoridae) from Indochina. Species Divers 6:185–242
Kurahashi H, Magpayo FR (2000) Blow flies (Insecta: Diptera: Calliphoridae) of the Philippines. Raffles Bull Zool Suppl 9:1–78
Kurahashi H, Benjaphong N, Omar B (1997) Blow flies (Insecta: Diptera: Calliphoridae) of Malaysia and Singapore. Raffles Bull Zool Suppl 5:1–88
Lee HL, Krishnasamy M, Abdullah AG, Jeffery J (2004) Review of forensically important entomological specimens in the period of 1972–2002. Trop Biomed 21:69–75
Sukontason K, Sukontason KL, Ngern-klun R, Sripakdee D, Piangjai S (2004) Differentiation of the third instar of forensically important fly species in Thailand. Ann Entomol Soc Am 97:1069–1075
Sukontason K, Narongchai P, Kanchai C, Vichairat K, Sribanditmongkol P, Bhoopat T, Kurahashi H, Chockjamsai M, Piangjai S, Bunchu N, Vongvivach S, Samai W, Chaiwong T, Methanitikorn R, Ngern-klun R, Sripakdee D, Boonsriwong W, Siriwattanarungsee S, Srimuangwong C, Hanterdsith B, Chaiwan K, Srisuwan C, Upakut S, Moopayak K, Vogtsberger RC, Olson JK, Sukontason KL (2007) Forensic entomology cases in Thailand: a review of cases from 2000 to 2006. Parasitol Res 101:1417–1423
Sukontason K, Sribanditmongkol P, Ngoen-klan R, Klong-klaew T, Moophayak K, Sukontason KL (2010) Differentiation between Lucilia cuprina and Hemipyrellia ligurriens (Diptera: Calliphoridae) larvae for use in forensic entomology application. Parasitol Res 106:641–646
Tumrasvin W, Sucharit S, Kano R (1978) Studies on medically important flies in Thailand. IV. Altitudinal distribution of flies belonging to Muscidae and Calliphoridae in Doi Indhanondh Mountain, Chiengmai, in early summer season. Bull Tokyo Med Dent Univ 25:77–81
Verves YG (2005) A catalogue of Oriental Calliphoridae (Diptera). Int J Dipterol Res 16:233–310
Acknowledgments
We thank the Faculty of Medicine, Chiang Mai University and Thailand Research Fund for the support of this project.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Moophayak, K., Sa-nit, Sn., Sukontason, K. et al. Morphological descriptions for the identification of Hypopygiopsis tumrasvini Kurahashi (Diptera: Calliphoridae). Parasitol Res 109, 1323–1328 (2011). https://doi.org/10.1007/s00436-011-2424-2
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00436-011-2424-2