
Abstract
During a parasitological survey of two non-native fish species—racer goby Neogobius gymnotrachelus and monkey goby Neogobius fluviatilis—in the Włocławek Reservoir on the lower Vistula River, the monogenean Gyrodactylus proterorhini was recorded for the first time in Poland, and for the first time, the racer goby was listed as the parasite host. Specimens of G. proterorhini were detected on body surface, fins, and gills of the fish studied. In 2006, the monkey goby was infected with the prevalence 41%, racer goby with the prevalence of 47%, at maximal intensity of 6 and 9 parasites per fish, (respectively). The infection level in relation to the fish size and sampling season was also discussed. The presented study supports the hypothesis of progressive introduction of the parasite with gobiids to the colonized areas.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
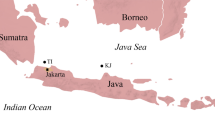
The Włocławek Reservoir on the lower course of the Vistula River (Fig. 1) was recently colonized by three species of Ponto–Caspian goby fish. Racer goby Neogobius gymnotrachelus (Kessler, 1857) has been noted there since 2000 (Kostrzewa and Grabowski 2001) and Monkey goby Neogobius fluviatilis (Pallas, 1814) since 2002 (Kostrzewa and Grabowski 2002). Quite recently in 2008, tubenose goby Proterorhinus marmoratus (Pallas, 1814) has appeared in the lower Vistula River course near Plock (Grabowska et al. 2008). Subsequently, it has been found in fish catches in the central part of the Włocławek Reservoir, but described as P. semilunaris (Heckel, 1837) (Piotr Hliwa unpublished). The natural range of the above-mentioned gobiids is Eurasia; rivers and estuaries of Black, Azov, and Caspian Seas. Proterorhinus spp. also occur in the Mediterranean area—in the Evros and Strymon systems, which empty in the northern Aegean Sea (Kvach and Oğuz 2009). Those three fish species reached the Vistula basin and Poland probably through the Pripyat-Bug canal commonly crossed by Ponto–Caspian organisms on their way to Europe (Bij de Vaate et al. 2002; Grabowska et al. 2008). Goby fish have become a numerous species in the near-shore fish communities in the lower Vistula and stimulated considerable changes in the local food webs (Kakareko et al. 2003; 2005; 2009; Grabowska and Grabowski 2005; Grabowska et al. 2009).

Sampling site (S) of gobiids in the Włocławek Reservoir (WR) and a map of Poland, with the Vistula River (VR) position shown
Non-native species could introduce exotic parasites to the colonized areas. Monitoring of this “secondary” introduction (in the meaning of symbiotic species introduction with their introduced host) is essential in predicting the effects of biological invasions, due to the potential susceptibility of native fish species for infection with an alien parasites, or indirectly, due to changes in the communities structure of native parasites (Torchin et al. 2003; Torchin and Mitchell 2004).
Gyrodactylus proterorhini Ergens 1967 [syns G. gussevi Najdenova, 1966; G. najdenovae Malmberg 1970, and G. arcuatus Bykhovskii sensu Ergens] is a common parasite of gobiids inhabiting a littoral zone of the Black and Azov Seas and their estuaries (Ergens 1972). The species was initially described by Ergens (1967) from the tubenose goby Proterorhinus marmoratus in its natural range in the Danube River (in southern Slovakia). Formerly, G. proterorhini was incorrectly recognized as G. arcuatus from the same fish host in the Czechoslovakian part of the Danube, i.e., outside the natural range of the species (Ergens 1962). Afterwards, four more species of Gobiidae in their natural range were mentioned as the G. proterorhini hosts: grass goby Zosterisessor ophiocephalus (formerly Gobius ophiocephalus), giant goby Gobius cobitis, black goby G. niger, and round goby Neogobius melanostomus (Najdenova 1974; Ergens 1985; Dmitrieva and Dimitrov 2002; Longshaw et al. 2003; Harris et al. 2004; Őzer 2007). Miroshnichenko (2008) adds three further species: monkey goby N. fluviatilis, flatsnout goby N. platyrostris, and knout goby Mesogobius batrachocephalus to this pool. Study of the introduction of G. proterorhini broadens the list of the parasite hosts to one more (the eighth)—bighead goby Neogobius kessleri, infected with the parasite in the New Danube channel (at the Slovak stretch of Danube) and in the Horn River. The infection of bighead goby was probably related to co-occurrence with tubenose goby at these particular study sites (Ondračková et al. 2005; 2009).
Francová (2007) has confirmed the G. proterorhini introduction with round goby in the stretch of the Danube in Slovakia and Austria. However, the round goby in Polish coastal waters of the Baltic Sea (the area invaded in recent decades) is not infected with Monogenea at all (Rolbiecki 2006). Accordingly, the present paper reports for the first time the occurrence of G. proterorhini in Poland, as well as for the first time in racer goby N. gymnotrachelus. It is worth noting that this species of monogeneans is common for both: the freshwater and the marine tubenose goby (Kvach and Oğuz 2009). The freshwater Proterorhinus semilunaris, liberated from the species of P. marmoratus on the basis of genetic investigations (see Stepien and Tumeo 2006), is expanding its range widely at present (Manné and Poulet 2008), also in the Vistula River and the Włocławek Reservoir, but till now, there has been no information on G. proterorhini introduction to the areas colonized with this fish host (Kvach and Stepien 2008). The population dynamics of the parasite during the widening the list of its hosts and the constant increase of their numbers in the Włocławek Reservoir are interesting and are ongoing.
Material and methods
Specimens of racer goby and monkey goby were collected in the Włocławek Reservoir (central Poland) by electrofishing (battery-powered unit IUP-12, RADET Ltd., Poznań, 225/300 V, 6A, 50–90 Hz) close to Dobiegniewo locality (52° 37–38′ N; 19° 19–20′ E) (Fig. 1) in the shallow (up to 1 m deep) near shore zone of the flooded area of the reservoir. Fish samples were collected three times in 2006: in spring (29th of May), in summer (7th of August), and in autumn (2nd of October). The number of racer goby caught 10, 9, and 10, (respectively in seasons) was larger than number of monkey goby in related samples: 4, 4, and 9. All individuals were measured, weighed, and analyzed for the presence of monogeneans on their gills, fins, and body surface. Monogeneans were removed and fixed with a mixture of glycerine ammonium picrate (GAP) (Malmberg 1970). Smears of scrapings stained with Giemsa dye were also analyzed. Specimen identification was based on morphological analysis and measurements of the haptor's hard elements according to Gusev (1985) under a compound light microscope (Nikon ECLIPSE E600W).
Prevalence (percentage of the fish infected), intensity of infection (mean value and range of the parasite number per the infected fish), and abundance (mean number of the parasite per fish studied) were estimated to characterize the infection level. The differences in the fish size, as well as the differences in the infection level in samples, were compared by the Mann–Whitney's U Test. Spearmann's correlation coefficient was applied to find the relations between the fish size (standard length (SL), total length (TL), and weight) and the infection level (number of G. proterorhini per fish studied, both infected and non-infected individuals were included).
Results
In the Włocławek Reservoir, 41% of racer goby and 47% of monkey goby analyzed in 2006 were infected with Gyrodactylus proterorhini. Specimens of the parasite were detected predominantly on fins and body surface of racer goby (18 out of the 25 specimens) and almost exclusively on gills of monkey goby (22 out of the 23 specimens). The prevalence in particular seasons ranged from 0% to 80% and from 25% to 75% in racer goby and monkey goby, respectively (Table 1). According to the U test, the infection of racer goby was significantly higher in spring (U = 10, z = −3.02, p = 0.002) and in summer (U = 25, z = −2.30, p = 0.022) than in autumn (Table 1). The same test, however, did not confirm the significance (p > 0.05) of seasonal differences in the infection level of monkey goby (Table 1). A non-parametric Spearman's test revealed a weak positive correlation between the infection level and the body length of racer goby (sl/number of G. proterorhini R = 0.22, p < 0.05), but did not confirm such a dependence for monkey goby at all (p > 0.05). Individuals of racer goby studied in autumn were significantly smaller than those analyzed in the spring (U = 15, z = 2.65, p = 0.008). The sizes of monkey goby in seasonal samples were comparable (U test at p > 0.05) (Fig. 2).

Comparison of the fish total length (TL) in samples by Mann–Whitney U test. The fish sampling carried out three times (seasonally): in 2006 (29th of May, 7th of August, and 2nd of October). Individuals of racer goby (N. gymnotrachelus) in sample of autumn significantly smaller than those in sample of spring (p < 0.05)
Specimen description
Monogeneans detected in round goby and monkey goby were similar. The measurements of haptoral hard parts (Fig. 3) (48 specimens included): anchors with additional processes (Fig. 3a)—the total length of anchor 40–44 μm, the length of shaft 30–35 μm, the length of the root 9–14 μm, and of the point 20–22 μm. Ventral bar with small anterolateral processes (Fig. 3a)—the length of ventral bar 5 μm, the width 18 μm, the length of membrane 12 μm. Dorsal bar (Fig. 3a)—the length 2 μm, the width 19 μm. Marginal hooks (Fig. 3b)—total length 22–24 μm, the length of sickle 4 μm. The measurements of monogenean body (15 best-preserved specimens included): total length 330–370 μm and the width 70–90 μm

Haptoral hard parts of Gyrodactylus proterorhini. Specimen detected in racer goby from the Włocławek Reservoir (Vistula River, Poland) fixed in GAP. a anchors linked with ventral and dorsal bars, b marginal hooks
Discussion
Significant seasonal differences in the occurrence of G. proterorhini were not observed in monkey goby but were observed in racer goby (Table 1). This last difference could be related to the significant difference observed in racer goby size collected in different seasons (Fig. 2). The relationship between levels of infection and host size is, however, not completely conclusive in the present work as the Spearman's test indicate no correlation in monkey goby (p > 0.05) but correlation on racer goby (R = 0.22, p < 0.05). All these results should be confirmed in a near future with the observation of a much bigger number of fish host in all seasons.
At natural range, depending on the area of study and the fish species, the infection of Gobiidae with G. proterorhini varies substantially. In the Black Sea near Sevastopol, the extremely high intensity of infection up to 500, 320, and 98 parasites per fish was observed (Najdenova 1974) in Gobius cobitis, G. niger and G. opiocephalus (respectively). In the Azov Sea, maximal intensity in Neogobius fluviatilis reached 165. However, the marine Proterorhinus marmoratus, as well as the freshwater tubenose goby P. semilunaris, were infected with relatively low abundance in summer in the Sukhyi Estuary, Gulf of Odessa, and Dniester River did not exceed 0.3 (Kvach and Oğuz 2009).
In the colonized area (the Slovak section of the Danube River), the abundance of G. proterorhini in bighead goby Neogobius kessleri varied between 0.03 in autumn and 13.25 in spring, at a prevalence of 17% and 90%, respectively (in seasons) (Ondračková et al. 2005; 2009).
In general, the infection of gobiids with G. proterorhini in the Włocławek Reservoir was relatively low. The prevalence, as predicted (Chubb 1977), picked in spring, achieving 80% and 75%, but the abundance did not exceed 2.3 parasites per fish, even in warmer months (Table 1). However, the population growth of the parasite during the progressive expansion of its hosts is highly probable and expected. The specific behavior of gobiids during the spawning season: the accumulation of fish in coastal zones, the high concentration of larvae in the spawning grounds, and the quick transmission of monogeneans from males guarding the nest directly to eggs and to offspring, all support the infection process. As the laboratory study proved, the incubated eggs are infected with monogeneans a week after laying, and the larvae are then invaded by the parasite just after hatching (Najdenova 1974). The infection with monogeneans is also reinforced by an increase in temperature during the spawning season and by the reduced immunity of fish after overwintering (Chubb 1977; Appleby 1996).
The occurrence of G. proterorhini in the lower course of the Vistula River supports the hypothesis of the progressive introduction of the parasite with gobiids to the colonized areas. This species of monogenea is currently becoming a permanent element of parasite communities in Central and Western Europe (Ondračková et al. 2005; 2009; Francová 2007; Kvach and Oğuz 2009), and its hosts are becoming a permanent component of the ichthyofauna in these areas.
References
Appleby C (1996) Population dynamics of Gyrodactylus sp. (Monogenea) infecting the sand goby in the Oslo Fjord, Norway. J Fish Biol 49:402–410
Bij de Vaate A, Jażdżewski K, Ketelaars HAM, Gollasch S, Van der Velde G (2002) Geographical patterns in range extension of Ponto-Caspian macroinvertebrate species in Europe. Can J Fish Aquat Sci 59:1159–1174
Chubb JC (1977) Seasonal occurrence of helminths in freshwater fishes. Part 1. Monogenea. Adv Parasitol 15:133–199
Dmitrieva EV, Dimitrov G (2002) Variability in the taxonomic characters of Black Sea gyrodactylids (Monogenea). Syst Parasitol 51:199–206
Ergens R (1962) Further new species of the genus Gyrodactylus Nordmann, 1832 (Monogenoidea) in fauna of Czechoslovakia. Folia Zool 11:327–331
Ergens R (1967) New species of the genus Gyrodactylus (Monogenoidea) from the Danube Basin. Folia Parasitol (Praha) 14:377–379
Ergens R (1972) Nomenclatoric problems of Gyrodactylus gussevi Najdenova, 1966 and G. proterorhini Ergens, 1967 (Monogenoidea, Gyrodactylidae). Folia Parasitol 19:333
Ergens R (1985) Order Gyrodactylidea Bychowsky, 1937. In: Gusev AV (ed) Key to the parasites of the freshwater fishfauna of the USSR. Leningrad Nauka Vol 2:269–347
Francová K (2007) Diversity of parasites of the invasive species of fish Neogobius melanostomus (Pallas, 1814) in the Danube. Dissertation, Masaryk University, Brno
Grabowska J, Grabowski M (2005) Diel feeding activity in early summer of racer goby Neogobius gymnotrachelus (Gobiidae): a new invader in the Baltic basin. J Appl Ichthyol 21:282–286
Grabowska J, Pietraszewski D, Ondračková M (2008) Tubenose goby Proterorhinus marmoratus (Pallas, 1814) has joined three other Ponto–Caspian gobies in the Vistula River (Poland). Aquat Inv 3:261–265
Grabowska J, Grabowski M, Kostecka A (2009) Diet and feeding habits of monkey goby (Neogobius fluviatilis) in a newly invaded area. Biol Invasions 11:2161–2170
Gusev AV (1985) Parasitic Metazoa. In: Bauer ON (ed) Key to parasites of freshwater fishes of SSSR vol 2. Publishing House Nauka, Leningrad
Harris PD, Shinn AP, Cable J, Bakke TA (2004) Nominal species of the genus Gyrodactylus von Nordmann 1832 (Monogenea: Gyrodactylidae), with a list of principal host species. Syst Parasitol 59:1–27
Kakareko T, Płąchocki D, Kobak J (2009) Relative abundance of Ponto–Caspian gobiids in the lower Vistula River (Poland) 3- to 4 years after appearance. J Appl Ichthyol 25:647–651
Kakareko T, Żytkowicz J, Kowalska M (2003) Preliminary studies on trophic relationships between the invasive gobiid of genus Neogobius, and small percids in the littoral zone of the Włocławek Reservoir. Suppl Acta Hydrobiol 6:29–38
Kakareko T, Żbikowski J, Żytkowicz J (2005) Diet partitioning in summer of two syntopic neogobiids from two different habitats of the lower Vistula River, Poland. J Appl Ichthyol 21:292–295
Kostrzewa J, Grabowski M (2001) Racer goby (naked-head), Neogobius gymnotrachelus (Kessler, 1857) (Gobiidae, Perciformes)—the new fish species in the Vistula River. Przegl Zool 45:101–102
Kostrzewa J, Grabowski M (2002) Monkey goby Neogobius fluviatilis (Pallas, 1811) in the Vistula River—phenomenon of the invasion of Ponto–Caspian Gobiidae. Przegl Zool 46:235–242
Kvach Y, Stepien C (2008) Metazoan parasites of introduced round and tubenose gobies in the Great Lakes: support for the “Enemy Release Hypothesis”. J Great Lakes Res 34:23–35
Kvach Y, Oğuz MC (2009) Communities of metazoan parasites of two fishes of the Proterorhinus genus (Actinopterygii: Gobiidae). Helminthologia 46:168–176
Longshaw M, Pursglove M, Shinn AP (2003) Gyrodactylus quadratidigitus n. sp. (Monogenea: Gyrodactylidae), a parasite of the leopard-spotted goby Thorogobius ephippiatus (Lowe) from the south-western coast of the UK. Syst Parasitol 55:151–157
Malmberg G (1970) The excretory systems and marginal hooks as a basis for the systematics of Gyrodactylus (Trematoda, Monogenea). Arkiv Zool 2:1–235
Manné S, Poulet N (2008) First record of the western tubenose goby Proterorhinus semilunaris (Heckel, 1837) in France. Knowl Managt Aquatic Ecosyst 389:03. doi:10.1051/kmae:2008009
Miroshnichenko AI (2008) Monogenea. In: Lisitsina OI, Miroshnichenko AI (eds) Catalogue of Helminths of Ukraine Acanthocephala. National Science Academy of Ukraine, Kiev
Najdenova NN (1974) Parasite fauna of fishes of the goby family from the Black and Azov seas. Naukova Dumka, Kiev
Ondračkova M, Dávidová M, Pečínková M, Blažek R, Gelnar M, Valová Z, Černý J, Jurajda P (2005) Metazoan parasites of Neogobius fishes in the Slovak section of the River Danube. J Appl Ichthyol 21:345–349
Ondračková M, Dávidová M, Blažek R, Gelnar M, Jurajda P (2009) The interaction between an introduced fish host and local parasite fauna: Neogobius kessleri in the middle Danube River. Parasitol Res 105:201–208
Őzer A (2007) Metazoan parasite fauna of the round goby Neogobius melanostomus Pallas, 1811 (Perciformes: Gobiidae) collected from the Black Sea coast at Sinop, Turkey. J Nat Hist 41:483–492
Rolbiecki L (2006) Parasites of the round goby, Neogobius melanostomus (Pallas, 1811), an invasive species in the Polish fauna of the Vistula Lagoon ecosystem. Oceanologia 48:545–561
Stepien CA, Tumeo MA (2006) Invasion genetics of Ponto-Caspian gobies in the Great Lakes: a ‘cryptic’species, absence of founder effects, and comparative risk analysis. Biol Inv 8:61–78
Torchin ME, Lafferty KD, Dobson A, McKenzie VJ, Kuris AM (2003) Introduced species and their missing parasites. Nature 421:628–630
Torchin ME, Mitchell CE (2004) Parasites, pathogens, and invasions by plants and animals. Front Ecol Environ 2:183–190
Acknowledgments
This study was supported by Warmia and Mazury University in Olsztyn, Poland, project number 0802.0201, and by Polish Ministry of Science and Higher Education, grant number N N304 027436
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Mierzejewska, K., Martyniak, A., Kakareko, T. et al. Gyrodactylus proterorhini Ergens, 1967 (Monogenoidea, Gyrodactylidae) in gobiids from the Vistula River—the first record of the parasite in Poland. Parasitol Res 108, 1147–1151 (2011). https://doi.org/10.1007/s00436-010-2175-5
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00436-010-2175-5