
Abstract
The estuarine-dependent marine fish includes marine species that inhabit, at some stage of their life cycle, the Mar Chiquita coastal lagoon, Buenos Aires Province, Argentina. In the present study, we examined for parasites a total of 209 fishes belonging to following the estuarine-dependent marine species: Brevoortia aurea, Odontesthes argentinensis, Mugil platanus, Paralichthys orbignyanus, Micropogonias furnieri, and Pogonias cromis. The parasite fauna in the six host species comprised 43 parasite species, the majority of which represented new locality and new host records. A high percentage of fish was parasitized with at least one parasite species. The greatest values of prevalence were registered for M. furnieri, B. aurea, and P. cromis, at the component community level, followed by P. orbignyanus, O. argentinensis, and M. platanus. On the other hand, B. aurea, O. argentinensis, M. furnieri, and P. cromis showed similar percentages of both larval and adult stages of parasites. In the case of M. platanus, adult stages dominated the total number of parasites, whereas P. orbignyanus harbored mainly larval stages. Out of the six fish species herein studied, M. platanus seems to generally act only as definitive host in the local parasite’s life cycle. From a parasitological point of view, the expression “estuarine-dependent marine fishes” remains valid, although the contribution of the fish species in the lagoon to the maintenance of parasite populations is relatively minor.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Mar Chiquita coastal lagoon is a World Reserve of Biosphere since 1996 and is located in the Buenos Aires Province, Argentina (37°32′ S, 57°19′ W). The interaction between freshwater and saltwater is a permanent but irregular phenomenon due to the combined effects of wind, rain, and tides. Therefore, the lagoon can be divided into a freshwater zone, characterized by continental water discharge without tidal effects, and an estuarine zone which communicates with the open sea. The estuarine zone is characterized by mixo-euryhaline waters and is greatly influenced by marine water (Reta et al. 2001).
The fauna of the lagoon consists of freshwater, marine, estuarine, and migratory organisms which are adapted to the local variations in salinity and water levels (Olivier et al. 1972; Cousseau et al. 2001). More than 30 fish species with different bioecological characteristics among them—the estuarine-dependent marine fishes—include marine species which are predominantly found in lagoons at some stage of their life cycle (Moyle and Cech 2004). These species reproduce offshore but utilize estuarine shallow waters as either nursery grounds, feeding areas, or overwintering refuge (Cousseau et al. 2001). The most representative species in this category are Brevoortia aurea Spix et Agassiz, 1829 (Brazilian menhaden); Odontesthes argentinensis Valenciennes, 1835 (silver side); Mugil platanus Günther, 1880 (grey mullet); Paralichthys orbignyanus Valenciennes, 1839 (flounder); Micropogonias furnieri Desmarest, 1823 (whitemouth croaker); and Pogonias cromis Linnaeus, 1766 (black drum). The available literature on parasites in vertebrate hosts from Mar Chiquita lagoon consists almost exclusively of systematic studies on birds, whereas fish parasites have received relatively little attention (Etchegoin et al. 1996; Etchegoin and Martorelli 1997a, b; Cremonte et al. 1999a, b; Etchegoin et al. 2000; Navone et al. 2000). Taking into account the special features of the lagoon as environment as well as the lack of information on fish parasites in this lagoon, further studies of fish parasites are needed to shed light on the role of these fish hosts in the parasitic transmission chains in this region. The aims of the present study are (1) to improve the knowledge on parasite diversity in six species of estuarine-dependent marine fishes in the lagoon and to characterize quantitatively their parasite populations and communities and (2) to establish the possible influence of the lagoon on the composition of parasite assemblages in these fish hosts and their possible contribution to the parasite’s life cycle in the area, analyzing the relationships between the presence of parasites and the diet and ecological habits of their fish hosts.
Materials and methods
The species of estuarine-dependent marine fishes selected for this study were B. aurea, O. argentinensis, M. platanus, P. orbignyanus, M. furnieri, and P. cromis. Fishes examined (n = 209; Table 1) were caught monthly between January 2004 and June 2008 near the Mar Chiquita lagoon mouth (Fig. 1). Fishes were captured by using different nets and transported to the laboratory for examination immediately after capture. All the fishes examined were adults, and the total length mean ± SD (in millimeters) for each host species was B. aurea (363 ± 42.74), O. argentinensis (339 ± 61.05), M. platanus (463 ± 62.98), P. orbignyanus (398 ± 117.36), M. furnieri (348 ± 101.73), and P. cromis (510 ± 114.60). The branchial cavities, the external surface, and the viscera of each fish were removed and examined under a stereoscopic microscope. Ecto- and endoparasites were recovered and quantified according to a standardized protocol. At the population level, prevalence (P), mean intensity (MI), and mean abundance (MA) according to Bush et al. (1997; with 95% bootstrap confidence intervals for the MI and MA, following Rózsa et al. 2000) were calculated for each parasite species in each host species using Quantitative Parasitology 3.0 (Reiczigel and Rózsa 2005). At the infracommunity level, the total mean number of parasites (n), the species richness (s), the Brillouin’s diversity index because each infracommunity was fully censused (Magurran 2005), and the evenness index (Brillouin’s index; E) to investigate the uniformity in the parasite species distribution in each infracommunity were calculated. At the component community level, total prevalence, total number of parasites (N), the species richness (S), and the Berger–Parker species dominance index (D) to know the proportion of dominant species individuals with respect to the total individuals in each component community were also calculated. In order to determine the role of fishes as intermediate or definitive hosts in the parasite’s life cycle, the relationships of species richness between larval and adult stages were calculated and expressed as percentage.

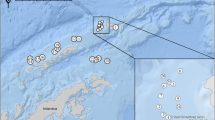
Map of Mar Chiquita coastal lagoon showing the sampling zone
Results
Component communities
The parasite fauna of the six estuarine-dependent marine fishes comprised 43 species belonging to eight taxa: Copepoda (six), Monogenea (four), Digenea (nine), Aspidogastrea (one), Nematoda (twelve), Acanthocephala (six), Cestoda (four), and Hirudinea (one). The majority of the parasite species found during this study represented both new locality and host records (Table 3). A high percentage of fish species was parasitized by, at least, one parasite species. The greatest values of prevalence were registered for M. furnieri, B. aurea, and P. cromis, with percentages higher than 90% at the component community level and followed, in order of importance, by P. orbignyanus, O. argentinensis, and M. platanus (Table 1).
Dominance
Different parasite species were dominant in each host species. In B. aurea, one parasite, the monogenean Mazocraeoides georgei, showed extreme dominance (Fig. 2a), whereas four of the host species (M. platanus, P. orbignyanus, M. furnieri, and P. cromis) presented one dominant specie with similar values ranging from 0.6 to 0.7 in the component community (Fig. 2c–f). On the other hand, in O. argentinensis, the dominant species were the Echinostome-type metacercariae and Profilicollis chasmagnathi (Fig. 2b).

Berger–Parker dominance index (D) calculated for the parasites component communities in the six host species
Parasite’s life cycles
With respect to the role of fishes in the parasite’s life cycles, four host species (B. aurea, O. argentinensis, M. furnieri, and P. cromis) harbored similar percentages of both larval and adult stages of parasites (Fig. 3). In the case of M. platanus, adult stages represented 83% of the parasite species richness, whereas P. orbignyanus harbored mainly larval stages (67% of the parasite species richness) (Fig. 3).

Percentage of larval and adult stages of parasite species in each host species
Infracommunities
Comparisons of parasitic indices for the six hosts revealed that P. orbignyanus presented infracommunities with higher values parasites number as well as species richness. On the other hand, the diversity in the infracommunities of this host was similar to M. furnieri. As regards parasite species mean richness, M. furnieri showed the highest value, followed by flounder and by the rest of fishes with similar values. The greatest evenness values were registered for the infracommunities of M. platanus, P. cromis, B. aurea, and M. furnieri, followed by P. orbignyanus and O. argentinensis with values lower than 0.55 (Table 2).
Populations
In general terms, the acanthocephalans P. chasmagnathi (in O. argentinensis, P. orbignyanus, M. furnieri, and P. cromis) and Floridosentis mugilis (in M. platanus), the nematodes Cucullanus marplatensis (in O. argentinensis), Contracaecum sp. (in P. orbignyanus), and Dichelyne sciaenidicola and Dichelyne mariajuliae (in M. furnieri and P. cromis, respectively) showed the highest values of prevalence (Table 3). On the other hand, the Echinostome-type metacercariae (in O. argentinensis) and the adult digeneans Dicrogaster fastigatus (in M. platanus), Bacciger microacetabularis, and Prosorhynchoides labiatus (in P. orbignyanus) registered the greatest values of mean intensity and abundance. Furthermore, in the latter host, the Grillotia sp. tapeworm showed high values of mean intensity and abundance (Table 3). In B. aurea, one monogean species (M. georgei) registered high values of prevalence, mean intensity, and abundance values (Table 3).
Discussion
Among all the different host features, the feeding habits can influence the number of parasite species exploiting a fish host species (Luque et al. 2004; Poulin 2007). Thus, the diet of the different hosts is a main factor affecting the parasite community structure, in the case of parasites transmitted to definitive hosts through prey–predator relationships, e.g., the digeneans (Sasal et al. 1999). Consequently, predatory fishes should be exposed to a higher number of infective stages of parasites, in contrast with planktivorous fishes. Parasite communities of benthic fishes (with broader diets) should be composed of a higher number of species than pelagic fishes (which tend to have a more specialized diet) (Luque and Poulin 2004). Also, fishes presenting broader geographical distributions (with access to a greater depth range) and ability to enter brackish waters should harbor more helminths than those with more restricted distributions, simply because of the result of the greater variety of prey they must feed on (Marcogliese 2002, 2003; Chambers and Dick 2005; Klimpel et al. 2006). In addition, the ecology of fish in brackish waters allows the interchange of parasite species among freshwater and marine fish species. The reason for this interchange is due that marine parasite species utilize both freshwater and marine fish species as intermediate or final hosts, and marine fish can harbor freshwater parasites species (Valtonen et al. 2001).
In the present study, the fishes equitably played the roles of intermediate and definitive hosts, with the exception of M. platanus which might be regarded mainly as definitive host. This assumption is supported by the fact that five out of the six species of parasites harbored by M. platanus were present at adult stages (including the haploporid digenean D. fastigatus, the monogenean Metamicrocotyla macracantha, and the copepod Ergasilus versicolor). The digenean D. fastigatus presents an abbreviated life cycle involving only one intermediate host (a snail), and fish infection occurs via direct ingestion of encysted metacercariae found in the sediment of the lagoon. Martorelli (1989a) and Etchegoin and Martorelli (1998) described three morphological types of cercariae belonging to the Haploporidae family parasitizing the Heleobia conexa and Heleobia australis gastropods from Mar Chiquita coastal lagoon, one of which might be D. fastigatus. On the other hand, the monogenean M. macracantha and the copepod E. versicolor have direct life cycles. Therefore, out of the six estuarine-dependent marine fishes under study, grey mullet may be the only host that, because of their prolonged incursions into the lagoon, becomes part of the local parasites’ life cycles (e.g., D. fastigatus). In fact, Mar Chiquita coastal lagoon is considered a nursery ground for M. platanus (Cousseau et al. 2001; Cousseau and Perrota 2004; González Castro 2007). The juveniles enter into the lagoon from the sea, where they undergo morphophysiological changes in their digestive system causing a diet change from zooplanktophagous to herbivorous or detritivorous feeding (Acha 1990). At this stage, the mullet starts feeding on plant material obtained by grubbing through bottom detritus and they even start the gonadal maturation (González Castro 2007; González Castro et al. 2009a, b). Then, the reproductive migration from the lagoon toward the sea occurs and when finished, grey mullets return to the lagoon and start to feed (Cousseau et al. 2001; Cousseau and Perrota 2004; González Castro 2007; González Castro et al. 2009a, b). It is worth noting the low parasite diversity found in M. platanus in relation to other species of this genus at a worldwide level (García and Williams 1985; Fernández 1987; Knoff et al. 1997). The low parasite diversity of grey mullet from Mar Chiquita might be due to the detritivorous feeding habits of this fish in the lagoon that limit the acquisition of parasites larval stages harbored mainly by invertebrates. In fact, the parasite species with highest values of prevalence and intensity in the parasite component community of M. platanus were the digenean D. fastigatus (transmitted to fish directly by ingestion of encysted metacercariae present in the sediments), the monogenean M. macracantha, and the copepod E. versicolor (both species with monoxenous life cycles).
In the case of B. aurea (a phytoplanktivorous-type fish), the medium-sized fish feed mainly on diatoms and dinoflagellates, while the larger size prefer copepods (Cousseau et al. 2001). Therefore, the acquisition of parasites through their diet could be restricted, limiting their role in the parasites’ life cycles and being mainly a suitable host for parasites with monoxenous cycles. The component community of B. aurea from the Mar Chiquita coastal lagoon showed a parasite species richness composed by only five parasite species of which the monogenean M. georgei was the predominant species. These results agreed with previous studies on Brazilian menhaden in other similar regions in the southwestern Atlantic Ocean, i.e., from the Jacarepaguá lagoon, Río de Janeiro, Brazil (22°59′ S, 43°23′ W), where M. georgei was the species with the highest values of abundance, prevalence, and dominance (Tavares et al. 2004). Moreover, Tavares et al. (2004) suggested that the schooling behavior exhibited by this host could favor the transmission of ectoparasites and therefore might explain the dominance of monoxenous parasites like M. georgei.
The parasite fauna of P. cromis could be considered poor in comparison with the species composition of the parasite communities harbored by other sciaenids, including M. furnieri. Both host species, P. cromis and M. furnieri, share the distribution area and show a similar feeding behavior, characterized mostly by the ingestion of crustaceans. The differences in the parasite fauna of these hosts might be due to small differences in their diets and to possible behavioral changes of adult fishes in the marine environment. Unfortunately, to our knowledge, no information about this particular subject is available. Ecological studies on P. cromis are mainly restricted to feeding habits and distribution. These previous studies on the open sea indicate that P. cromis feeds on bottom organisms, particularly crustaceans, mollusks, and fish (Cousseau and Perrota 2004). However, no previous records of parasites of this host in the Argentine Sea are known. The present study represents the first contribution to the knowledge of the parasite community of this fish host.
With reference to the other sciaenid fish, M. furnieri, the trophic spectrum of young specimens in the lagoon is composed by benthic invertebrates (mainly polychaetes), isopods, gammarid amphipods, and eventually crabs (Cyrtograpsus angulatus Dana, 1851 and Neohelice granulata Dana, 1851), ostracods, mysids, and very occasionally fishes (Hozbor and García de la Rosa 2000). In the open sea, the whitemouth croaker adults feed mainly on bottom organisms (polychaetes, bivalves, snails, shrimp, other small crustaceans, etc.) and occasionally on small fish, such as anchovy (Cousseau and Perrota 2004). The diversity of food items recorded for this species suggests that it has a generalist–opportunist feeding behavior. There are previous studies on the parasite component community of M. furnieri from Mar del Plata coast and Bahía Samborombón in Argentina (Sardella et al. 1995). Comparing both study areas, there were differences between the parasite species richness of Mar Chiquita croakers and the one registered for the same host in marine zones (18 species vs 11 species). Anyway, it should be emphasized that the sample sizes and the total length of the hosts studied in both areas were different. Thus, this factor may have caused the differences in the parasites species found in each zone.
The parasite fauna of the O. argentinensis in the present study included eight metazoan parasite species. Among them, Ergasilus sp. and the digenean sp. 1 should be considered accidental parasites (only two and one specimens were found during this study, respectively). Although there are no published data regarding silver side feeding habits in Mar Chiquita coastal lagoon, the examination of intestinal contents during the present study indicated that insects, algae crabs, and cochliopid snails were the major food items for this fish host. The high percentage of parasites larval stages (Echinostome-type metacercariae, cystacanths, and plerocercoids) harbored by silver side suggests the importance of this host as consumer of the food chains first trophic link in the study area. For example, the presence of larval stages of the acanthocephalan P. chasmagnathi is a result of the ingestion of the N. granulata and C. angulatus crabs, which act as intermediate host of this parasite in the lagoon (Etchegoin 1997). In this particular case, O. argentinensis would act as paratenic host for this parasite.
Finally, P. orbignyanus feeds mainly on pelagic and benthic crustacean species and fish (Díaz de Astarloa and Munroe 1998; Cousseau and Perrota 2004). The diet of the young fishes from Mar Chiquita lagoon is based on the consumption of invertebrates, principally crustaceans amphipods, gammarids, and polychaetes (Rivera Prisco et al. 2001). The parasitological information on P. orbignyanus is restricted to taxonomical descriptions of some parasite species (Martorelli and Suriano 1983; Incorvaia and Díaz de Astarloa 1998; Braicovich and Alarcos 2007; Alarcos et al. 2008). It should be emphasized that, out of the total parasite species registered in flounder from Mar Chiquita, four specimens of nematodes belonging to Cucullanus sp., Lappetascaris sp., Anisakis sp., and Hysterothylacium sp. genus and one specimen of the acanthocephalan Corynosoma cetaceum could be considered as accidental parasites.
The results of the present study clearly indicate a close relationship between fish diet habits and parasite community composition. Although the majority of fish species examined showed a marine parasite fauna, the expression “estuarine-dependent marine fishes” cannot be ruled out from a parasitological point of view. With the exception of M. platanus, the absence of typical brackish water parasite species in most fish species studied might be due to the fact that the prey item of these fishes in the lagoon (crustaceans principally) are parasitized with larval stages utilizing birds as definitive hosts. Consequently, the digenean Microphallus szidati and acanthocephalan P. chasmagnathi showing high prevalences in N. granulata and C. angulatus crabs (Etchegoin 1997) were found in stomachal and intestinal contents of some of the fishes examined. However, these parasite species’ final hosts include the following birds: Larus atlanticus (olrog’s gull) and Himantopus mexicanus (white-backed stilt; Martorelli 1986a, b, 1989b; Etchegoin et al. 1996). These findings might also indicate that, except for the gray mullet, the presence of the rest of the fish species in the lagoon is too short for them to be included as hosts in the local parasites’ life cycles. Given the results, one might consider that the lagoon serves primarily as a temporary feeding area for the fish hosts studied. For this reason, the expression “estuarine-dependent marine fishes” remains valid, despite the minor contribution of the fish species studied to the parasites’ life cycles in the lagoon.
References
Acha EM (1990) Estudio anatómico-ecológico de la liza (Mugil liza) durante su primer año de vida. Frente Marít 7:37–43
Alarcos AJ, Etchegoin JA, Cremonte F (2008) Redescription of Bacciger microacetabularis (Martorelli et Suriano, 1983) nov. comb. parasitizing Paralichthys orbignyanus (Pisces: Paralichthyidae) from Argentina. Acta Parasitol 53:365–368
Braicovich PE, Alarcos AJ (2007) Redescription of Brasilochondria riograndensis (Copepoda: Chondracanthidae) parasitic on flounder, Paralichthys orbignyanus (Actinopterygii: Pleuronectiformes) from South American Atlantic waters. Acta Parasitol 52:273–277
Bush AO, Lafferty KD, Lotz JM, Shostak AW (1997) Parasitology meets ecology on its own terms: Margolis et al. revisited. J Parasitol 83:575–583
Chambers CA, Dick TA (2005) Trophic structure of one deep-sea benthic fish community in the eastern Canadian Arctic: application of food, parasites and multivariate analysis. Environ Bio Fisches 74:365–378
Cousseau MB, Perrota R (2004) Peces marinos de Argentina. Biología, distribución, pesca. Publicaciones especiales INIDEP, Mar del Plata
Cousseau MB, Díaz De Astarloa JM, Figueroa DE (2001) In: Iribarne O (ed) Reserva de Biosfera Mar Chiquita: Características físicas. Biológicas y Ecológicas Editorial Martín, Argentina, pp 187–207
Cremonte F, Etchegoin JA, Martorelli SR (1999a) Nuevos hospedadores de los digeneos Maritrema bonaerensis (Microphallidae) y de Stephanoprora podicipei (Echinostomatidae) en Argentina. Stephanoprora manei Holcman-Spector et Olagüe, 1986 como un nomen nudum. Neotropica 45:105–107
Cremonte F, Navone GT, Etchegoin JA (1999b) A new species of Sciadiocara Skrjabin, 1916 (Nematoda: Acuariidae) parasite of shorebirds in Argentina. Syst Parasitol 42:213–217
Díaz De Astarloa JM, Munroe TA (1998) Systematics, ecology and distribution of commercially important paralichthyd flounders occurring in Argentinean–Uruguayan waters (Paralichthys, Paralichthyidae): an overview. J Sea Res 39:1–9
Etchegoin JA (1997) Sistemas parasitarios presentes en la albufera Mar Chiquita. Doctoral thesis, Universidad Nacional de Mar del Plata, Argentina
Etchegoin JA, Martorelli SR (1997a) Description of a new species of Maritrema (Digenea: Microphallidae) from Mar Chiquita coastal lagoon (Buenos Aires, Argentina) with notes on its life cycle. J Parasitol 83:709–713
Etchegoin JA, Martorelli SR (1997b) A new species of the genus Stephanoprora Odhner, 1902 (Digenea, Echinostomatidae) from Argentina. Acta Parasitol 42:74–76
Etchegoin JA, Martorelli SR (1998) Nuevas cercarias en Heleobia conexa (Mollusca: Hydrobiidae) de la albufera Mar Chiquita. Neotropica 44:41–50
Etchegoin JA, Martorelli SR, Sardella NH (1996) Nuevo registro de Microphallus szidati Martorelli, 1986 (Digenea: Microphallidae) en Mar Chiquita (Buenos Aires, Argentina). Neotropica 42:117–118
Etchegoin JA, Cremonte F, Navone GT (2000) Synhimantus (Synhimantus) laticeps (Rudolphi, 1819) Stiles et Hassal, 1920 (Nematoda: Acuariidae) parasitic in Tyto alba (Aves: Tytonidae) in Argentina. Acta Parasitol 45:99–106
Fernández J (1987) Los parásitos de la lisa Mugil cephalus L., en Chile: Sistemática y aspectos poblacionales (Perciformes: Mugilidae). Gayana Zoología 51:3–58
García JR, Williams EH (1985) Temporal dynamics of metazoan parasite infections in the white mullet Mugil curema Valenciennes from Joyuda Lagoon, Puerto Rico. Car J Sci 21:39–53
González Castro M (2007) Los peces representantes de la familia Mugilidae en Argentina. Doctoral thesis, Universidad Nacional de Mar del Plata, Argentina
González Castro M, Abachain B, Perrota RG (2009a) Age and growth of the striped mullet, Mugil platanus (Actinopterygii, Mugilidae), in a southwestern Atlantic coastal lagoon (37°32′ S–57°19′ W): a proposal for a life-history model. J Appl Ichthyol 25:61–66
González Castro M, Astarloa DDE, JM CMB, Figueroa DE, Delpiani SM, Bruno D, Guzonni JM, Blasina GE, Deli Antoni MY (2009b) Fish composition in a south-western Atlantic temperate coastal lagoon: spatial–temporal variation and relationship with environmental variables. J Mar Biol Assoc UK 89:593–604
Hozbor NM, García De La Rosa SB (2000) Alimentación de juveniles de corvina rubia (Micropogonias furnieri) en la laguna costera Mar Chiquita (Buenos Aires, Argentina). Frente Marít 18:59–70
Incorvaia IS, Díaz De Astarloa JM (1998) Estudio preliminar de las larvas (Nematoda: Ascaridida) parásito de Paralichthys orbignyanus (Valenciennes, 1839) y Paralichthys patagonicus (Pisces: Pleuronectiformes). Bol Chil Parasitol 53:38–42
Klimpel S, Palm HW, Busch MW, Kellermanns E, Rückert S (2006) Fish parasites in the Artic deep-sea: poor diversity in pelagic fish species vs. heavy parasite load in a demersal fish. Deep Sea Res 53:1167–1181
Knoff M, Luque JL, Amato JFR (1997) Community ecology of metazoan parasites of grey mullet, Mugil platanus (Osteichthyes: Mugilidae) from the littoral of state of Río de Janeiro, Brazil. Rev Bras Biol 57:441–454
Luque JL, Poulin R (2004) Use of fish as intermediate hosts by helminth parasites: a comparative analysis. Acta Parasitol 49:353–361
Luque JL, Mouillot D, Poulin R (2004) Parasite biodiversity and its determinants in coastal marine teleost fishes of Brazil. Parasitology 128:671–682
Magurran AE (2005) Measuring biological diversity. Blackwell Science, UK
Marcogliese DJ (2002) Food webs and the transmission of parasites to marine fish. Parasitology 124:83–89
Marcogliese DJ (2003) Food webs and biodiversity: are parasites the missing link? J Parasitol 82:389–399
Martorelli SR (1986a) Estudio sistemático y biológico de un digeneo perteneciente a la familia Microphallidae Travassos, 1920. I: Microphallus szidati sp.nov. parásito intestinal de Rallus sanguinolentus sanguinolentus (Aves: Rallidae) e Himantopus melanurus (Aves: Recurvirostridae). Rev Ibér Parasitol 46:373–378
Martorelli SR (1986b) Estudio sistemático y biológico de un digeneo perteneciente a la familia Microphallidae Travassos, 1920. II: desarrollo del ciclo biológico de Microphallus szidati en dos ambientes de condiciones ecológicas diferentes. Rev Ibér Parasitol 46:379–385
Martorelli SR (1989a) Estudios parasitológicos en la albufera de Mar Chiquita, provincia de Buenos Aires, República Argentina. II: Cercarias (Digenea) parásitas de Heleobia conexa (Mollusca: Hydrobiidae), pertenecientes a las familias Schistosomatidae, Haploporidae y Homalometridae. Neotropica 35:81–90
Martorelli SR (1989b) El rol de Cyrtograpsus angulatus (Crustacea: Brachyura) en los ciclos de vida de Microphallus szidati (Digenea; Microphallidae) y Falsifilicollis chasmagnathi (Acantocephala; Filicollidae). Algunos aspectos de su ecología parasitaria. Mem Inst Oswaldo Cruz 84:567–574
Martorelli SR, Suriano DM (1983) Estudios parasitológicos en la albufera de Mar Chiquita, Provincia de Buenos Aires. República Argentina. I. Neotropica 29:195–207
Moyle PB, Cech JJJR (2004) Fishes. An introduction to ichthyology. Prentice Hall, Upper Saddle River
Navone GT, Etchegoin JA, Cremonte F (2000) Contracaecum multipapillatum (Nematoda: Anisakidae) from Egretta alba (Aves: Ardeidae), and commnets on other species of this genus in Argentina. J Parasitol 86:807–810
Olivier SR, Escofet A, Penchaszadeh P, Orensanz JM (1972) Estudios ecológicos de la región estuarial de Mar Chiquita (Buenos Aires, Argentina). I. Las comunidades bentónicas. An Soc Cient Argent 193:237–268
Poulin R (2007) Evolutionary ecology of parasites. Princeton University Press, Princeton
Reiczigel J, Rózsa L (2005) Quantitative Parasitology 3.00. Budapest
Reta RP, Martos P, Perillo GME, Piccolo MC, Ferrante A (2001) Características hidrográficas del estuario de la laguna Mar Chiquita. In: Iribarne O (ed) Reserva de Biosfera Mar Chiquita: Características físicas. Biológicas y Ecológicas Editorial Martín, Argentina, pp 31–52
Rivera Prisco A, García De La Rosa SB, Díaz De Astarloa JM (2001) Feeding ecology of flatfish juveniles (Pleuronectiformes) in Mar Chiquita coastal lagoon (Buenos Aires, Argentina). Estuaries 24:917–925
Rózsa L, Reiczigel J, Majoros G (2000) Quantifying parasites in sample hosts. J Parasitol 86:228–232
Sardella NH, Etchegoin JA, Martorelli SR (1995) Las comunidades parasitarias de Micropogonias furnieri (corvina) en Argentina. Bol Inst Oceanogr Venezuela Univ Oriente 34:41–47
Sasal P, Niquil N, Bartoli P (1999) Community structure of digenean parasites of sparid and labrid fishes of the Mediterranean Sea: a new approach. Parasitology 119:635–648
Tavares LER, Luque JL, Bicudo AJA (2004) Metazoan parasites of Brazilian menhaden Brevoortia aurea (Spix et Agassiz, 1829) (Osteichthyes: Clupeidae) from the coastal zone of the state of Rio de Janeiro, Brazil. Braz J Biol 64:553–554
Valtonen ET, Pulkkinen K, Poulin R, Julkunen M (2001) The structure of parasite component communities in brackish water of the northeastern Baltic Sea. Parasitology 122:471–481
Acknowledgments
The authors gratefully thank Laboratory of Marine Sciences from Universidad Nacional de Mar del Plata for kindly providing fish samples. We also wish to thank Dr. Robert Poulin (University of Otago, New Zealand) for his critical comments and valuable suggestions on an earlier version of the manuscript. The present study was funded by a grant from Universidad Nacional de Mar del Plata to JAE (EXA 411/08 15/E359) and by a Doctoral grant from Consejo Nacional de Investigaciones Científicas y Técnicas (CONICET) to AJA.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Alarcos, A.J., Etchegoin, J.A. Parasite assemblages of estuarine-dependent marine fishes from Mar Chiquita coastal lagoon (Buenos Aires Province, Argentina). Parasitol Res 107, 1083–1091 (2010). https://doi.org/10.1007/s00436-010-1974-z
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00436-010-1974-z