
Abstract
Actin depolymerizing factor (ADF) is an essential actin-binding protein that plays a key role in the control of actin dynamics and actin-based motility processes in intracellular parasites. To determine the effects of diclazuril on ADF gene of second-generation merozoites (mz-ADF) mRNA expression in Eimeria tenella, mz-ADF gene was cloned by RT-PCR from extracted RNA in second-generation merozoite of E. tenella and successfully expressed by pET-28a vector in Escherichia coli BL21(DE3). Results showed that the full length of the cloned cDNA sequence of the mz-ADF gene is 476 bp including an ORF of 375 bp. The sequence has 100% homology with a published sequence of sporozoite stage E. tenella ADF mRNA (GenBank EF195234.1). The recombinant protein was induced to be expressed by 1 mM isopropyl β-d-1-thiogalactopyranoside in vitro. Sodium dodecyl sulfate–polyacrylamide gel electrophoresis analysis showed that 16.99 kDa fusion protein existed in solvable form. Compared with the infected/control group, mz-ADF mRNA expression level was downregulated by 63.86% in the infected/treatment group with the treatment of diclazuril. In conclusion, the data presented here indicate that mz-ADF gene participates in an important role in the invasion host of E. tenella. Downregulation of mz-ADF mRNA expression enrich the mechanism study of diclazuril on E. tenella.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Actins dynamics, or the rapid turnover of actin filaments, play a central role in numerous cellular processes (Fuhrmanna et al. 2007; Leadsham and Gourlay 2008; Prodon et al. 2009) as well as the rocketing motion of intracellular parasites. A large and diverse cast of characters, accessory proteins known as actin-binding proteins, modulate actins dynamics (Staiger and Blanchoin 2006; Michelot et al. 2007). Apicomplexan genomes contain relatively few conventional actin-binding proteins including ADF (Santos et al. 2009) and which controls actin dynamics and actin-based motility processes.
E. tenella, an obligate intracellular apicomplexan (coccidian) parasite, can efficiently and rapidly invade chicken cecum epithelial cell to sustain living and expand colonia, which is an important causal agent of clinical coccidiosis in chickens. E. tenella lifecycle is complex involving endogenous developmental stage (schizogamy and gametogony) and exogenous developmental stage (sporogony). It is in the schizogamy stage that a large number of second-generation merozoites released by host cell lysis do not grow or undergo cell division extracellularly and must rapidly attach to and actively reinvade other host cells (Russell 1983) and eventually lead the most damage to the intestinal tissues. Xu et al. (2008) reported that ADF was expressed in all life stages of E. tenella, including merozoites, sporozoites, unsporulated oocysts, and sporulated oocysts by southern blot analysis, and ADF protein existed in higher amounts in merozoites and sporozoites than in sporulated and unsporulated oocysts by dot-blotting analysis. However, character of mz-ADF sequence has not been completely defined. Cloning and sequence of mz-ADF gene of E. tenella laid an important foundation for further functional study.
Diclazuril, a benzeneacetonitrile anticoccidial, has potent activity against the asexual and sexual stages of E. tenella development (Xie et al. 1991; Maes et al. 1988; Verheyen et al. 1988; Nodeh et al. 2008). However, the exact mechanisms that drive inhibition of diclazuril in second-generation merozoite remain incompletely understood. In the present study, cloning and expression of mz-ADF gene of E. tenella was analyzed in vitro. To further elucidate potential mechanisms contributing to dicluzuril on E. tenella, mz-ADF mRNA expression were also determined.
Materials and methods
Preparation of inoculum
Oocysts of E. tenella were kindly provided by Shanghai Veterinary Research Institute, Chinese Academy of Agricultural Sciences (CAAS). Oocysts were propagated, isolated, sporulated, and counted prior to inoculation of subjects. The total number of sporulated oocysts was estimated by multiplication of the obtained number by the dilution factor.
Experimental drug
Diclazuril (>99%, Shanghai Veterinary Research Institute, CAAS. Product No: 20080812) was given through the feed at concentration of 1 mg/kg.
Chickens and treatment
Two hundred forty 14-day-old Chinese Yellow Broiler male chickens, from Hatchery Huizhong with a good reputation of producing diseased-free chickens, were randomly allotted to two groups of 120. Each group was maintained as the following: (1) Chickens were challenged with E. tenella oocysts and were administrated with normal feed, infected/control group; (2) Chickens were challenged with E. tenella oocysts and administrated with 1 mg/kg diclazuril in feed at 96 h after inoculation, for 24 h, infected/treatment group. Chickens were inoculated by oral gavage with an 8 × 104 oocysts/chick suspended in 1 ml of distilled water. The experimental design was approved by the local committee of the Faculty of Veterinary Medicine and conformed to the guidelines of Institutional Animal Care and Use Committee of China.
Preparation of the second-generation merozoite
With the combination application of enzymatic digestion, centrifugation, erythrocytes disruption, and Percoll density gradient centrifugation, the second-generation merozoites of E. tenalla were obtained from parasitic ceca by the technique modified based on what has been described previously (Liu et al. 2006) for further study.
Total RNA extraction and purification and cDNA synthesis
Total RNA was extracted with TRIzol® Reagent (Invitrogen, USA) according to the manufacturer’s instructions. To avoid DNA contamination, the extracted RNA preparations were additionally treated with RNase-free DNase I (40 U/mg RNA Takara, China) for 30 min at 37°C. DNase I was inactivated afterwards by heating (75°C, 10 min). The total RNA was purified by the RNeasy Mini Kit (Qiagen, Germany) according to the manual. cDNA was synthesized by second-generation merozoite purified total RNA using SuperScript™ II Reverse Transcriptase kit (Invitrogen, USA) and pd (N)6 random hexamer primers. The cDNA was then used as template for further study.
Cloning of E. tenella mz-ADF gene
According to the sequences obtained from GenBank: EF195234.1, two specific primers P1 and P2 (Table 1) were used through using a PC-818A Progaram Temp Control System (Astec, Japan) under the following condition: 30 cycles of 94°C for 30 s, 55°C for 30 s, and 72°C for 1 min, followed by 10 min at 72°C. The PCR product was purified by electrophoresis in a 1% agarose gel and extracted using gel extraction kit (Qiagen, Germany) according to manufacturer’s instructions. The purified PCR amplified fragments were ligated into the pGEM®-T-Easy cloning vector (Promega, USA) following the manufacturer’s recommendations, and the ligation mixture was used to transform Escherichia coli strain DH5α. TIANprep Mini Plasmid Kit (Tiangen, China) preparations of the recombinant plasmid (pGEM-mz-ADF) were analyzed by EcoRI enzyme digestion and electrophoresis. Positive recombinant clone was sequenced using an ABI 3730 automated sequencer by Shanghai Sunny Biotechnology Co. Ltd., and sequence data were assembled and analyzed by DNAstar software.
Expression of His6-mz-ADF fusion protein
Based on the positive recombinant plasmid (pGEM-mz-ADF) as template, cDNA fragments corresponding to the ORF of E. tenella mz-ADF were amplified by PCR using specific primers P3 and P4 (Table 1) with HindIII and EcoRI sites at the 5′ and 3′ ends of the fragment, respectively. The cloned products were digested with HindIII and EcoRI. The purified PCR amplified fragments using gel extraction kit (Qiagen, Germany) were then ligated overnight into pET-28a vector digested by the same restriction enzyme HindIII and EcoRI with T4 DNA ligase (Promega, USA) at 16°C, which was transformed into E. coli strain BL21 (DE3). The empty plasmid was also transformed into E. coli strain BL21 (DE3) as a negative control. The expression was induced by isopropyl β-d-1-thiogalactopyranoside (IPTG). Recombinant-expressed E. tenella mz-ADF with His6-tag were enriched and purified from the lysated supernatant using the His·Bind Purification® Kit (Novagen, Germany) and determined by sodium dodecyl sulfate–polyacrylamide gel electrophoresis (SDS-PAGE).
Real-time PCR analysis of mz-ADF gene expression
The expression level of mz-ADF gene was quantified by real-time amplification of this purpose gene and the housekeeping gene 18S rRNA as control from the above cDNA preparation using the RG-3000A real-time PCR system (RoterGene, USA) and SYBR® Premix Ex Taq™ (Perfect Real Time) kit (Takara, China). The real-time PCR reaction mixture (20 μl) contained 10 μl SYBR® Premix Ex Taq™ (2×), 0.2 μM of above-described primer, 1 μl cDNA template, and 7 μl RNase-free distilled H2O. The real-time PCR protocol included an initial denaturation at 95°C for 15 s. This was followed by 40 PCR cycles consisting of a denaturation step at 95°C for 5 s, an annealing step at 55°C for 10 s, an extension step at 72°C for 15 s. The PCR reaction was then subjected to a melting protocol. The specificity of amplification was confirmed by agarose gels with ethidium bromide and direct sequencing of the PCR products. The sequences of the primers are reported in Table 2.
Statistical analysis
Data are expressed as mean ± SD. Statistical analyses were performed by Student’s t test. Values of p < 0.05 were considered significant.
Results
Amplification of the ADF gene
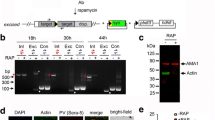
The 476-bp mz-ADF gene amplification products were generated from the cDNA of second-generation merozoite (Fig. 1). PCR products purified by gel extraction kit was cloned into pGEM-T-Easy vector; recombinant plasmid were analyzed by electrophoresis (Fig. 2) and EcoRI enzyme digestion (Fig. 3). Positive recombinant clones were sequenced by Shanghai Sunny Biotechnology Co. Ltd., and sequence data (Fig. 4) have 100% homology with a published sequence of sporozoite stage E. tenella ADF mRNA (GenBank: EF195234.1).

Agarose gel electrophoresis of mz-ADF RT-PCR products of 476 bp fragment of E. tenella. M DL2000 DAN marker; 1 and 2, mz-ADF RT-PCR products; 3 negative control

Identification of recombinant plasmid (pGEM-mz-ADF) containing 476 bp fragment by PCR amplification. M DL2000 DNA marker; 1 and 2 amplification by specified primer; 3 negative control

Identification of recombinant plasmid (pGEM-mz-ADF) containing 476 bp fragment by digestion with endonuclease EcoR I. M1 DL2000 DNA marker; 1–4 pGEM-mz-ADF digested by EcoRI. M2 DL12000 DNA marker

Sequence of mz-ADF fragment of E. tenella. Sporozoite (Spo); second-generation merozoite (SGM)
Recombinant protein expression of mz-ADF
A DNA fragment encoding 117 amino acids of mz-ADF gene, which included the amino initiation and terminus of the mature protein, was amplified from positive recombinant plasmid pGEM-mz-ADF by PCR and subcloned into the expression vector pET-28a to give rise to the construct pET-28a-mz-ADF. Recombinant expression plasmid pET-28a-mz-ADF was analyzed by electrophoresis (Fig. 5) and EcoRI/HindIII enzyme digestion (Fig. 6). Selected recombinant clones were sequenced by Shanghai Sunny Biotechnology Co. Ltd. Expression of the recombinant proteins in E. coli BL21 (DE3) was induced using 1 mM IPTG at 37°C, for 4 h, to get to high expression. The expressed and purified protein was high to visualize by Coomassie Blue staining of whole bacterial lysates separated by SDS-PAGE (Fig. 7) and had a theoretical molecular weight of 16.99 kDa solvable fusion protein, including the mass contributed by the fusion.

Agarose gel electrophoresis of mz-ADF gene (ORF) PCR products. M DL2000 DNA marker; 1–4 PCR products

Agarose gel electrophoresis of EcoRI/HindIII restriction enzyme assay of recombinant expression plasmid. M2 DL 12000 DNA marker; 1 and 2 restriction enzyme assay of recombinant plasmid pET-28a-mz-ADF; M1 DL2000 DNA marker

Expression of ADF-mz in E. coli BL21 (DE3). M Protein marker; 1 negative control (not induced); 2–7 induced at different time (1, 2, 4, 6, 8, and 10 h, respectively); Su supernatant; Pe pellet; Pu purified product assay of recombinant plasmid pET-28a-mz-ADF
Expression of mz-ADF mRNA
In order to determine the effect of diclazuril on mz-ADF, the relative expression level of mz-ADF mRNA was measured using real-time PCR. The standard curves of 18S rRNA and mz-ADF gene were y = −3.174 × log(x) + 31.172 and y = −3.349 × log(x) + 41.118, respectively (Fig. 8), which were obtained by correlation of the Ct values with the dilution series. The amplification efficiencies of the 18S rRNA and mz-ADF were 107% and 99%, respectively. The mz-ADF mRNA expression level was downregulated by 63.86% with diclazuril administration (Fig. 9). Melting curves profile (Fig. 8) showed high amplification specificity.

Standard and melting curves of 18S rRNA and mz-ADF

The relative expression level of ADF
Discussion
Protozoan parasites of the genus Eimeria cause great economic losses in the poultry industry. E. tenella, an important protozoan parasite, is one of the most virulent of seven species of Eimeria that infects chickens, develops in the intestinal cecum, and provokes hemorrhage and, in severe cases, anemia and death due to blood loss and shock. In the whole lifecycle, E. tenella compulsory invasive to susceptible host cell depending on a system of adhesion-based motility called “gliding” to actively penetrate host cells which are characterized by a unique complex of specialized structures and driving force from motility of actin filaments. ADF can enhance the turnover and treadmilling of actin filaments (Didry et al. 1998; Carlier et al. 1997). Actin polymerization-dependent motility facilitates parasite migration across cellular barriers, enables dissemination within tissues, and powers invasion of host cells (Sibley 2004). Polymerization at barbed ends of actin filaments has been proposed to provide the driving force for forward movement, while dissociation of actin subunits from free pointed ends by ADF allows for filament actin turnover and not to completely depolymerize actin (Ono 2007). As depolymerization is a rate-limiting step in actin dynamics, ADF is suspected to increase microfilament dynamics by increasing the rate at which actin monomers leave the pointed end of the filament and by a filament-severing activity (Maciver and Hussey 2002). The ADF has been shown to play an important role in the rapid depolymerization of actin filaments via a pH-dependent during actin-based motility, such as cytokinesis (Abe et al. 1996), polarization (Chen et al. 2002; Bernstein and Bamburg 2004), and tip growth in plants (Augustine1 et al. 2008). In the field of protozoon, Tammana et al. (2008) reported that when the ADF/cofilin gene of Leishmania flagellum was knocked out by targeted gene replacement, the resultant mutants were completely immotile.
In the present study, an E. tenella mz-ADF cDNA was cloned and sequenced, and the sequence has 100% homology with a published sequence of sporozoite stage E. tenella ADF mRNA (GenBank EF195234.1). High-sequence homology and different patterns of temporal and spatial expression of ADF endow it an important functional member during the whole life of E. tenella. The recombinant protein was induced to be expressed and purified as fusion protein, which were shown by SDS-PAGE (Fig. 7). Since the pET-28a having six His-label was selected as the expression vector in this experiment, these expressed recombinant protein could be purified by His Bind Resin after nondenaturalization, and the function of mz-ADF can be further researched in the next step.
Diclazuril is a benzeneacetonitrile belonging to the group of nucleoside analogs. Studies have demonstrated that this compound has a broad spectrum activity against all pathogenic Eimeria species in chickens. In E. tenella, detailed histological, clinical, and ultrastructural studies have shown that diclazuril, given in a single oral dose at different periods after inoculation, was fully effective against asexual as well as sexual stages (Maes et al. 1988; Verheyen et al. 1988). It was found that treatment with diclazuril downregulated mz-ADF mRNA expression by 63.86% in the present study. Indications from this study were that diclazuril downregulated the mz-ADF mRNA expression, which in turn, interferes filament turnover and inhibit second-generation merozoite invasion to host. mz-ADF is prone to be a pathway that diclazuril antagonize coccidiosis, which has not been reported so far.
In conclusion, these results indicate that mz-ADF gene participate in an important role in the invasion host of E. tenella. Downregulation of mz-ADF mRNA expression enrich the mechanism study of diclazuril on E. tenella. Further, mz-ADF, as an attractive target for control coccidiosis, remains to be investigated.
References
Abe H, Obinata T, Minamide LS, Bamburg JR (1996) Xenopus laevis actin-depolymerizing factor/cofilin: a phosphorylation-regulated protein essential for development. J Cell Biol 132:871–885
Augustine RC, Vidali L, Kleinman KP, Bezanilla M (2008) Actin depolymerizing factor is essential for viability in plants, and its phosphoregulation is important for tip growth. Plant J 54:863–875
Bernstein BW, Bamburg JR (2004) A proposed mechanism for cell polarization with no external cues. Cell Motil Cytoskeleton 58:96–103
Carlier MF, Laurent V, Santolini J, Melki R, Didry D, Xia GX, Hong Y, Chua NH, Pantaloni D (1997) Actin depolymerizing factor (ADF/Cofilin) enhances the rate of filament turnover: Implication in actin-based motility. J Cell Biol 136:1307–1322
Chen CY, Wong EI, Vidali L, Estavillo A, Hepler PK, Wu HM, Cheung AY (2002) The regulation of actin organization by actin-depolymerizing factor in elongating pollen tubes. Plant cell 14:2175–2190
Didry D, Carlier MF, Pantaloni D (1998) Synergy between actin depolymerizing factor/cofilin and profilin in increasing actin filament turnover. J Biol Chem 273:25602–25611
Fuhrmanna J, Käs J, Stevens A (2007) Initiation of cytoskeletal asymmetry for cell polarization and movement. J Theor Biol 249:278–288
Leadsham JE, Gourlay CW (2008) Cytoskeletal induced apoptosis in yeast. Biochim Biophys Acta 1783:1406–1412
Liu LH, Li J, Xu LX, Li XR, He TW (2006) Separation and purification of second-generation merozoites of Eimeria tenella. Animal Husbandry & Veterinary Medicine 38:38–40
Maciver SK, Hussey PJ (2002) The ADF/cofilin family: actin-remodeling proteins. Genome Biol 3:3007.1–3007.12
Maes L, Coussement W, Vanparijs O, Marsboom R (1988) In vivo action of the anticoccidial diclazuril (Clinacox®) on the developmental stages of Eimeria tenella: a histological study. J Parasitol 74:931–938
Michelot A, Berro J, Guérin C, Boujemaa-Paterski R, Staiger CJ, Martiel JL, Blanchoin L (2007) Actin-filament stochastic dynamics mediated by ADF/cofilin. Curr Biol 17:825–833
Nodeh H, Mansoori B, Rahbari S, Modirsanei M, Aparnak P (2008) Assessing the effect of diclazuril on the intestinal absorptive capacity of broilers infected with experimental coccidiosis, using D-xylose absorption test. J Vet Pharmacol Ther 31:265–267
Ono S (2007) Mechanism of depolymerization and severing of actin filaments and its significance in cytoskeletal dynamics. Int Rev Cytol 258:1–82
Prodon F, Hanawa K, Nishida H (2009) Actin microfilaments guide the polarized transport of nuclear pore complexes and the cytoplasmic dispersal of Vasa Mrna during GVBD in the ascidian Halocynthia roretzi. Dev Biol 330:377–388
Russell DG (1983) Host cell invasion by Apicomplexa: an expression of the parasite’s contractile system? Parasitology 87:199–209
Santos JM, Lebrun M, Daher W, Soldati D, Dubremetz JF (2009) Apicomplexan cytoskeleton and motors: key regulators in morphogenesis, cell division, transport and motility. Int J Parasitol 39:153–162
Sibley LD (2004) Intracellular parasite invasion strategies. Science 304:248–253
Staiger CJ, Blanchoin L (2006) Actin dynamics: old friends with new stories. Curr Opin Plant Biol 9:554–562
Tammana TV, Sahasrabuddhe AA, Mitra K, Bajpai VK, Gupta CM (2008) Actin-depolymerizing factor, ADF/cofilin, is essentially required in assembly of Leishmania flagellum. Mol Microbiol 70:837–852
Verheyen A, Maes L, Coussement W, Vanparijs O, Lauwers F, Vlaminckx E, Borgers M, Marsboom R (1988) In vivo action of the anticoccidial diclazuril (Clinacox®) on the developmental stages of Eimeria tenella: an ultrastructural evaluation. J Parasitol 74:939–949
Xie MQ, Fukata T, Gilbert JM, McDougald LR (1991) Evaluation of anticoccidial drugs in chicken embryos. Parasitol Res 77:595–599
Xu JH, Qin ZH, Liao YS, Xie MQ, Li AX, Cai JP (2008) Characterization and expression of an actin-depolymerizing factor from Eimeria tenella. Parasitol Res 103:263–270
Acknowledgements
This study was financially supported by central grade public welfare fundamental science fund for scientific research institute (contract grant number: 2008JB09).
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Zhou, Bh., Wang, Hw., Xue, Fq. et al. Actin-depolymerizing factor of second-generation merozoite in Eimeria tenella: clone, prokaryotic expression, and diclazuril-induced mRNA expression. Parasitol Res 106, 571–576 (2010). https://doi.org/10.1007/s00436-009-1699-z
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00436-009-1699-z