
Abstract
A new multivalvulid myxozoan parasite, Kudoa unicapsula n. sp., is described from the intestinal mesentery, intestine and pyloric caeca of the thin-lipped grey mullet Liza ramada (Risso 1826) and the golden grey mullet L. aurata (Risso, 1810) from the Mediterranean coastal waters of Spain. It is characterized by the presence of elongated, rice corn-like white cysts of 0.47–0.56 × 0.18–0.38 mm, filled with tetracapsulate, slightly asymmetric spores, rectangular in apical view and tear-shaped in lateral view with four polar capsules of considerably different size and slightly unequal spore valves with rounded edges, overlapping each other on the apex of the spore. One large polar capsule includes a polar filament coiled in two to three turns, and the other three polar capsules, which are very small, posses only a rudimental filament. Both light and electron microscopy data showed that this species differs from all previously described Kudoa spp. with unequal polar capsules. The molecular analysis based on 18S and 28S ribosomal deoxyribonucleic acid DNA sequence data of K. unicapsula n. sp. indicates a close relationship and thus phylogenetic clustering together with K. trifolia, a myxozoan from the same host and the same geographical location.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Currently, 84 species, including 25 unnamed, belong to the genus Kudoa Meglitch 1947. Most species of the genus are histozoic parasites of more than 180 species of marine and estuarine fishes. Thirty-seven identified and 19 unnamed species infect the muscle, while the others occur in different internal organs. Five species have been found in the mesentery associated with the intestine and in the pyloric caeca of fish. Some representatives of the genus Kudoa cause important economical losses because of the reduction in the market value of products from infected fish (e.g. K. azoni Aseeva 2004, K. megacapsula Yokoyama and Itoh 2005, K. thyrsites (Gilchrist 1924), K. histolytica (Perard 1928), K. musculoliquefaciens (Matsumoto 1954), K. amamiensis Egusa and Nakajima 1980, K. miniauriculata Whitaker, Kent and Sakanari 1996, K. rosenbuschi (Gelormini 1943); Whitaker et al. 1994; Aseeva 2004; Moran et al. 1999; Naidenova Gaevskaya 1991) but also because of superficial ulceration of the fish skin (e.g. K. clupeidae (Hahn 1917), K. mirabilis Naidenova and Gaevskaya 1991).
Myxozoans with apparently unicapsulate spores were found during a parasitological study of mullets from the estuary of the River Ebro and from littoral parts off the Spanish Mediterranean coast, in May 2004 and June 2005. The ultrastructural analysis exposed three reduced polar capsules, and together with molecular data, the species presented itself as a member of the genus Kudoa. In the present study, the spore morphology, ultrastructural features and molecular identity of this new myxozoan species are presented.
Materials and methods
Mullets were purchased at local fish markets from three localities in the Spanish Mediterranean during May/June 2004 and 2005 namely, from Santa Pola Bay, Santa Pola Salt Marshes and the River Ebro Delta. A total of 350 specimens, belonging to five species of Mediterranean mullets were examined: flathead mullet Mugil cephalus Linnaeus 1758 (179 specimens), golden grey mullet Liza aurata (Risso 1810) (121 specimens), thin-lip mullet L. ramada (Risso 1826) (29 specimens), leaping mullet L. saliens (Risso 1810) (ten specimens) and thick-lip grey mullet Chelon labrosus (Risso 1827) (ten specimens). Fish were transported to the laboratory on ice and dissected on arrival according to a standardised protocol specifically developed for mullets (Kostadinova et al. 2004). Morphological and molecular data of the myxozoan described in this study were obtained from fresh fish, whereas fish used for prevalence data were frozen at −20°C and thawed on the day of examination. Skin, gills, brain, digestive tract, gall and swim bladder, liver, kidney, heart, spleen, muscle and eyes were examined for myxosporean infections. Fresh preparations of spores on slides were embedded in gelatin–glycerin (1:1). Spores were photographed and measured on digital images. Descriptions of the spores were based on the guidelines of Lom and Arthur (1989), Lom and Dykova (1992) and Schulman et al. (1997). According of Lom and Dykova (1992), the measurements of Kudoa spp. include width, length and thickness of the spore. Some additional measurements of spore width and length were taken because of existing asymmetry (see Fig. 1 for definition of new spore measurements).

Diagram of mature spore of K. unicapsula n. sp. in apical (a) and lateral (b) view and method of measurement.W1, W2 width of the spore, T1, T2 thickness of the spore, L length of the spore
For ultrastructural analyses, infected tissues were fixed in a 2.5% (v/v) glutaraldehyde in 0.1 M sodium cacodylate buffer (pH 7.4) for several days at 4°C. After washing twice with 0.1 M sodium cacodylate buffer and post-fixation in 2.0% (v/v) osmium tetroxide in cacodylate buffer for 1 h at 4°C, the pieces were dehydrated and embedded in Epon–Araldite solution using a standard procedure (Vavra and Maddox 1976). Blocks of embedded tissues were sectioned with an LKB III ultra-microtome. Semi-thin sections were stained with methylene blue. Ultra-thin sections were mounted on copper grids, double stained with uranyl acetate and lead citrate and examined in a JEM 100B electron microscope operated at 80 kV.
For molecular analysis, deoxyribonucleic acid (DNA) was extracted from three individual rice corn-sized plasmodia from three individuals of L. ramada. Isolated plasmodia were transferred to TNES–urea, and DNA was extracted using a phenol–chloroform protocol as previously described (Holzer et al. 2004). 18S ribosomal DNA (rDNA) was amplified using universal eukaryotic 18S primers (ERIB1: 5′-ACCTGGTTGATCCTGCCAG-3′ and ERIB10: 5′-CTTCCGCAGTTCACCTACGG-3′, Barta et al. 1997). Sequencing reactions were carried out as in Holzer et al. 2006. The polymerase chain reaction (PCR) products obtained were purified for sequencing using the High-Pure PCR Product Purification Kit (Roche Diagnostics, Germany). Primers ERIB1 and ERIB10 (see above) as well as LIN3 (5′-GCGGTAATTCCAGCTCCA-3′, Lin et al. 1999) and LIN10 (5′-CACTCCACGAACTAAGAA-3′, Lin et al. 1999) were used for cycle sequencing of the 18S fragments in a 48 capillary ABI 3730 sequencer (Applied Biosystems) using the BIG Dye Terminator v3.1 Ready Sequencing Kit (Applied Biosystems). Because of the close molecular similarity of the 18S rDNA sequences of the present myxozoan with that of K. trifolia, additional 28S rDNA sequences were obtained for both species, using primers Kt28S1F (5′-CAAGACTACCTGCTGAAC-3′, Whipps et al. 2004) and 28S1R (5′-GTGTTTCAAGACGGGTCG-3′, Whipps et al. 2004). Sequences of K. trifolia were obtained from two samples used for sequencing of the18S rDNA (Holzer et al. 2006). Sequence alignment with other Kudoa spp. was conducted using Clustal X (Thompson et al. 1997). Tree inference was carried out by a likelihood-based Bayesian tree sampling procedure using MrBayes v 3.0 (Ronquist and Huelsenbeck 2003) with parameters corresponding to the general time-reversible model GTR + I + Г (Tamura and Nei 1993). The MCMC was allowed to run for 1,000,000 generations sampling every 100th tree. The first 24,000 generations were later discarded as the burn-in period.
Results
The following are the characteristics of Kudoa unicapsula sp. n.:
-
Type hosts: Thin-lip mullet Liza ramada (Risso 1826) and golden grey mullet L. aurata (Risso, 1810)
-
Prevalence: 40% (10/25) in L. ramada and 15% (18/121) in L. aurata
-
Type localities: Mediterranean; Santa Pola Bay (38°11′N, 0°34′W) and Ebro Delta (40°42′N, 0°43′W), Spain
-
Site of infection: Intestinal mesentery, intestine, and pyloric caeca
-
Pathogenicity: Parasite plasmodia were sometimes encapsulated by fibrous connective tissue (Fig. 2b,c) but no other host tissue reactions were observed. Infection caused no clinical signs in any of the fish species
-
Etymology: Species name refers to the presence of only one large, easily visible polar capsule and three reduced polar capsules, which are difficult to be discriminated by light microscopy
-
Material deposited: Syntypes of gelatin–glycerin-embedded spores on slides 1 MRE 1, 11; 2 MRE 11, 23; 2 MRS 35, 45, 48, 50, 59; 3 MRS 6; 2 MAE 38 were deposited in Department of Ecological Parasitology, Institute of Biology of the Southern Seas, Sevastopol, Ukraine, and in the Microbiology Slide Collection of the Natural History Museum, London, UK
-
Light microscopy data (Figs. 1, 2a,b,d; Table 1): Plasmodia polysporous, elongated, rice corn-like in shape, 0.47–0.56 × 0.18–0.38 mm (n = 20) in size (Fig. 2a). Spores asymmetrical, rectangular in apical view and drop-shaped in lateral view with slightly flattened apical and rounded ventral pole. Polar capsules ovoid, distinctly different in size (Fig. 2d): One large polar capsule with polar filament in two to three coils and three degenerate polar capsules, inconspicuous by light microscopical observation. Four slightly unequal shell valves enclosing the spore contents. Valve containing large capsule slightly bigger then the others and rounded. Other three shell valves smaller, with straighter edges. Spore measurements are presented in Table 1.
-
Transmission electron microscopy (Figs. 2c,e, 3): Parasite plasmodium encapsulated by collagenous border of 0.04 mm width containing numerous, sometimes interconnected microfibres (Fig. 2c). Shell valves of spore 70 nm thick, adhering along sutural lines (Fig. 3a). Valve edges round. Thickened valve flanges of 150 nm in width overlapping on apical pole of the spore (Fig. 3d,f). Overlapping parts of shell valves contain regular patches of electron-dense material (Fig. 3d). Only one large polar capsule fully developed in mature spore (Fig. 3a,d,g). Remaining three capsules degenerated with weakly developed polar filaments (Fig. 3a,b,e). Large polar capsule with 230-nm-wide wall consisting of two layers, i.e. an internal, electron-lucent layer, 170 nm wide and an external, finely granular electron-dense cover (width?). Both layers continue into polar filament wall (Fig. 3d,g). Polar capsule opening filled with finely granular material of the same structure as outer layer of capsule wall (Fig. 3d). Capsular primordium of immature spore composed of finely granular matrix core and coarse, electron-dense cortex of future polar filament (Fig. 3f). Polar filament appears as a flattened, S-like tube in cross-sections, 200 nm thick and 310 nm wide, with an outer, electron-transparent layer, 70 nm wide and an inner, electron-dense layer, 50 nm wide. Discharged polar filament electron dense (Fig. 2e). Degenerate polar capsules with low number of electron-dense units (Fig. 3c). Sporoplasm consists of two uninucleate cells, a primary cell enveloping a secondary cell (Fig. 3c). Membrane-bound vacuole measures 700 nm in diameter and harbours one of sporoplasm nuclei (Fig. 3f).
-
Molecular data (Fig. 4): 18S rDNA sequencing of K. unicapsula n. sp. produced a 1,505-bp sequence (GenBank accession number AM490334). Alignment with 18S rDNA sequences of Kudoa spp. identified K. unicapsula n. sp. as a member of the genus and showed close sequence similarity between K. unicapsula n. sp. and K. trifolia (99.5%), a myxozoan from the same geographical area and the same mugilid hosts. The comparison of 28S rDNA sequences of K. unicapsula n. sp. (728 bp, AM490335) and K. trifolia (795 bp, AM490336) showed only slightly lower (89.8%) sequence identity between the two species. As a result of the high sequence similarity, the two species clustered together with 1.00 clade posterior probability in both the 18S (not shown) and the 28S (Fig. 4) rDNA phylogenetic analysis of Kudoa. This signifies that all the trees sampled during the analysis contained the K. unicapsula n. sp./K. trifolia branch.
-
Taxonomic remarks: The myxozoan parasite described can be placed in Kudoa Meglitsch, 1947 based on the shape and structure of the spore but also because of its molecular similarity to other representatives of the genus. To date, nine species of Kudoa have been reported to parasitize mullets. Five of them, K. cascasia, K. chilkaensis, K. ciliatae, K. dianae and K. iwatai, were recorded from the mesentery surrounding the intestine and from the pyloric caeca of mullets from commercial fish of the Indian and the Pacific Oceans (Sarkar and Chaudhury 1996; Tripathi 1951; Lom et al. 1992; Dykova et al. 2002; Egusa and Shiomitsu, 1983). The spore morphology of K. unicapsula n. sp. is distinct from all these species because of the presence of one large and three very small, degenerate polar capsules. Unequal polar capsules have been described from eight Kudoa species (Table 2). Of these, K. unicapsula n. sp most closely resembles Kudoa sp. Bunton and Poynton 1991 infecting Morone americana (Gmelin) of North American coastal waters (Bunton and Poynton 1991). Both species were recorded in euryhaline waters but differ by hosts and tissue location. However, differences in shape and/or morphometry exist between K. unicapsula n. sp. and all species summarized in Table 2, so that the unique morphology and molecular characteristics classify it as a new species.

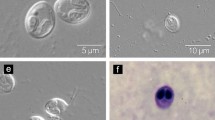
K. unicapsula n. sp. light and ultrastructural images of plasmodial (a, b, c) and spore (d, e) morphology. a Elongated rice corn-like plasmodia (arrow) form the pyloric caeca of Liza aurata. b Semi-thin section of spore forming plasmodium stained with methylene blue. The plasmodium is surrounded by host connective tissue (arrow). c. Ultra-thin section of part of the same plasmodium as in b. Collagen microfibres (arrowed) and mature spores (S) are visible. d Interference-contrast image of fresh spores of K. unicapsula n. sp. e Ultrathin section of the spore with polar filament (PF) extruded from large polar capsule (PC). Scale bars: a = 0.2 mm, b = 0.1 mm, c = 2.0 μm, d = 4.8 μm, e = 1.0 μm

Ultrastructural features of mature spores of K. unicapsula n. sp. a Transverse section through basal part of the immature spore showing two polar capsules (PC), mitochondrion (M), valvogenic cells (VC) and suture lines (arrows). Big polar capsule contains two coils of polar filament (PF). b Cross-section of mature spore showing four unequally sized polar capsules (PC). c Spore with two unequal nuclei (Nu) and small polar capsule (PC). d Longitudinally sectioned large polar capsule with polar filament arranged in longitudinal coils. The polar capsular opening (PP) is filled by finely granular material of the same structure as outer layer of capsule wall. The polar capsule entrance is covered by the shell valve flanges containing patches of electron dense material. e Longitudinal section of small polar capsule containing weakly developed polar filament (PF). f Diagonally sectioned immature spore. Four shell valves are overlap at the spore apex (arrowed). Valvogenic cell (VC) and capsulogenic cell with developing polar filament (PF) and nucleus (Nu) are shown. Membrane-bounded vacuole (Va) is filled by amorphous substance. g Longitudinal section of spore. Large polar capsule (PC) contains fully developed polar filament. End of polar filament (PP) is located near junction of shell valves. Scale bars: a = 1.1 mm, b = 1.0 mm; c = 1.2 mm, d = 0.8 μm, e = 0.5 μm, f = 1.1 μm, g = 0.8 μm

Phylogenetic tree based on Bayesian inference (GTR + I + Г) of 28S rDNA data of Kudoa spp. showing position of K. unicapsula n. sp. Henneguya salminicola and Myxobolus cerebralis are outgroup species. The numbers at the nodes show the clade posterior probability, which is the proportion of the sampled trees containing that branch
Discussion
Out of five examined mullet species from the Spanish Mediterranean, only two, L. ramada and L. aurata, were infected by K. unicapsula n. sp. indicating certain host specificity. K. unicapsula n. sp. was found in wild population of grey mullets from the Santa Pola Bay and the Ebro Delta but not in the Santa Pola Salt Marshes. Today, these marshes are not used for salt production any more but are filled with freshwater from the adjacent river and used for freshwater aquaculture. This might be indicative for the presence of the alternate host in marine and brackish water but not in freshwater. This is in accordance with the fact that Kudoa is a generally marine genus, which might be dependent on generally marine alternate host species.
Despite the three reduced polar capsules, K. unicapsula n. sp. shows a morphology that is typical for Kudoa, with four polar capsules and subquadrate spores. However, its closest relative on a molecular basis (18S and 28S rDNA), i.e. K. trifolia, shows a considerably different morphology because of the extensive enlargement of a valve cell. At the same time, these two species show a large amount of molecular identity in two ribosomal gene regions, with only 0.5 (18S) and 10.2% (28S) sequence divergence. A similar amount of divergence (0.2–1%) can be found between the 18S rDNA sequences of different geographic isolates of K. thyrsites; however, K. thyrsites intraspecific divergence is lower (3–9.6%) than the one between K. unicapsula n.sp. and K. trifolia (10.2%) when the 28S rDNA region is compared (Whipps and Kent 2006). The sequence data of K. thyrsites also shows that 28S rDNA divergence between geographical isolates from the same oceanic region (Eastern Pacific, Atlantic, Australia and Japan) is below 1% for 28S, but K. unicapsula n. sp. and K. trifolia occur in a very small enclosed habitat, i.e. the Mediterranean with much higher sequence divergence. Interspecific variance between the members of Kudoa is 1–28% for 18S rDNA and 9.4–33% for 28S rDNA. This shows that on the basis of the 18S rDNA, K. unicapsula n. sp. and K. trifolia could be conspecific but not when 28S rDNA sequences are compared. However, it has been indicated several times that the 18S rDNA sequence is of limited use in clarifying the systematics of closely related myxozoans (e.g. Kallert et al. 2006; Fiala 2006). Another feature, i.e. site preference, strongly indicates that K. unicapsula n. sp. and K. trifolia are not conspecific: K. unicapsula n. sp. occurs exclusively in the pyloric caeca, intestine and intestinal mesentery, whereas K. trifolia can be found in various sites in and along the digestive tract and in other internal organs but in contrast to K. unicapsula n. sp. is always closely associated with connective tissue (Holzer et al. 2006). Site of infection assists in the identification of myxozoan species, as tissue tropism and cyst location in some cases have been shown to be more important than morphology, geographical distribution or host specificity (Eszterbauer 2004; Fiala 2006). Spore morphology might also confirm the establishment of the new species K. unicapsula n. sp. to some degree. Morphological characters of the spores can show some variability within the same species as the number of polar capsules and valve cells vary in Kudoa (Egusa 1986; Whipps et al. 2003) or the length of floating appendages in actinosporeans (Hallett et al. 2002). Thus, spore morphology is the most limited of all proofs for establishing a new species. However, in the present case, the morphological differences between K. unicapsula n. sp. and K. trifolia do not represent a simple multiplication or elongation of a spore part but would require a combination of structural changes, i.e. shrinking of a strongly enlarged polar capsule and considerable expansion of a valve cell with allocation of the sporoplasm within it. Such drastic changes have not been observed between spores of the same species to date. As both species inhabit the same mugilid hosts in the same geographical area (Spanish Mediterranean), it is possible that speciation of these two representatives of Kudoa has been going on for a relatively short time, resulting in small genetic differences and morphological speciation of K. unicapsula sp. n., and K. trifolia might be more pronounced than the one observed on a molecular basis. Summarizing, the combination of 28S rDNA data, host tissue location and morphology lead us to conclude that K. unicapsula n. sp. should be considered a new valid species.
To observe and interpret small molecular differences between species or geographical isolates, genes other than the 18S rDNA, which is commonly used for myxozoan, should be tested for their suitability for phylogenetic studies. Whipps and Kent (2006) made a successful approach involving the more variable ribosomal gene regions (28S, ITS1 and ITS2) as well as sequences of heat shock protein 70 for a phylogeographic study of K. thyrsites isolates. In the current study, the 28S rDNA sequences published by these authors were analysed phylogenetically together with the sequences obtained for K. unicapsula n. sp. and K. trifolia. The tree topology produced by the analysis of Kudoa spp. 28S rDNA sequences differs from the one produced by 18S rDNA sequences (see, e.g. Holzer et al. 2006; Diamant et al. 2005), with notably higher confidence values obtained in the 28S rDNA analysis, which can partially be ascribed to the higher sequence variability between different species of Kudoa. This suggests that this gene region could be useful for the analysis of phylogenetic relationships within closely related species of Myxozoa.
References
Aseeva NL (2004) New species of Myxosporea from genus Kudoa (Myxosporea, Multivalvulida) found in muscles of some fishes of the Sea of Japan. Vestnik Zool 38(2):75–77 (in Russian)
Barta JR, Martin DS, Liberator PA, Dashkevicz M, Anderson JW, Feighner SD, Elbrecht A, Perkins-Barrow A, Jenkins MC, Danforth HD, Ruff MD, Profous-Juchelka H (1997) Phylogenetic relationships among eight Eimeria species infecting domestic fowl inferred using complete small subunit ribosomal DNA sequences. J Parasitol 83:262–271
Bunton TE, Poynton SL (1991) Kudoa sp. (Myxosporea, Multivalvulida) infection in juvenile white perch, Morone americana (Gmelin): histopathology and spore morphology. J Fish Dis 14:589–594
Diamant A, Ucko M, Paperna I, Colorni A, Lipshitz A (2005) Kudoa iwatai. (Myxosporea: Multivalvulida) in wild and cultured fish in the Red Sea: redescription and molecular phylogeny. J Parasitol 91:1175–1189
Dykova I, Avila EJF, Fiala I (2002) Kudoa dianae sp. n. (Myxosporea: Multivalvulida), a new parasite of bullseye puffer, Sphaeroides annulatus (Tetraodontiformes: Tetraodontidae). Folia Parasitol 49:17–23
Egusa S (1986) The order Multivalvulida Shulman, 1959 (Myxozoa: Myxosporea): a review. Fish Pathol 21:261–274
Egusa S, Shiomitsu T (1983) Two new species of the genus Kudoa (Myxosporea: Multivalvulida) from marine cultured fishes in Japan. Fish Pathol 18:163–171 (in Japanese)
Eszterbauer E (2004) Genetic relationship among gill-infecting Myxobolus species (Myxosporea) of cyprinids: molecular evidence of importance of tissue-specificity. Dis Aquatic Org 58:35–40
Fiala I (2006) The phylogeny of Myxospera (Myxozoa) based on small subunit ribosomal RNA gene analysis. Int J Parasitol 36(14):1521–1534
Gilchrist JDF (1924) A protozoal parasite (Chlorornyxurn thyrsites sp-n.) of the Cape sea-fish, the “snoek” (Thyrsites atun, Euphr.). Trans R Soc S Afi 11:263–273
Hallett SL, Atkinson SD, El-Matbouli M (2002) Molecular characterisation of two aurantiactinomyxon (Myxozoa) phenotypes reveals one genotype. J Fish Dis 25:627–631
Holzer AS, Sommerville C, Wootten R (2004) Molecular relationships and phylogeny in a community of myxosporeans and actinosporeans based on their 18S rDNA sequences. Int J Parasitol 34:1099–1111
Holzer AS, Blasco-Costa I, Sarabeev VL, Ovcharenko MO, Balbuena JA (2006) Kudoa trifolia sp. n.—molecular phylogeny suggests a new spore morphology and unusual tissue location for a well-known genus. J Fish Dis 29:743–755
Kalavati C, Anuradha I (1993) Two new species of myxosporeans infecting Valamugil cunnesius in Visakhaptnam harbour, east coast of India. Uttar Pradesh J Zool 13:148–152
Kallert DM, Eszterbauer E, El-Matbouli M, Erseus C, Haas W (2006) The life cycle of Henneguya nuesslini Schuberg & Schröder, 1905 (Myxozoa) involves a triactinomyxon–type actinospore. J Fish Dis 28:71–79
Kostadinova AK, Ovcharenko MO, Balbuena JA, Sarabeev VL (2004) Sampling procedures for mullet parasites. Available at: http://cetus.uv.es/mullpardb/SempPrD.html
Lin D, Hanson LA, Pote LM (1999) Small subunit ribosomal RNA sequence of Henneguya exilis (Class Myxosporea) identifies the actinosporean stage from an oligochaete host. J Eukaryot Microbiol 46:66–68
Lom J, Arthur JR (1989) A guideline for the preparation of species descriptions in Myxosporea. J Fish Dis 12:151–156
Lom J, Dyková I (1992) Protozoan parasites of fishes. Elsevier, Amsterdam
Lom J, Rohde K, Dykova I (1992) Studies on protozoan parasites of Australian fishes. I. New species of the genera Coccomyxa Leger et Hess, 1907, Ortholinea Shulman, 1962 and Kudoa Meglitsch, 1947 (Myxozoa, Myxosporea). Folia Parasitol 39:289–306
Moran JDW, Whitaker DJ, Kent ML (1999) A review of the myxosporean genus Kudoa Meglitsch, 1947, and its impact on the international aquaculture industry and commercial fisheries. Aquaculture 172:163–196
Naidenova NN, Gaevskaya AV (1991) Kudoa mirabilis sp. n. (Myxosporea, Multivalvulea) from musculature of Indian Ocean cutlass fish. Zool Zh 70(5):131–133 (in Russian)
Perard C (1928) Su rune maladie du maquereau (Scomber scomber L.) due á une Myxosporidie: Chloromyxum histolyticum n. sp. C R Acad Sci Paris 186:108–110
Ronquist F, Huelsenbeck JP (2003) MrBayes 3: Bayesian phylogenetic inference under mixed models. Bioinformatics 19:1572–1574
Sarkar NK, Mazumder SK (1983) Studies on myxosporidian parasites (Myxozoa: Myxosporea) from marine fishes in West Bengal, India. I. Description of 3 new species from Tachysurus spp. Arch Protistenkd 127:59–63
Sarkar NK, Chaudry SR (1996) Kudoa cascasia. sp. n. (Myxosporea: Kudoidae) parasitic in the mesentery of Sicamugil cascasia (Ham.) from Hooghly estuary of west Bengal, India. Acta Protozool 35:335–338
Schulman SS, Donec ZS, Kovaleva AA (1997) Class of myxosporeans (Myxosporea) of the world fauna, vol. 1. Nauka, St. Petersburg (in Russian)
Tamura K, Nei M (1993) Estimation of the number of nucleotide substitutions in the control region of mitochondrial DNA in humans and chimpanzees. Mol Biol Evol 10:512–526
Thompson JD, Gibson TJ, Plewniak F, Jeanmougin F, Higgins DG (1997) The CLUSTAL_X windows interface: flexible strategies for multiple sequence alignment aided by quality analysis tools. Nucleic Acids Res 25:4876–4882
Tripathi YR (1951) Studies on parasites of Indian fishes. I. Protozoa myxosporidia together with a check list of parasitic protozoa described from Indian fishes. Rec Indian Mus 50:63–89
Vávra J, Maddox JV (1976) Methods in microsporidiology. In: Bulla JLA, Cheng TC (eds) Comparative pathobiology, vol. 1. Plenum, New York, pp 281–320
Whipps CM, Kent LM (2006) Phylogeography of the cosmopolitan marine parasite Kudoa thyrsites (Myxozoa: Myxosporea). J Eukaryot Microbiol 53:364–373
Whipps CM, Adlard JRD, Bryant MS, Robert JG, Lester RJG, Findlay V, Michael L, Kent ML (2001) First report of three Kudoa species from eastern Australia: Kudoa thyrsites from Mahi mahi (Coryphaena hippurus), Kudoa amamiensis and Kudoa minithyrsites n. sp. from Sweeper (Pempheris ypsilychnus). J Euk Microbiol 50(3):215–219
Whipps CM, Adlard RD, Bryant MS, Kent ML (2003) Two unusual myxozoans, Kudoa quadricornis n. sp. (Multivalvulida) from the muscle of goldspotted trevally (Carangoides fulvoguttatus) and Kudoa permulticapsula n. sp (Multivalvulida) from the muscle of Spanish mackerel (Scomberomorus commersoni) from the Great Barrier Reef, Australia. J Parasitol 89:168–173
Whipps CM, Grossel G, Adlard RD, Yokoyama H, Bryant MS, Munday BL, Kent ML (2004) Phylogeny of the multivalvulidae (Myxozoa: Myxosporea) based on comparative ribosomal DNA sequence analysis. J Parasitol 90:618–622
Whitaker DJ, Kabata Z, Margolis L (1994) Myxosporean parasites and their potential impact on the aquaculture industry, with emphasis on Kudoa species. In: Conley DC (ed) Kudoa workshop proceedings, Nanaimo, BC. Aquaculture Industry Development Report 94-01. British Columbia Ministry of Agriculture, Fisheries and Food, Victoria, pp 2–7
Yokoyama H, Itoh N (2005) Two multivalvulid Myxozoans causing postmortem myoliquefaction: Kudoa megacapsula n. sp. from red barracuda (Sphyraena pinguis) and Kudoa thyrsites from splendid alfonso (Beryx splendens). J Parasitol 91(5):1132–1137
Yurakhno VM (1991) New species of myxosporidia from fishes of the Black Sea. Parasitologiya, 25(2):104–109 (in Russian)
Acknowledgements
We would like to thank Isabel Blasco-Costa at the Cavanilles Institute of Biodiversity and Evolutionary Biology, University of Valencia, for the help with the collection of material. This research was supported by the INTAS grant (project number 03-51-5998). A.S.H. benefited from a postdoctoral Marie-Curie Fellowship of the European Union (MTKD-CT-2004-003175). We assure that the experiments comply with the current laws of the country in where they were performed.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Yurakhno, V.M., Ovcharenko, M.O., Holzer, A.S. et al. Kudoa unicapsula n. sp. (Myxosporea: Kudoidae) a parasite of the Mediterranean mullets Liza ramada and L. aurata (Teleostei: Mugilidae). Parasitol Res 101, 1671–1680 (2007). https://doi.org/10.1007/s00436-007-0711-8
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00436-007-0711-8