
Abstract
Adult trematodes of the genus Stegodexamene Macfarlane, 1951 were recorded from the intestines of two species of freshwater eels, the Pacific short-finned eel Anguilla obscura Günther and the speckled longfin eel Anguilla reinhardtii Steindachner from the Pouembout and La Foa Rivers, New Caledonia, South Pacific. They were identified as Stegodexamene anguillae Macfarlane, 1951, a species previously reported only from eels in New Zealand. The morphology of adult S. anguillae seems to be practically identical with that of the later described Stegodexamene callista Watson 1984 from eels in Australia. The finding of S. anguillae in A. obscura and A. reinhardtii in New Caledonia represents new geographical and host records. In addition to this trematode, two larval specimens of a cestode of the order Proteocephalidea were found in the intestine of one A. obscura from the Pouembout River. No monogenean was found in eels of New Caledonia.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Although New Caledonia is an interesting area from the zoogeographical point of view, with a high degree of endemism, almost no attention has been paid to the parasites of native fishes in freshwater habitats. As far as we know, the only two papers dealing with helminths of freshwater fishes in New Caledonia are those recently published dealing with the occurrence of camallanid nematodes, a new species of Procamallanus Baylis, 1923 from eels in New Caledonia and some other Pacific islands, and Camallanus cotti Fujita, 1927, a pathogenic parasite of fishes introduced into New Caledonia (Moravec et al. 2006; Moravec and Justine 2006). A few lepocreadiid trematodes have recently been reported from New Caledonian marine fishes (Bray and Justine 2006).
In July 2003 and September 2004, samples of eels from two localities in New Caledonia, the Pouembout and La Foa Rivers and swamps near Ponérihouen, were examined for the presence of helminth parasites. Results of the evaluation of nematodes recorded have already been published (Moravec et al. 2006), whereas those concerning the trematodes and cestodes are treated below.
Materials and methods
Eels were caught by local fishermen in the Pouembout River near Pouembout (west coast, Province Nord) and by Dr. C. Pöllabauer by electrofishing in the La Foa River near La Foa (west coast, Province Sud), both localities in the mainland of New Caledonia, South Pacific. Some eels from Pouembout were kept in captivity for several months before their examination. Three native species of eels were obtained and examined in September 2004: Pacific short-finned eel, Anguilla obscura Günther (three specimens from Pouembout; body length 67–88 cm), speckled longfin eel, Anguilla reinhardtii Steindachner (two specimens from Pouembout and 11 from La Foa; body length 71–72 and 18–45 cm, respectively), and the giant mottled eel, Anguilla marmorata Quoy et Gaimard (two specimens from La Foa; body length 28–45 cm). The eels were transported alive to the laboratory of the Institut de Recherche pour le Développement (IRD) in Nouméa, killed, and immediately examined for the presence of helminth parasites. One specimen of A. reinhardtii from Pouembout (body length 104 cm) and three specimens of A. obscura from swamps near Ponérihouen, east coast, Province Nord (body length 52–76 cm) were examined in July 2003 but had no digeneans. No parasites were found in the examined A. marmorata. The recovered trematodes and cestodes were washed in physiological saline and then fixed in hot 4% formaldehyde solution or, slightly pressed under the cover slip, in cold 4% formaldehyde solution. For light microscopic examination, the specimens were stained in carmine, dehydrated through an ethanol series, and mounted in Canada balsam as permanent slides. Drawings were made with the aid of a Zeiss drawing attachment. The scientific names of fishes follow Fishbase (Froese and Pauly 2006).
Results
The following observations were made for Stegodexamene anguillae Macfarlane, 1951 (Trematoda: Lepocreadiidae).
Description
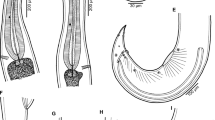
As shown in Fig. 1a–f, in our eight gravid specimens from A. reinhardtii (measurements of two largest specimens from A. obscura are in parentheses), the body is oval to elongate-oval, usually somewhat tapering towards anterior end, with the maximum width at about its middle. The body length is 1.09–3.58 (2.77–3.17) mm and the maximum width 394–721 (707–856) μm. The tegument is covered with minute spines. Remnants of the eyespots of cercariae (as scattered pigment granules) are present on the dorsal side. The oral sucker is oval, subterminal, 111–190 (150–204) μm long, and 138–218 (190–218) μm wide. The ventral sucker is circular to somewhat oval, situated approximately in the anterior third of the body, 126–204 (177–190) μm long, and 135–218 (177–231) μm wide; the size ratio of the oral to the ventral suckers is 0.95–1.14: 1. The prepharynx is very short and not visible in all specimens. The pharynx is oval, muscular, 51–105 (95–111) μm long, and 60–102 (95–120) μm wide, with a lobular anterior margin. The esophagus is narrow and 69–408 (299–313) μm long. The intestinal bifurcation is in the posterior part of the fore body, somewhat anterior to the ventral sucker (except for the smallest specimen where it is at the level of the anterior edge of ventral sucker). The caeca are narrow, ending blindly close to the posterior extremity. The testes are two, oval to almost spherical, nearly equal in size, oblique, at about the middle of body. The anterior testis is situated on the opposite (left) side of the ovary, somewhat posterior to the ovary or nearly at its level, while the posterior testis is in about the same longitudinal line with the ovary. The anterior testis is 78–231 (231–326) μm long and 102–313 (272) μm wide; the posterior testis is 99–272 (299–313) μm long and 114–272 (245–354) μm wide. The external seminal vesicle is absent. The cirrus sac is claviform, 255–707 (435–503) μm long and 68–177 (136–204) μm wide, curved around the right side of the ventral sucker, extending into the hind body. It contains a bipartite seminal vesicle, a pars prostatica, and an invaginated cirrus; the posterior part of the seminal vesicle is oval and much larger than the anterior one; the cirrus is with small patches of tiny spines near its end (Fig. 1f), which are invisible in most specimens. The genital pore lies ventrally and slightly to the left of the midline between the ventral sucker and the intestinal bifurcation. The metraterm is distinct; glandular cells surrounding the exit of the metraterm and cirrus are present. The ovary is oval to spherical, 57–177 (136–204) μm long, and 84–204 (177–204) μm wide, pretesticular, situated somewhat asymmetrically to the right of the midline. The seminal receptacle is well-developed, 39–286 (258–394) μm long, and 30–136 (150–299) μm wide, located to the right from the ovary. Vitelline follicles are numerous, surround caeca dorsally, ventrally, and externally, and become confluent posterior to testes, extending from the ovary nearly to the end of the body. The vitelline follicles are poorly developed in the smallest specimens (Fig. 1a). Uterine loops are anterior to the ovary and contain 2–38 (5–12) eggs. The eggs are operculated, yellow-brown, 60–69 (63–69) μm long, and 36–45 (36–45) μm wide. The excretory bladder is tubular, long, somewhat curved between testes, extending anteriorly to at least the level of the intestinal bifurcation.

Stegodexamene anguillae Macfarlane, 1951 (a–f). a Smallest specimen with eggs, b large specimen with many eggs, c largest specimen, d small specimen with few eggs, e cirrus sac, f part of inverted cirrus with patches of spines inside cirrus sac). g Proteocephalidea gen. sp. larva (scale bars: a–d = 1 mm; e, g = 200 μm; f = 50 μm)
Hosts
The hosts are the eels A. obscura and A. reinhardtii (Anguillidae, Anguilliformes), and the site of infection is the intestine. Localities are the Pouembout River near Pouembout, Province Nord (A. obscura, A. reinhardtii) and the La Foa River near La Foa, Province Sud (A. reinhardtii) in New Caledonia. The rate of infection in the localities was as follows. In the Pouembout River, the prevalence in A. obscura was one eel infected/three eels examined, and the intensity was 7 specimens; the prevalence in A. reinhardtii was 2/3, with an intensity of 14 and 25 specimens. In the La Foa River (A. reinhardtii), prevalence was 1/11 and intensity was 2 specimens. In the Ponérihouen swamps (A. obscura), prevalence was 0/3. The voucher specimens are deposited at the Muséum National d’Histoire Naturelle, Paris, France (Cat. Nos. JCN1273, JCN1274, JCN1275, JCN1311) and at the Institute of Parasitology, Biology Centre of the Academy of Sciences of the Czech Republic, České Budějovice (Cat. No. D-607).
In addition to S. anguillae, two cestode larvae of an unidentified genus and family, referable to the order Proteocephalidea, were recorded from A. obscura caught in the Pouembout River near Pouembout, northern New Caledonia.
The two Proteocephalidea gen. sp. larvae were found free in the intestine of one of three eels examined. Their total body length including the scolex is 789 and 843 μm. The length of the scolex is 136 and150 μm and its maximum width is 231 and 286 μm, while the maximum width of the posterior part of the body (posterior to the scolex) is 204 and 340 μm. As shown in Fig. 1 (G), the scolex is with four small, almost circular suckers 84–99 μm in diameter. The top of the scolex is with a simple apical sucker measuring 36–60 × 63–75 μm.
The morphology of the scolex of these larvae and the freshwater environment suggest that they belong to the order Proteocephalidea. Adults of this order are parasites of freshwater fishes, amphibians, and reptiles (Khalil et al. 1994). The voucher specimens are deposited at the Muséum National d’Histoire Naturelle, Paris, France (Cat. No. JCN1277) and at the Institute of Parasitology, Biology Centre of the Academy of Sciences of the Czech Republic, České Budějovice (Cat. No. C-436).
Discussion
The trematode genus Stegodexamene Macfarlane, 1951 was erected to accommodate the species S. anguillae Macfarlane, 1951, an intestinal parasite of freshwater eels originally described from Anguilla dieffenbachii Gray and Anguilla australis schmidti Philipps from New Zealand (Macfarlane 1951); from these two hosts in New Zealand, S. anguillae was also reported by Macfarlane (1952), Hewitt and Hine (1972), and Rid (1973). However, according to Froese and Pauly (2006), the latter fish occurs only in New Caledonia; therefore, it is probable that the second host was in fact A. australis australis Richardson, distributed on the coast of Australia and New Zealand, extending north to New Caledonia (Froese and Pauly 2006). Later S. anguillae was reported in New Zealand as a common parasite of the endemic New Zealand longfin eel, A. dieffenbachii, and the shortfin eel, A. australis (see, e.g., Manter 1954; Hine 1980; Hine and Francis 1980; Blair 1984; Graynoth and Taylor 2000).
Srivastava (1962) erected the genus Rhynchocreadium Srivastava, 1962 for the newly described species Rhynchocreadium aculeatum Srivastava, 1962 from the freshwater spiny eel Macrognathus aculeatus (Bloch) in India. Yamaguti (1971) took Rhynchocreadium for a subgenus of Stegodexamene, listing in it two species, S. aculeatum Srivastava, 1962 and S. singhia Pershad, 1965, both from the same host species (M. aculeatus) in India. At present, Rhynchocreadium Srivastava, 1962 is considered a junior synonym of the genus Allocreadium Looss, 1900 (family Allocreadiidae; see Caira and Bogéa 2005).
Consequently, in addition to the type species of Stegodexamene, the only other representative of this genus is Stegodexamene callista Watson, 1984 described from the speckled longfin eel A. reinhardtii Steindachner in Australia (Watson 1984). According to Watson (1984), adults of this species are very similar to those of S. anguillae, differing from them in a somewhat smaller body size; a less extensive pars prostatica; their excretory bladder, which may extend even anterior to the intestinal bifurcation (not reported for S. anguillae); in more numerous flame cells (50 vs 42); and the presence of minute cirrus spines. However, all these differences are questionable.
The range of the body length of gravid specimens of the present New Caledonian material corresponds to both species. Regarding the number of flame cells, which can be studied with difficulties in living specimens only, Macfarlane (1951) believed that the flame cell pattern of S. anguillae, which he illustrated, was incomplete. As noted by Watson (1984), the minute cirrus spines were not reported by Macfarlane (1951), but these are difficult to observe and are easily overlooked; they were not visible in most mounted specimens of the present material. New Zealand adult specimens of S. anguillae obtained from A. australis were also studied by Watson (1984), who had not observed any morphological differences between S. anguillae and S. callista.
Therefore, the only interspecific differences between S. anguillae and S. callista should be in the structure of cercariae, namely in the number and arrangement of their caudal setae, provided that the observations on the life cycles published by Macfarlane (1951) and Watson (1984) are reliable. Nevertheless, a comparison of the New Zealand and Australian trematodes by molecular methods and new comparative studies of their life cycles conducted preferably by the same reseacher are highly desirable to confirm the existence of S. callista as an independent species.
Because the body length in Stegodexamene specimens of the present material attains nearly the maximum length of S. anguillae, their morphology is in accordance with the description of this species, and there may be some doubts concerning the validity of another species, S. callista, which may parasitize the same host species (A. australis) as S. anguillae; they are identified as S. anguillae. This species, previously known only from New Zealand, is now reported for the first time from New Caledonia and A. obscura and A. reinhardtii represent its new host records.
Macfarlane (1951) mentions that in New Zealand, adults of S. anguillae occur in the intestines of eels longer than 35 cm (ca. 7 years old). Also in New Caledonia, this parasite was found only in larger eels (body length 45–72 cm in A. reinhardtii and 88 cm in A. obscura), whereas they were absent from ten smaller A. reinhardtii (body length 18–40 cm) and two A. marmorata (body length 28 and 45 cm) from the La Foa River. This is associated with the trematode’s life cycle and the ecology of eels, where the source of infection for eels are some forage fishes (in New Zealand mainly of the families Eleotridae and Galaxiidae) serving as the second intermediate host of this parasite; metacercariae of S. anguillae encysted in fish tissues may be progenetic, producing viable eggs capable of further development (Holton 1984; Poulin and Lefebvre 2005). The first intermediate hosts of S. anguillae in New Zealand are the gastropods Potamopyrgus antipodarum (Gray) and Potamopyrgus badia Gould (Hydrobiidae) (Macfarlane 1951). Field surveys show that in natural New Zealand eel populations, 26–78% of A. australis and 25–40% of A. dieffenbachii are parasitized by adult S. anguillae (Rid 1973; Hine 1980). The present data indicate that S. anguillae is a common parasite of eels also in New Caledonia. Although S. anguillae co-occurred with the nematode Procamallanus pacificus Moravec, Justine, Würtz, Taraschewski et Sasal, 2006 in A. obscura and A. reinhardtii of the Pouembout River (Moravec et al. 2006), this nematode parasite was completely absent from eels of the La Foa River.
Although negative results are often not reported in parasitological papers, we believe it is worth mentioning that no swim bladder nematodes or monogeneans on gills were found in specimens of all three species of eels collected in New Caledonia. One of the A. obscura from Pouembout had small white dots on the gills, which contained myxozoan spores.
References
Blair D (1984) A checklist and bibliography of parasites of New Zealand freshwater fish. Mauri Ora 11:5–50
Bray RA, Justine JL (2006) Hypocreaduim toombo n. sp. (Digenea: Lepocreadiidae) in the yellow-spotted triggerfish Pseudobalistes fuscus (Perciformes: Balistidae) and additional lepocreadiids parasitizing fishes from the waters off New Caledonia. Zootaxa 1326:37–44
Caira JN, Bogéa T (2005) Family Allocreadiidae Looss, 1902. In: Jones A, Bray RA, Gibson DI (eds) Keys to the Trematoda, vol 2. CABI Publishing, Wallingford, UK
Froese R, Pauly D (eds) (2006) FishBase. http://www.fishbase.org, version 01/07/2006
Graynoth E, Taylor MJ (2000) Influence of different rations and water temperatures on the growth rates of shortfinned eels and longfinned eels. J Fish Biol 57:681–699
Hewitt GC, Hine PM (1972) Checklist of parasites of New Zealand fishes and of their hosts. N Zealand J Mar Freshw Res 6:69–114
Hine PM (1980) Distribution of helminths in the digestive tracts of New Zealand freshwater eels. 1. Distribution of digeneans. N Zealand J Mar Freshw Res 14:329–338
Hine PM, Francis RICC (1980) Distribution of helminths in the digestive tracts of New Zealand freshwater eels. 3. Interspecific associations and conclusions. N Zealand J Mar Freshw Res 14:349–356
Holton AL (1984) Progenesis as a means of abbreviating life histories in two New Zealand trematodes, Coitocaecum parvum Crowcroft, 1945 and Stegodexamene Anguillae Macfarlane, 1951. Mauri Ora 11:63–70
Khalil LF, Jones A, Bray RA (eds) (1994) Keys to the cestode parasites of vertebrates. CAB International, Wallingford, UK
Macfarlane WV (1951) The life cycle of Stegodexamene anguillae n. g., n. sp., an allocreadiid trematode from New Zealand. Parasitology 41:1–10
Macfarlane WV (1952) Bionomics of two trematode parasites of New Zealand eels. J Parasitol 38:391–397
Manter HW (1954) Some digenetic trematodes from fishes of New Zealand. Trans Roy Soc N Z 82:475–568
Moravec F, Justine J-L (2006) Camallanus cotti (Nematoda: Camallanidae), an introduced parasite of fishes in New Caledonia. Folia Parasitol 53:287–296
Moravec F, Justine J-L, Würtz J, Taraschewski H, Sasal P (2006) A new species of Procamallanus (Nematoda: Camallanidae) from Pacific eels (Anguilla spp.). J Parasitol 92:130–137
Poulin R, Lefebvre F (2005) Alternative life-history and transmission strategies in a parasite: first come, first served? Parasitology 132:135–141
Rid LE (1973) Helminth parasites of the long-finned, Anguilla dieffenbachii, and the short-finned eel, A. australis. Mauri Ora 1:99–106
Srivastava CB (1962) A new allocreadiid trematode, Rhynchocreadium aculeatum gen. et sp. nov., from the freshwater eel, Rhanchobdella aculeata Bloch. Indian J Helminthol 14:1–4
Watson RA (1984) The life cycle and morphology of Tetracerasta blepta, gen. et sp. nov., and Stegodexamene callista, sp. nov. (Trematoda: Lepocreadiidae) from the long-finned eel, Anguilla reinhardtii Steindachner. Aust J Zool 32:177–204
Yamaguti S (1971) Synopsis of digenetic trematodes of vertebrates, vol. I, II. Keigaku Publishing Co., Tokyo
Acknowledgements
Yann Pellequer (Direction du Développement Économique et de l’Environnement, Province Nord, Poindimié, New Caledonia) kindly provided the eels from Pouembout and Ponérihouen. Dr. Christine Pöllabauer (Erbio, Nouméa, New Caledonia) kindly organized two electrofishing expeditions to the La Foa and Dumbéa Rivers and is thanked for her collaboration, fish identification, and fruitful discussions. Julie Mounier, Anaïs Guérin, Audrey Guillou, Charles Beaufrère, and Eric Bureau, students, participated in the fishing expeditions and parasitological surveys. Angelo Di Matteo (IRD, Nouméa) is thanked for technical help. Thanks are also due to Irena Husáková from the Institute of Parasitology, Biology Centre, ASCR in České Budějovice for her technical assistance and help with the preparation of illustrations. This study was partly supported by grant no. 524/06/0170 from the Grant Agency of the Czech Republic and by the research projects of the Institute of Parasitology, ASCR (Z60220518 and LC522).
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Moravec, F., Justine, JL. Stegodexamene anguillae (Digenea: Lepocreadiidae), an intestinal parasite of eels (Anguilla spp.) in New Caledonia. Parasitol Res 100, 1047–1051 (2007). https://doi.org/10.1007/s00436-006-0386-6
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00436-006-0386-6