
Abstract
Several factors capable of affecting the amount of resources available to an individual parasite, such as the number of other parasites in the host or host quality, may cause variability in reproductive success among parasites. Variation in egg output and mean egg volume was investigated among adult females of the nematode Graphidioides subterraneus, parasitic in the herbivorous subterranean rodent Ctenomys talarum (Octodontidae). Female nematode body size correlated strongly with the number of eggs produced. However, neither host body mass nor the number of other nematodes per host had any influence on the number or volume of eggs produced by the parasites. There was also no evidence for a trade-off between the number of eggs produced and mean egg volume among female nematodes. All these results suggest that resource supply to individual worms is not limited by host size or by the number of conspecific parasites vying for the same resources, despite the 30-fold variation in intensity of infection and the twofold variation in host body mass observed in the present study. Instead, resource availability does not appear to constrain reproduction in G. subterraneus, with its host providing a stable, predictable environment.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
The reproductive ecology of parasites has received some attention over the years, but the determinants of reproductive output by individual parasites remain only poorly understood (Poulin 1998). Variation in body size among conspecific parasites explains much of the variability in reproductive output among individuals, but other factors also matter. On the one hand, different host individuals vary in quality, i.e. in the amount or quality of nutrients or other resources that they provide to growing and reproducing parasites. Some studies have demonstrated that parasite reproductive output is affected by the general quality of the host, based on its age, size, diet or immune status (Ito et al. 1986; Quinnell 1988; Poulin 1996). On the other hand, epidemiological models suggest that for parasite abundance to be regulated in a density-dependent manner, the number of conspecifics sharing a host should influence the average per capita fecundity of parasites (see Anderson and May 1978; Anderson 1993). Indeed, experimental studies suggest that intensity of infection has a negative effect on per capita egg production in many helminths (Krupp 1961; Jones et al. 1989), and the evidence for density-dependent regulation is generally convincing (Keymer 1982; Quinnell et al. 1990; Shostak and Scott 1993). What remains to be assessed, however, is the relative role of all these factors in determining the individual reproductive output of parasites: what is most important, the direct effect of parasite body size, or the indirect effects of host quality and/or the number of conspecifics?
Another aspect of individual variation in reproductive output among parasites that has received little attention is the partitioning of reproductive output between quantity (egg number) and quality (egg volume) of offspring. A trade-off is expected between egg number and egg volume because energy allocated to producing more eggs cannot simultaneously be used to provide more resources to each egg. The net result is a negative correlation between egg number and size. Across individual parasites, a continuum of strategies from the production of many small eggs to the production of few large ones might be expected. This trade-off has been documented in a range of animal taxa (e.g. Elgar 1990; Guisande et al. 1996; Christians 2000; Kinnison et al. 2001; Brown 2003). This trade-off should also apply to parasitic organisms. Thus, at an interspecific level, there is a negative correlation between relative egg size and the number of eggs per clutch in some parasitic taxa like copepods (Poulin 1995), but not in others such as nematodes (Skorping et al. 1991). However, whether or not there exists a trade-off between egg size and egg number within species of parasites is unknown. Parasites are not constrained by resource supply, and perhaps they escape the trade-off at an intraspecific level. A recent study indicates that this is the case for an ectoparasitic copepod of fish (Timi et al. 2005), but information on endoparasitic helminths is lacking. Inside their hosts, parasite individuals have a resource supply that is practically infinite. Thus, high resource availability may allow parasites to allocate resources simultaneously to both egg number and egg size. Hence, the high resource availability experienced by parasites may neutralise the trade-off between the number and size of eggs.
Here, we explore intraspecific and inter-individual variation in reproductive output in the nematode G. subterraneus, recently described by Rossin et al. (2005), parasitic in the herbivorous subterranean rodent Ctenomys talarumThomas, 1898 (Octodontidae), from Argentina. C. talarum lives in permanently sealed individual burrow systems, and although most of its activities are restricted to its tunnel systems, animals make brief surface excursions to collect plant material. Individuals of all ages (except young that occupy their mother’s tunnel system) are sedentary and maintain exclusive territories (Busch et al. 1989). Since most of the life of C. talarum is spent underground, certain phases of its life history and ecology are poorly studied, especially its parasite fauna. In a previous study on endoparasites of C. talarum from another locality (Necochea, Provincia de Buenos Aires, Argentina) only two parasitic nematodes were found, Heligmostrongylus sp. and Trichuris pampeana (Rossin and Malizia 2002, 2005).
This host-parasite system is ideal for such an investigation for at least three reasons. First, G. subterraneus is the only helminth species in the stomach of C. talarum, thus eliminating any potential interspecific effects. Second, this nematode produces relatively few eggs, therefore it is possible to quantify egg productions of individual females, as opposed to previous studies in which fecal egg counts, subsequently divided by the number of worms in a host, are often used as estimates of mean egg production. Third, in our sample there was a 30-fold variation in intensity of infection, a threefold variation in individual worm volume, and a twofold variation in host body mass, providing an excellent opportunity to examine the effects of worm size, infection intensity and host quality on reproductive output in this nematode.
Our specific objectives were (1) to assess the influence of worm size, intensity of infection and host mass on the fecundity, mean egg volume and total reproductive output of the nematode G. subterraneus, and (2) to search for any sign of a trade-off between egg number and egg volume, and determine whether this trade-off is influenced by intensity of infection and host quality. The results of this study will provide an insight into which factors are most important in determining inter-individual variation in reproductive tactics among parasites.
Materials and methods
Eighty-one specimens of C. talarum were collected during 2000 and 2001 at Mar de Cobo, Buenos Aires Province, Argentina (37° 58′S, 57° 34′W). Individual hosts were weighted, dissected, and nematodes were collected from their stomach. Living nematodes were recovered from 12 parasitized hosts, fixed in 4% formaldehyde solution, and subsequently preserved in 70% ethanol.
All adult female worms, with fully developed eggs in the uterus, were taken from each host, cleared with lactophenol, and measured (length and width); when more than 10 adult females were present in a host, ten specimens were randomly selected and measured. For each female the number of fully developed eggs was counted, and five eggs (those closest to the vulva) were measured (length and width). Then, both body and egg volumes were calculated, in mm3, by equating their shape to that of a cylinder.
We obtained data on 78 worms from 12 host individuals. We made the reasonable assumption that the number of eggs found in utero was proportional to the rate at which eggs are produced, and thus to the fecundity of individual worms. For each worm, we calculated total reproductive output as the product of the number of eggs in utero and mean egg volume.
Two variables, number of conspecifics per host and number of eggs per worm, did not meet the assumptions of parametric tests and were log-transformed. Multiple regressions were used to estimate the effects of predictor variables on the three dependent variables (egg number, mean egg volume, and total reproductive output). The predictor variables were worm volume, number of conspecifics in the host, and host mass; in the analysis with mean egg volume as the dependent variable, egg number was added as a fourth predictor variable, thus allowing a test of the trade-off between fecundity and egg volume while controlling for variation in other variables.
Results
Both prevalence and mean abundance of G. subterraneus were low (24.7% and 5.0±20.7, respectively) in the host population. Among the 12 hosts analysed, intensity of infection ranged from 5 to 155 worms per host. Among the 78 female worms, body volume ranged from 2.24 mm3 to 6.73 mm3, and fecundity ranged from 4 to 202 eggs per female.
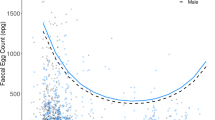
Neither host mass nor number of conspecifics had any significant influence on the number of eggs produced by a female. Host mass did tend to correlate positively with number of eggs, but this trend was only very weak (P=0.08). However, there was a strong positive relationship between number of eggs and female volume (Table 1, Fig. 1). In contrast, mean egg volume and total reproductive output did not covary significantly with neither of the independent variables (Table 1). There was a tendency for total reproductive output to correlate positively with the number of conspecifics, but this trend was rather weak, however (P=0.08).

Relationship between the size of females of Graphidioides subterraneus and the number of eggs in the uterus
There was also no relationship between egg number and mean egg volume, when the other variables were taken into consideration (Table 1). In other words, for a given body length, females that produce many eggs do not tend to produce relatively small eggs, and vice versa (Fig. 2).

Relationship between the number of eggs produced by females of Graphidioides subterraneus and the mean volume of eggs per female
Discussion
As in most animals, fecundity of parasites correlates with body size (Poulin 1998) as shown here by the strong positive relationship between number of eggs and female volume in G. subterraneus. In fact, previous studies have shown that parasitic nematodes show variation in egg production as a function of body size, both at the intraspecific level and across taxa (Morand 1996). This can be interpreted as an evolutionary consequence of selection acting to maximize fecundity. In parasitic nematodes high rates of egg production and lifetime fecundity have been favoured through long prepatency periods, resulting in longer adult life span and large body size (Morand 1996; Poulin 1998).
Parasite reproductive output can be affected by the variability in host quality, as related to the latter’s age, size, diet or immune status (Ito et al. 1986; Quinnell 1988; Poulin 1996). We did find a weak association between host mass and parasite egg output, but it was not statistically significant. Thus, despite the twofold variation in host body mass observed in our sample, it appears that different individuals of C. talarum do not vary in quality as hosts for G. subterraneus. Any slight variation in the amount or quality of nutrients or other resources that they provide to parasites had no detectable influence on the number of eggs, the mean egg volume and the total reproductive output of female nematodes. Thus, this host species can be considered as a stable and homogeneous habitat for parasites.
The stability of C. talarum as an environment for parasites could be a consequence of their ecological habits. Octodontid rodents of the genus Ctenomys are the dominant mammals exploiting the subterranean niche (Nevo 1979; 1991). Representatives of this genus are distributed in a wide variety of soils, altitudes and climates. This habitat heterogeneity is counterbalanced by the more stable and predictable microclimatic conditions of humidity and temperature of the permanently sealed burrow systems in which these rodents live, ensuring a relative independence from the external conditions (Malizia, 1998). Ctenomys talarum from Mar de Cobo shares with other species of the genus many ecological attributes related to the subterranean life style. In that sense, they are K-strategists, having relatively slow development, relatively long lives, late sexual maturity, long gestation periods, small litter sizes, low recruitment, low mortality and, therefore, relatively constant population densities (65 individuals/ha) (Busch et al. 1989). Furthermore, this rodent inhabits natural coastal grasslands characterized by a high plant biomass and hence high productivity, which together with low population density of rodents ensures a constant supply of nutrients for both hosts of all ages and their parasites. Indeed, as G. subterraneus lives at low densities in the hosts, particularly if taking into account the volume of parasites relative to the volume of the host’s stomach, the number of conspecifics sharing a host, and therefore potential intraspecific competition, is not constraining the availability of resources invested in per capita parasite fecundity. Thus, parasite abundance is not regulated in a density-dependent manner. Indeed, although we observed a tendency for total reproductive output of nematodes to correlate with the number of conspecifics per host, this trend was not only very weak (P=0.08), but it was also positive.
Although the partitioning of reproductive output between quantity (egg number) and quality (egg volume) of offspring among parasites has received little attention, a trade-off between egg number and egg volume is expected. This result can be explained by the fact that the energy allocated to provide more resources to each egg cannot simultaneously be used to produce more eggs, resulting in a negative correlation between egg number and size. At an interspecific level, there is a negative correlation between relative egg size and number of eggs per clutch in some parasitic taxa like copepods (Poulin 1995), but not in others such as nematodes (Skorping et al. 1991). To our knowledge, this is the first study of intraspecific reproductive trade-offs in parasitic nematodes and the second one in parasitic organisms. A previous study at the intraspecific level on copepods parasitic on fish (Timi et al. 2005), showed that despite a high variability in body size of female copepods, there was no association between egg number and egg volume once effects of body size were taken into account. Similar results were obtained in the present study, supporting the idea that high resource availability may overcome any conflict regarding resource allocation between the number and size of offspring in parasites. In contrast to other animal taxa, individual female parasites, at least nematodes and copepods of the two species so far studied, appear capable of investing resources in both number and size of eggs simultaneously. In other words parasites seem not to be constrained by resource supply and thus they can escape the trade-off at an intraspecific level.
It is concluded that G. subterraneous is not constrained by resource availability and all individuals have a resource supply that is practically infinite. Therefore, only worm size, but not intensity of infection nor host quality, influences their reproductive success. This could be due to the characteristics of both the ecological traits of their host and their demographic attributes. However, the fact that similar intraspecific trends were also found among ectoparasitic copepods (Timi et al. 2005), could indicate that these aspects of reproductive ecology are common to parasites in general, and not a consequence of the particular subterranean life style. Further evidence on reproductive ecology of parasites living in hosts in unpredictable habitats, or in host populations whose structure is shaped by competition, are needed to test these hypotheses.
References
Anderson RM (1993) Modern parasitology. Blackwell, Oxford
Anderson RM, May RM (1978) Regulation and stability of host-parasite population interactions. I. Regulatory processes. J Anim Ecol 47:219–247
Brown CA (2003) Offspring size—number trade-offs in scorpions: an empirical test of the van Noordwijk and de Jong model. Evolution 57:2184–2190
Busch C, Malizia AI, Scaglia OA, Reig OA (1989) Spatial distribution and attributes of a population of Ctenomys talarum (Rodentia: Octodontidae). J Mamm 70:204–208
Christians JK (2000) Trade-offs between egg size and number in waterfowl: an interspecific test of the van Noordwijk and de Jong model. Funct Ecol 14:497–501
Elgar MA (1990) Evolutionary compromise between a few large and many small eggs: comparative evidence in teleost fish. Oikos 59:283–287
Guisande C, Sanchez J, Maneiro I, Miranda A (1996) Trade-off between offspring number and offspring size in the marine copepod Euterpina acutifrons at different food concentrations. Mar Ecol Prog Ser 143:37–44
Ito A, Kano S, Hioki A, Kasuya S, Ohtomo H (1986) Reduced fecundity of Hymenolepis nana due to thymus-dependent immunological responses in mice. Int J Parasitol 16:81–85
Jones JT, Breeze P, Kusel JR (1989) Schistosome fecundity: influence of host genotype and intensity of infection. Int J Parasitol 19:769–777
Keymer AE (1982) Density-dependent mechanisms in the regulation of intestinal helminth populations. Parasitology 84:573–587
Kinnison MT, Unwin MJ, Hendry AP, Quinn TP (2001) Migratory costs and the evolution of egg size and number in introduced and indigenous salmon populations. Evolution 55:1656–1667
Krupp IM (1961) Effects of crowding and of superinfection on habitat selection and egg production in Ancylostoma caninum. J Parasitol 47:957–961
Malizia AI (1998) Population dynamics of the fossorial rodent Ctenomys talarum (Rodentia: Octodontidae). J Zool Lond 244:545–551
Morand S (1996) Life-history traits in parasitic nematodes: a comparative approach for the search of invariants. Funct Ecol 10:210–218
Nevo E (1979) Adaptive convergence and divergence of subterranean mammals. Ann Rev Ecol Syst 25:127–144
Nevo E (1991) Evolutionary theory and processes of active speciation and adpative radiation in subterranean mole-rats, Spalax ehrenbergi superspecies in Israel. Evolutionary biology. Plenum , New York
Poulin R (1995) Clutch size and egg size in free-living and parasitic copepods: a comparative analysis. Evolution 49:325–336
Poulin R (1996) The evolution of life history strategies in parasitic animals. Adv Parasitol 37:107–134
Poulin R (1998) Evolutionary ecology of parasites: from individuals to communities. Chapman and Hall, London
Quinnell RJ (1988) Host age and the growth and fecundity of Hymenolepis diminuta in the rat. J Helminthol 62:158–162
Quinnell RJ, Medley GF, Keymer AE (1990) The regulation of gastrointestinal helminth populations. Philos Trans Rl Soc Lond B 330:191–201
Rossin A, Malizia AI (2002) Relationship between helminth parasites and demographic attributes of a population of the subterranean rodent Ctenomys talarum (Rodentia: Octodontidae). J Parasitol 88:1268–1270
Rossin MA, Malizia AI (2005) Redescription of Trichuris pampeana (Nematoda: Trichuridae) from the South American subterranean rodent Ctenomys talarum Thomas, 1898 (Rodentia: Octodontidae). J Parasitol 91:127–130
Rossin A, Timi JT, Malizia AI (2005) Graphidioides subterraneus n. sp. (Nematoda: Trichostrongylidae) from the South American subterranean rodent Ctenomys talarum Thomas, 1898 (Rodentia: Octodontidae). Parasite (in press)
Shostak AW, Scott ME (1993) Detection of density-dependent growth and fecundity of helminths in natural infections. Parasitology 106:527–539
Skorping A, Read AF, Keymer AE (1991) Life history covariation in intestinal nematodes of mammals. Oikos 60:365–372
Timi JT, Lanfranchi AL, Poulin R (2005) Is there a trade-off between fecundity and egg volume in the parasitic copepod Lernanthropus cynoscicola?. Parasitol Res 95:1–4
Acknowledgements
Financial support by grants from Universidad Nacional de Mar del Plata. The experiments performed during this research comply with the current laws of the Argentinian government.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Rossin, M.A., Poulin, R., Timi, J.T. et al. Causes of inter-individual variation in reproductive strategies of the parasitic nematode Graphidioides subterraneus. Parasitol Res 96, 335–339 (2005). https://doi.org/10.1007/s00436-005-1400-0
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00436-005-1400-0