
Abstract
Most pharmacokinetic studies on anthelmintic drugs have been performed on non-parasitized animals. However, it seems likely that the parasite burden could influence the deposition of such drugs. The pharmacokinetics of moxidectin administered orally and by subcutaneous injection was compared in lambs exposed to nematode infection and in parasite naive lambs. Plasma samples were analyzed for moxidectin over 40 days post-treatment. The main pharmacokinetic parameters calculated demonstrated a significant change in drug deposition in infected lambs when compared to controls. The area under the plasma concentration—time curve was decreased 54% and 46% by infection in the subcutaneous and oral groups, respectively. There was also a major decrease in the mean residence time in parasitized lambs. In parallel, the clearance of the drug was increased by infection. Thus, parasite infection dramatically influences the disposition of moxidectin in lambs. These results may contribute to determining a therapeutic strategy adapted to heavily infested animals.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Moxidectin is a broad-spectrum antiparasitic drug for the control of internal and external parasites. It is a semi-synthetic derivative of nemadectin (Asato and France 1990), a macrocyclic lactone of the milbemycin family, produced by fermentation in a culture of Streptomyces cyanogriseus. Moxidectin is largely used as an injectable or pour on antiparasitic product for beef cattle, as an oral drench and injectable formulation for sheep, and in tablet form for companion animals. Moxidectin is effective against nematodes resistant to ivermectin and it may be the only efficient drug against Haemonchus contortus in countries where anthelmintic resistance is problematic (Van Wyk et al. 1997).
Due to its lack of sugar moieties on the C13 of the macrocyclic ring, moxidectin is more lipophilic than ivermectin, the reference molecule of the avermectin family. The high affinity for fat tissue confers the molecule’s the ability to remain longer in the organism (Zulalian et al. 1994; Lanusse et al. 1997; Lifschitz et al. 2000; Craven et al. 2002). Consequently, moxidectin efficacy is very long lasting and the drug can be effective against parasites for several weeks (Kerboeuf et al. 1995). It is metabolized in the liver into C29 and C14 monohydroxy metabolites (Zulalian et al. 1994) with the involvement of the cytochrome P450 3A pathway (Dupuy et al. 2001).
The gastrointestinal parasite burden is associated with dramatic physiological changes such as intestinal dysfunction and nutritional stress leading to poor body condition. These changes may have a major impact on the plasma, tissue and gastrointestinal disposition of anthelmintic drugs and consequently on their anthelmintic efficacy. However, most pharmacokinetic studies on anthelmintics have been carried out in non-parasitized animals. The few studies performed on parasitized animals treated with oxfendazole (Hennessy et al. 1993), febantel (Landuyt et al. 1995) and fenbendazole (Marriner et al. 1985) revealed changes in the systemic pharmacokinetics of benzimidazoles or their precursors, ending with a lower animal drug impregnation. Very few data are available on the influence of parasitism on endectocide disposition. Those that are show no significant effect (McKellar et al. 1991; Echeverria et al. 2002).
Because endectocide chemotherapy is usually performed on parasitized animals, we investigated the influence of natural infection with gastrointestinal strongyles in Martinik lambs on the pharmacokinetics of orally (PO) or subcutaneously (SC) administered moxidectin.
Material and methods
Animals and treatment
The animal experimentation took place at the INRA Zootechnic Research Unit, Guadeloupe (French West Indies). We used 5-month-old black-bellied sheep (Martinik hair sheep, n=16) weighing between 18 and 26 kg. They were divided in two groups of eight animals each. Infected animals were fed by grazing on contaminated pastures. Non-infected control animals were maintained indoors and fed with hay and concentrate. Control animals were divided into two groups: PO control, administered orally with moxidectin at a dose rate of 0.2 mg/kg body weight (n=4); SC control, subcutaneously injected with moxidectin at a dose rate of 0.2 mg/kg body weight (n=4). Similarly, parasitized animals were administered PO (infected, n=4), or SC injected moxidectin, 0.2 mg/kgbody weight (infected, n=4). The animal study complied with current French laws.
Parasite status
The parasite status was followed by fecal egg counts which were recorded weekly up to the 7th week after treatment to evaluate the long-term efficiency of moxidectin. Worm examination from necropsied animals was performed to determine the species, stage and number of the main gastrointestinal nematodes. Anthelmintic efficiency of PO and SC administered moxidectin was measured by fecal egg count reduction (FECRT) according to the WAAP procedures (Wood et al. 1995).
Blood sampling
Blood samples were collected from all sheep in each group from the jugular vein into sodium heparin vacutainers at regular intervals for 28 days after moxidectin administration: at time 0 (pre-treatment), and post-treatment days 0.042, 0.084, 0.168, 0.336, 0.500, 1, 1.336, 2, 3, 4, 6, 8, 10, 12, 15, 20, 25, 30, 40. Blood was centrifuged at 1,200 g for 15 min and plasma stored at −20°C until analysis.
Drug analysis
Moxidectin was analyzed by HPLC after automated solid-phase extraction and fluorescence detection using methods adapted from Alvinerie et al. (1995). Briefly, 1 ml acetonitrile was added to 1 ml plasma, mixed and centrifuged at 2,000 g for 2 min. The supernatant was transferred into a tube which was placed in the appropriate rack of a Bench mate II apparatus (Zymarck, Hopkinton, Mass., USA). Eluates were evaporated to dryness and dissolved in 100 µl N-methylimidazole (Aldrich, Milwaukee, Wis., USA) solution in acetonitrile. The derivation was initiated with 150 µl trifluoroacetic anhydride (Aldrich) solution in acetonitrile. An aliquot (100 µl) was injected into the HPLC system with fluorescence detection (excitation 355 nm, emission 465 nm, FP-920, Jasco, Tokyo, Japan). The separation was carried out on a stainless steel analytical column Supelcosil LC18, 5 µm (150×4.6 mm internal diameter, Supelco, Bellefonte, Pa., USA). The mobile phase of acetic acid (0.2% in water)-methanol-acetonitrile (4/40/56, v/v/v for moxidectin) was pumped at a flow rate of 1.5 ml/min. Calibration graphs for moxidectin were prepared in the range of 0.5–50 ng/ml using pooled lamb drug-free plasma. The limit of quantification of the method for moxidectin analysis was 0.1 ng/ml with a coefficient of variation of 6.95%.
Pharmacokinetic and statistical analysis
The following triexponential equation was fitted to the plasma concentration versus time data using a program adapted from Multi (Yamakoa et al. 1981):
in which A1, A2 and A3 are the intercepts, C the plasma concentration at time t, Ka is the estimated first order rate constant of moxidectin absorption and α and β are the first order rate constants of moxidectin distribution and elimination, respectively. The mean residence time (MRT) was calculated by the linear trapezoidal rule without extrapolation to infinity.
Means and standard deviation (SD) were calculated for all of the parameters investigated. Comparisons of the data between groups were performed using the Kruskal-Wallis test. Differences were considered significant at P<0.05.
Results
Parasite status of lambs and moxidectin efficiency
The fecal egg count performed just before moxidectin administration confirmed that housed animals were free from parasites whereas grazing animals were infected. The geometric means of EPG before drenching were 2,363, 2,463, 2,487 for the control, SC and PO infected groups, respectively, revealing that all parasitized animals were heavily infected before the experiment. The examination of necropsied animals revealed that H. contortus was the only parasite present in the abomasum. Only a few Trichostrongylus colubriformis were identified in the intestine (Table 1). Moxidectin efficiency as estimated by FECRT was higher than 99% for the oral and injectable forms. The long-term efficiency of moxidectin in infected sheep was 3 days and 14 days for oral drench and injection, respectively.
Pharmacokinetics of moxidectin
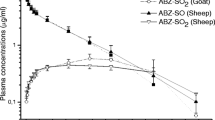
Moxidectin was detected in all animals in the first plasma sample collected 1 h after drug administration, with a maximum concentration occurring at 6–8 h post-treatment. In the non-infected group, moxidectin remained detectable at 40 days post-treatment for SC injected animals, whereas it was only detectable until 20 days in PO group infected sheep. The time—concentration curve for moxidectin in control and parasitized animals is given in Fig. 1A (SC) and in Fig. 1B (PO). The area under the curve (AUC) and the Cmax of moxidectin in the plasma of lambs was three- and fivefold higher when moxidectin was administered SC compared to PO administration. In addition, a lower clearance was measured after SC injection compared to PO drenching (Table 2).

Time—concentration curve of moxidectin in the plasma of lambs after: A subcutaneous injection, or B oral administration. Control parasite-free (open symbols) and infected (black symbols) lambs were given moxidectin at a dose rate of 0.2 mg/kg body weight. The drug concentration is shown for the plasma over a period of 20 days. Results are expressed as ng/ml and are the mean±SE of four animals
Infection induced clear modifications in moxidectin disposition reflected by major changes in the shape of the time—concentration curve (Fig. 1A, B) and in the main pharmacokinetic parameters (Table 2). In PO infected lambs, the moxidectin AUC was decreased twofold (not significant, Fig. 2A), with a strong shortening of the MRT (Fig. 2B, P<0.001). These changes were associated with an increase in clearance (P<0.05) and a 2.5-fold decrease in terminal half-life of elimination, t1/2β (Table 2).

Area under the time-concentration curve (AUC) and B mean residence time (MRT) of moxidectin after oral (PO) and subcutaneous (SC) administration to lambs. AUC and MRT were calculated from the time—concentration curve of moxidectin in sheep plasma. Data are the mean±SE of four animals
Similarly, in the SC group the AUC was diminished twofold (Fig. 2A, P<0.01), while the MRT of 10.84±1.56 days in controls was reduced to 3.66±1.51 days in parasitized animals (Fig. 2B, P<0.001). In parallel, the infection induced an increase in the clearance of moxidectin associated with a decrease of t1/2β (Table 2, P<0.02).
Discussion
It is well documented for anthelmintic endectocides that the quantity of drug in the plasma is the major determinant of the expression of its efficacy against parasites. The drug impregnation of treated animals is dependent on the dose, the route of administration as well as the species of host and parasite and the physiological status of the host. Among macrocyclic lactones, moxidectin is different, because it has a longer residence time in the organism inducing a longer effective time of activity. However, changes in physiological conditions such as fasting or lactation influence drug disposition (Alvinerie et al. 2000). These changes may modulate the therapeutic efficacy of macrocyclic lactones.
Because parasitism can be associated with a dramatic modification of host physiological function, we speculate that nematode infection can influence the behavior of moxidectin in the host organism. In order to verify this point, a group of sheep were naturally infected while grazing. The parasite status evaluation revealed that H. contortus was the main parasite found in necropsied animals. In parallel, non-infected sheep were maintained indoors and used as the control group.
Our results confirm the long residence time of the lipophilic moxidectin in control sheep. The parental molecule was detected as long as 40 days after moxidectin treatment, no matter whether the route of administration was SC or PO. As expected, the value of the AUC was higher after SC injection compared to PO administration.
Infection induced significant changes in moxidectin disposition, with an increase in drug clearance. In parallel, the t1/2β parameter, indicating the half-time of elimination of the drug in the terminal phase of the time—concentration curve, was also decreased. These changes were combined with significant decreases of the AUCs in parasitized animals. The lower drug concentration measured in control lambs after oral drench when compared to SC was also found in infected animals. Altogether, these results revealed that infected sheep were exposed to less drug compared to control animals. These effects were independent of the route of administration as they were observed in both the PO and SC groups.
Anthelmintic disposition may be influenced by parasitism. The AUC of orally administered oxfendazole was reduced in goats and sheep infected with H. contortus and Teladorsagia circumcincta (Hennessy et al. 1993). Similarly, dramatic changes were observed in fenbendazole pharmacokinetics in sheep heavily infected Oestortagia circumcincta (Marriner et al. 1985). However, according to the major differences existing between benzimidazoles and endectocides in terms of biotransformation, residence time, solubility and pH-dependence, the changes observed during parasitism for benzimidazole do not predict the effect of parasitism on endectocide disposition.
Only a few works have intended, albeit unsuccessfully, to demonstrate an effect of parasitism on endectocide disposition in sheep. Intestinal nematode infection with Nematodirus battus does not appear to significantly change the plasma kinetics of ivermectin administered by PO or SC administration in lambs. The disposition of levamisole and netobimin (a precursor of albendazole) was also unchanged when administered to infected sheep (McKellar et al. 1991). Although a mange infection in sheep tended to accelerate the absorption of ivermectin and to reduce the AUC, the changes were mild and not significant (Echeverria et al. 2002). However, it is probable that the changes occurring in drug disposition are closely dependent on the parasite burden and the extent of physiological alteration induced by the infection. In the study of McKellar et al. (1991), the parasite burden was low and the consequent physiological alterations probably too mild to have notable effects on drug disposition.
The low concentration of drug observed during infection in our experiment may be the result of a higher elimination rate associated with an increase in biotransformation of the drug. Indeed, infection with H. contortus induced antipyrine clearance in lambs (Kawalek and Fetterer 1990), consistent with an increase in cytochrome P450 activity. Similarly, Fasciola hepatica infection influenced biotransformation enzyme activities in sheep (Galtier et al. 1991; Benchaoui and McKellar 1993; Biro-Sauveur et al. 1995). The influence of parasitism on biotransformation enzymes may be a determinant in parental drug level in the treated host.
Besides drug biotransformation, host pathophysiological changes occurring during gastrointestinal parasitism may also influence the fate of drugs. H. contortus infection was associated with anemia and T. circumcincta infection with the loss of protein as albumin (Steel et al. 1980). Albumin is the main plasma binding protein for benzimidazole, but it is not a major carrier of endectocides that bind with more affinity to plasma lipoproteins (Lespine et al. 2003). We cannot exclude the possibility that some changes in lipid turnover may occur in parasitized lambs which account for the low moxidectin levels. The drop of abomasal pH occurring during infestation with O. circumcincta certainly interferes with the absorption rate of orally administered fenbendazole (Marriner et al. 1985), but must have limited consequences on the solubility and absorption rate of the non-ionic moxidectin. Furthermore, in our experiment the changes in drug deposition observed in parasitized animals occurred not only in PO treated animals, but also after SC administration, with a minor contribution from intestinal absorption. The increase in abomasal fluid flow rate after H. contortus and T. circumcincta infection may also account for the change in drug distribution, but once again this effect should be more relevant for benzimidazole given PO, for which the abomasum is the major absorption site.
Parasitism is also associated with an increase in intestinal secretions and gut motility. Because large amounts of unchanged moxidectin are excreted in the feces via bile (Zulalian et al. 1994; Lifschitz et al. 1999; Hennessy et al. 2000) and intestinal secretions (Laffont et al. 2002), these changes probably contribute to the increase in moxidectin clearance observed in infected lambs, whatever the route of administration. In agreement with the significant involvement of bile and intestinal secretions of endectocides, loperamide, an opioid derivative that reduced gastrointestinal secretions and motility, has been shown to induce a clearance reduction and AUC increase in moxidectin in cattle (Lifschitz et al. 2002).
In addition, heavy parasite infections are associated with nutritional stress, influencing the body fat composition. Considering the strong influence of fat on moxidectin disposition (Zulalian et al. 1994; Craven et al. 2002), an extensive loss of fat reserves may contribute to decrease the drug level and its activity time in infected animals.
In conclusion, we have demonstrated that natural gastrointestinal parasitism influences moxidectin disposition in sheep after both the PO and SC administration of moxidectin. The short residence time of the drug observed in infected animals may contribute to reducing the long-term efficiency of moxidectin. Consequently, reinfection may occur more rapidly when the drug is given to heavily infected animals, especially when it is administered PO. These results may contribute to defining better therapeutic strategies adapted to heavily infected animals.
References
Alvinerie M, Sutra J F, Badri M, Galtier P (1995) Determination of moxidectin in plasma by high-performance liquid chromatography with automated solid-phase extraction and fluorescence detection. J Chromatogr B Biomed Appl 674:119–124
Alvinerie M, Cabezas I, Rubilar L, Perez R (2000) enhanced plasma availability of moxidectin in fasted horses. J Equine Vet Sci 20: 575–578
Asato G, France D (1990) The American Cynamid Co. U S patent 4916, pp 154–158
Benchaoui HA, McKellar QA (1993) Effect of early treatment with rafoxanide on antipyrine clearance in sheep infected with Fasciola hepatica. Xenobiotica 23:439–448
Biro-Sauveur B, Eeckhoutte C, Baeza E, Boulard C, Galtier P (1995) Comparison of hepatic and extrahepatic drug-metabolizing enzyme activities in rats given single or multiple challenge infections with Fasciola hepatica. Int J Parasitol 25:1193–1200
Craven J, Bjorn H, Hennessy DR, Friis C (2002) The effects of body composition on the pharmacokinetics of subcutaneously injected ivermectin and moxidectin in pigs. J Vet Pharmacol Ther 25:227–232
Dupuy J, Escudero E, Eeckhoutte C, Sutra JF, Galtier P, Alvinerie M (2001) In vitro metabolism of 14C-moxidectin by hepatic microsomes from various species. Vet Res Commun 25:345–354
Echeverria J, Mestorino N, Errecalde JO (2002) Comparative pharmacokinetics of ivermectin after its subcutaneous administration in healthy sheep and sheep infected with mange. J Vet Pharmacol Ther 25:159–160
Galtier P, Alvinerie M, Plusquellec Y, Tufenkji AE, Houin G (1991) Decrease in albendazole sulphonation during experimental fascioliasis in sheep. Xenobiotica 21:917–924
Hennessy DR, Sangster NC, Steel JW, Collins GH (1993) Comparative kinetic disposition of oxfendazole in sheep and goats before and during infection with Haemonchus contortus and Trichostrongylus colubriformis. J Vet Pharmacol Ther 16:245–253
Hennessy DR, Page SW, Gottschall D (2000) The behaviour of doramectin in the gastrointestinal tract, its secretion in bile and pharmacokinetic disposition in the peripheral circulation after oral and intravenous administration to sheep. J Vet Pharmacol Ther 23:203–213
Kawalek JC, Fetterer RH (1990) Effect of Haemonchus contortus infection on the clearance of antipyrine, sulfobromophthalein, chloramphenicol, and sulfathiazole in lambs. Am J Vet Res 51:2044–2049
Kerboeuf D, Hubert J, Cardinaud B, Blond-Riou F (1995) The persistence of the efficacy of injectable or oral moxidectin against Teladorsagia, Haemonchus and Trichostrongylus species in experimentally infected sheep. Vet Rec 137:399–401
Laffont CM, Toutain PL, Alvinerie M, Bousquet-Melou A (2002) Intestinal secretion is a major route for parent ivermectin elimination in the rat. Drug Metab Dispos 30:626–630
Landuyt J, Debackere M, Vercruysse J, McKellar Q (1995) The influence of a heavy infection with sensitive and resistant strains of Ostertagia circumcincta and with Trichostrongylus colubriformis on the pharmacokinetics of febantel in lambs. J Vet Pharmacol Ther 18:180–186
Lanusse C, Lifschitz A, Virkel G, Alvarez L, Sanchez S, Sutra J F, Galtier P, Alvinerie M (1997) Comparative plasma disposition kinetics of ivermectin, moxidectin and doramectin in cattle. J Vet Pharmacol Ther 20:91–99
Lespine A, Bree F, Bassissi F, Sutra JF, Tillement JP, Alvinerie M (2003) Binding of ivermectin and moxidectin to the plasma lipoproteins. 9th International EAVPT Congress, 14–18 July, Lisbon
Lifschitz A, Virkel G, Imperiale F, Sutra JF, Galtier P, Lanusse C, Alvinerie M (1999) Moxidectin in cattle: correlation between plasma and target tissues disposition. J Vet Pharmacol Ther 22:266–273
Lifschitz A, Imperiale F, Virkel G, Munoz Cobenas M, Scherling N, DeLay R, Lanusse C (2000) Depletion of moxidectin tissue residues in sheep. J Agric Food Chem 48:6011-6015
Lifschitz A, Virkel G, Sallovitz J, Imperiale F, Pis A, Lanusse C (2002) Loperamide-induced enhancement of moxidectin availability in cattle. J Vet Pharmacol Ther 25:111–120
Marriner SE, Evans ES, Bogan JA (1985) Effect of parasitism with Ostertagia circumcincta on pharmacokinetics of fenbendazole in sheep. Vet Parasitol 17:239–249
McKellar QA, Jackson F, Coop RL, Jackson E, Scott E (1991) Effect of parasitism with Nematodirus battus on the pharmacokinetics of levamisole, ivermectin and netobimin. Vet Parasitol 39:123–136
Steel J, Symons LEA, Jones WO (1980) Effect of level of larval intake on the productivity and physiological and metabolic responses of lambs infected with Trichostrongylus colubriformis. Aust J Agric 31:821–838
Van Wyk JA, Malan FS, Randles JL (1997) How long before resistance makes it impossible to control some field strains of Haemonchus contortus in South Africa with any of the modern anthelmintics? Vet Parasitol 70:111–122
Wood IB, Bairden K, Duncan JL, Kassai T, Malone JB, Pankavich JA, Reinecke RK, Slocombe O, Taylor SM, Vercruysse J (1995) World Association for the Advancement of Veterinary Parasitology (W.A.A.V.P) second edition of guidelines for evaluating the efficacy of anthelmintics in ruminants (bovine, ovine, caprine). Vet Parasitol 58:181–213
Yamakoa TK, Nokaguana K, Unot T (1981) A pharmacokinetic analysis program (multi) for microcomputer. J Pharmacobiol Dyn 4:879–885
Zulalian J, Stout S, DaCunha A, Garces T, Miller P (1994) Absorption, tissue distribution, metabolism, and excretion of moxidectin in cattle. J Agric Food Chem 42:381–387
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Lespine, A., Sutra, JF., Dupuy, J. et al. The influence of parasitism on the pharmacokinetics of moxidectin in lambs. Parasitol Res 93, 121–126 (2004). https://doi.org/10.1007/s00436-004-1084-x
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00436-004-1084-x