
Abstract
The current study provides a generalizable account of the anatomo-functional associations as well as the connectivity of representational codes underlying numerical processing as suggested by the triple code model (TCM) of numerical cognition. By evaluating the neural networks subserving numerical cognition in two specific and substantially different numerical tasks with regard to both grey matter localizations as well as white matter tracts we (1) considered the possibility of additional memory-related cortex areas crucial for arithmetic fact retrieval (e.g., the hippocampus); (2) specified the functional involvement of prefrontal areas in number magnitude processing, and, finally; (3) identified the connections between these anatomo-functional instantiations of the representations involved in number magnitude processing and arithmetic fact retrieval employing probabilistic fiber tracking. The resulting amendments to the TCM are summarized in a schematic update, and ideas concerning the possible functional interplay between number magnitude processing and arithmetic fact retrieval are discussed.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Over the last 25 years functional brain imaging has substantially contributed to the development of the neuro-functional triple code model (TCM) of numerical cognition (TCM, e.g., Dehaene and Cohen 1995, 1997; Dehaene et al. 2003). So far, research on numerical cognition in general and on the TCM in particular focused on the identification of activated grey matter sites, with only very few studies investigating its neuro-structural white matter connections (Krueger et al. 2011; Uddin et al. 2010; Klein et al. 2013). Therefore, the aim of the present study was to complement the TCM of numerical cognition (Dehaene et al. 2003; Arsalidou and Taylor 2011) by identifying neuro-structural connections to/from cortical areas related to arithmetic fact retrieval and numerical magnitude processing.
The TCM of numerical cognition
In the TCM (Dehaene and Cohen 1995, 1997; Dehaene et al. 2003) three different representational codes are proposed for the processing of numerical information: first, a visual number form representation is involved in recognizing (strings of) Arabic digits and located in bilateral fusiform and lingual regions. Second, a bi-hemispheric number magnitude representation subserves the processing of numerical quantity information (e.g., involved in number comparison, etc.) and is associated with brain areas around the bilateral intraparietal sulcus (IPS). Third, a verbal representation of numbers is proposed to be activated in language-mediated operations like number naming and counting. It is assumed that arithmetic facts (e.g., additions <10; multiplication table facts) are also stored in a verbal code in long-term memory (e.g., Delazer et al. 2003; Grabner et al. 2009). The verbal representation is associated with left perisylvian language areas such as middle (MTG) and superior temporal gyrus (STG), supramarginal gyrus (SMG), Broca’s area, and the angular gyrus (AG).
In sum, the TCM suggests that different aspects of numerical information are processed in different codes within distinct cerebral regions of the human brain with strong bi-directional links between all three codes. One of the most important postulates of the TCM is the general distinction between numerical magnitude manipulation on the one hand and verbally mediated fact retrieval on the other. For complex tasks Dehaene and Cohen (1995) suggested that the representations of the TCM may be further supplemented by processes associated with prefrontal cortex (PFC) areas. Activation within these areas is not necessarily specific for number processing (cf. Pesenti et al. 2000). Instead, it may rather reflect more general cognitive processes such as attention, working memory, and cognitive control (e.g., Ashcraft and Kirk 2001; Imbo et al. 2007a, b; Miller 2000). Taken together, the fronto-parietal network of number processing (Dehaene et al. 2003) is a case of multi-modular processing involving different number-specific representations but also more general cognitive resources.
Recruitment of different representations in a given task
Examination of the different representational codes and their neural correlates including their connectivity requires the employment of specific tasks. The visual number form (VNF) is commonly involved whenever Arabic numbers are visually presented. However, because we aimed at specifying neuro-structural connectivity within the TCM, we needed to use tasks that recruit, but also allow for differentiating magnitude-related processing from fact retrieval. Therefore, we re-analyzed data from a number bisection task (NBT; Wood et al. 2008) and a mental exact/approximate addition task (Klein et al. 2009).
In the NBT participants have to decide whether or not the central number of a number triplet is also the arithmetical middle of the interval or not (e.g., 10_14_18 vs. 10_13_18, respectively). Wood et al. (2008) found increased IPS activation associated with magnitude manipulations for incorrectly bisected triplets with smaller distances from the central number to the correct mean (e.g., 10_13_18), correctly bisected triplets not part of a multiplication table (e.g., 21_23_25), and triplets crossing a decade boundary (e.g., 24_29_34). On the other hand, with increasing distance to the correct mean of incorrectly bisected triplets, when a correctly bisected triplet was part of a multiplication table, and when triplets did not cross a decade boundary, less IPS activation was observed. Additionally, the latter was accompanied by increasing activation in the left AG, probably indicating fact retrieval.
In the exact/approximate addition study by Klein et al. (2009) participants had to choose which one of two response alternatives was either the correct result (e.g., 25 + 31 = 56 vs. 51) or closer to the correct result (e.g., 25 + 31 = 54 vs. 51). The authors observed that large target distance (i.e., distance between correct result and target), small distractor distance (i.e., distance between target and distractor) and the need for a carry operation were associated with increased activation in the (p)IPS subserving magnitude-related processing. On the other hand, small target distance, large distractor distance and the absence of a carry were associated with increased activation in the left AG subserving fact retrieval.
In sum, the NBT and the exact/approximate addition task seem well suited to identify the influence of different numerical representations at least with respect to grey matter areas.
Involvement of prefrontal cortex and memory-related structures
In the TCM the involvement of PFC in numerical cognition is only vaguely specified, both in terms of function as well as in terms of more circumscribed cortex areas. By re-analyzing the numerical tasks described above—chosen because they recruit different representational codes—we aimed at specifying which parts of PFC are involved in which numerical operation. Beyond specifying frontal networks called upon in one numerical task or the other, we aimed at identifying frontal activation patterns which would generalize over numerical tasks—possibly indicating procedural similarity across tasks.
Moreover, the TCM states that participants retrieve arithmetic facts from long-term memory. However, the role of typical cortical regions subserving processes involving long-term memory such as hippocampus, parahippocampal, perirhinal or entorhinal cortex (Baddeley 1996; Montaldi and Mayes 2010) has not been specified explicitly. Instead, it is suggested that the left angular gyrus and, probably, left-hemispheric language areas may support the retrieval of arithmetic facts. In our view, it is reasonable to assume that structures associated with retrieval from long-term memory may also be involved in arithmetic fact retrieval. Therefore, we also considered areas commonly encountered in retrieval processes from long-term semantic memory and found activated in our re-analyses.
Objectives
Starting from the cortical structures which play a pivotal role in the TCM of numerical cognition this study aims at investigating their structural connectivity. The areas of interest comprise (1) the IPS (subserving number magnitude processing); (2) left perisylvian language areas (MTG, STG, SMG, Broca) and left AG (associated with arithmetic fact retrieval); and (3) fusiform gyrus (subserving visual number form processing). We also investigated the structural connectivity of areas, which have recently been proposed either in an update of the TCM, namely (4) prefrontal areas or in recent studies such as (5) the hippocampus, which appear to be underspecified with respect to their function in the current version of the TCM. To pursue these aims, we relied on probabilistic fiber tracking, employing seed-points motivated by both meta-analytic (TCM, Arsalidou and Taylor 2011; Dehaene et al. 2003) as well as empirical considerations from earlier studies (cf. Klein et al. 2013 for a similar procedure). After re-analyzing the functional data from the previous studies, DTI data were collected in a separate set of participants and fiber tracking was applied between the seeds identified in the re-analyses. In particular, we re-analyzed the impact of specific parametric predictors used in the NBT of Wood et al. (2008, i.e., distance to the arithmetic mean of the triplet, multiplicativity, and decade crossing) and the exact/approximate addition task of Klein et al. (2009, i.e., target distance, decade distance, and the need for a carry operation) by evaluating the joint impact of the respective parametric predictors in a conjunction analysis. To evaluate commonalities and differences in the architecture of the pathways connecting the fronto-parietal network of numerical cognition in the different tasks (i.e., NBT and exact/approximate addition) grey matter activation peaks were defined as seed and target regions for separate probabilistic trackings for both numerical tasks. Our specific hypotheses were as follows:
-
(a)
Since both, the exact/approximate addition task and the NBT employ visual presentation of the stimuli, the visual number form area (VNF) should be involved in both tasks. Because the VNF subserves a numerical input representation, it should be (directly) connected to the semantic numerical representation in the IPS.
-
(b)
The numerical magnitude representation in the bilateral IPS should be part of the magnitude-processing network in both tasks, because number magnitude is the critical information. For the NBT we expected increasing IPS activation with decreasing distance to the arithmetic mean of the triplet, inclusion of a decade crossing, and for non-multiplicative triplets. For the exact/approximate addition task increasing target distance, decreasing distractor distance and the need for a carry operation should be associated with an increase in activation in IPS as well. Because both tasks access numerical magnitude information, we expected activation in overlapping areas as well as similar dorsal and ventral fiber pathways connecting intraparietal with frontal cortex areas (cf. Klein et al. 2013).
-
(c)
We hypothesized activation in areas associated with arithmetic fact retrieval for both tasks. These are left-hemispheric language areas (Dehaene and Cohen 1997; Dehaene et al. 2003) such as SMG, STG as well as the left AG. In the NBT, arithmetic fact retrieval should be reflected by increasing activation in at least the left AG associated with increasing distance to the arithmetic mean of the triplet, no decade crossing within, and multiplicativity of a triplet. In the exact/approximate addition task, at least AG activation should increase with decreasing target distance, increasing distractor distance as well as for no carry problems. Additionally, we expected to identify neural activation associated with retrieval from long-term memory (e.g., in the hippocampus) for the case of arithmetic fact retrieval. Again, we hypothesized that for both tasks activation in overlapping areas connected via similar ventral and dorsal pathways should be observed.
-
(d)
With respect to PFC, we expected activation in Area 45 and Brodmann area (BA) 47 indicating the application of rules, which should be connected via similar pathways in both tasks.
fMRI methods
We re-analyzed the fMRI data of Klein et al. (2009) and Wood et al. (2008) to identify adequate seed regions for probabilistic fiber tracking. The resulting seed regions were then applied to independent DTI data from a sample comparable to those of both fMRI experiments with respect to age, handedness, and general level of education. No participant reported a neurological or psychiatric history or was taking psychoactive medication. All three participant groups gave their written consent and were scanned with the approval of the respective local ethics committee.
Number bisection task (Wood et al. 2008)
Participants
Twenty male right-handed volunteers (M = 24.2 years, range 21–30 years).
Stimuli and design
Participants were presented with number triplets (e.g., 10_13_18; numbers always in ascending order from left to right) and had to decide whether or not the central number was also the arithmetical middle of the interval. 360 number triplets in the range from 2 to 98 were presented in a 2 × 2 within-participant design for both correctly bisected (e.g., 23_26_29) and incorrectly bisected triplets (e.g., 23_25_29). For the former, bisection range (small: 4–8, e.g., 23_26_29 vs. large 12–18, e.g., 22_30_38) and multiplicativity (e.g., 21_24_27 vs. 22_25_28) were varied. For the latter, incorrectly bisected triplets, bisection possibility (possible: e.g., 21_24_29, vs. impossible: e.g., 21_24_28) and distance of the central number to the correct mean of the interval (large: 2–8, e.g., 6_7_18 vs. small: 0.5–1.5, e.g., 6_11_18) were manipulated. Decade crossing (e.g., 23_25_29 vs. 25_27_31) was matched between the respective stimulus groups.
Exact/approximate addition task (Klein et al. 2009)
Participants
Eighteen male right-handed volunteers (M = 23.6 years, range 19–31 years).
Stimuli and design
In a choice-reaction paradigm, 192 two-digit addition problems were presented with a pair of solution probes. Participants had to indicate the solution probe, which was either identical (50 % of the trials, e.g., 25 + 31 = 56 or 51) or closest to the correct result (50 % of the trials, e.g., 25 + 31 = 54 or 51), as quickly and as accurately as possible. In a 2 × 2 × 2 within-subjects design the factors distractor distance (small, i.e., 4–9 vs. large, i.e., 14–19 distance between target and distractor), target identity (identical vs. non-identical target) and carry operation (carry needed yes/no) were varied.
Re-analysis
Anatomical scans of participants were submitted to grey/white matter segmentation and then normalized and averaged in SPM8 (http://www.fil.ion.ucl.ac.uk/spm). The fMRI time-series were corrected for movement artifacts in SPM8. Images were motion corrected and realigned to each participant’s first image. Data were normalized into standard stereotaxic MNI (Montreal Neurological Institute) coordinates space. Images were resampled every 3.5 mm using 4th degree spline interpolation and smoothed with a 7.5-mm FWHM Gaussian kernel. The data were high-pass filtered (128 s) to remove low-frequency signal drifts and corrected for autocorrelation assuming an AR (1) process. Brain activity was convolved over all experimental trials with the canonical hemodynamic response function (HRF) and its derivative. Onsets of incorrectly answered trials were entered separately as a condition of no interest into the model. For the anatomical localization of effects the SPM Anatomy Toolbox was used (Eickhoff et al. 2005), available for all published cytoarchitectonic maps from http://www.fz-juelich.de/ime/spm_anatomy_toolbox. In the following, all regions we refer to are in cytoarchitectonic nomenclature except for BA 47 (for this region there are no maps available yet). Please note that areas 7A, 7P, 7PC, hIP1, hIP2, and hIP3 represent a parcellation of Brodmann area (BA) 7 with 7A, 7P, and 7PC defining subdivisions of the SPL (Scheperjans et al. 2008), while hIP1, hIP2, and hIP3 specify parts of the IPS (Uddin et al. 2010). Furthermore, PGa and PGp are parcellations of the AG, while PFt, PF, and PFm are three of five subdivisions of the SMG (Caspers et al. 2006).
Number bisection
A conjunction analysis of the impact of the three predictors distance to the arithmetic middle, decade crossing, and multiplicativity was run. A significant conjunction of activation with increasing distance to the arithmetic middle, inclusion of a decade crossing, and items not being part of a multiplication table should indicate areas subserving magnitude-related processing, whereas a significant conjunction of the reverse predictor activations (decreasing distance to the arithmetic middle, no decade crossing, multiplicative items) should be associated with activation in areas subserving verbally mediated processes of arithmetic fact retrieval.
Exact/approximate addition
For the re-analysis, predictors representing target distance (ranging 1–3), distractor distance (ranging 4–19), and carry-over (no = −1, yes = +1) were estimated in a parametric analysis. Again, we conducted a conjunction analysis on the impact of the predictors target distance, distractor distance, and carry operation. A significant conjunction of activation with increasing target distance, decreasing distractor distance and a carry operation should indicate areas subserving number magnitude processing, whereas a significant conjunction of activation with decreasing target distance, increasing distractor distance and no carry operation should be associated with activation in areas subserving verbally mediated processes of arithmetic fact retrieval.
fMRI results
Re-analysis of functional MRI data
Number bisection task (Wood et al. 2008)
The conjunction analysis using increasing distance to the arithmetic mean, decade crossing, and non-multiplicative triplets revealed significant activation in IPS (hIP3) and bilateral pIPS (7A and 7P; see Figure S1 A; Table 1). Further clusters of activated voxels were observed in the left IFG in Area 45, Area 44, and BA 47 as well as in the right IFG (Area 44). Moreover, activation was observed in left SMA (Area 6) and bilateral middle frontal gyri, possibly situating bilateral frontal eye fields (FEF). Additionally, activation was found in the bilateral fusiform gyrus, indicating activation of the VNF area, and visual cortices, bilaterally, as well in the basal ganglia including the thalamus, bilaterally, the left putamen and the right caudate nucleus.
The conjunction analysis using decreasing target distance, increasing distractor distance and no-carries revealed decreasing fMRI signal change in left AG (PGp). Further clusters of activated voxels were observed in left retro-splenial (RC) cortex, bilateral SMG (PF/PFm), bilateral hippocampus, left anterior and posterior MTG, and left ventromedial PFC (VMPFC, see Figure S2 A).
Exact/approximate addition task (Klein et al. 2009)
The conjunction analysis using increasing target distance, decreasing distractor distance, and carry operation revealed significant modulation of the fMRI signal in bilateral IPS (hIP2) and (pIPS, 7A; see Figure S3 A; Table 2). Further activated clusters were observed in left IFG in Area 45 and BA 47 as well as in right IFG (Area 45). Moreover, activation was observed in bilateral supplementary motor areas (SMA, Area 6) and bilateral middle frontal gyri, again most likely corresponding to the bilateral FEF. Additionally, activation was observed in left fusiform gyrus, possibly indicating activation of the VNF area and in visual cortices, bilaterally.
The conjunction analysis of decreasing target distance, increasing distractor distance, and no-carries revealed decreasing fMRI signal in left AG (PGp), bilateral RC and left MTG. Further clusters of activated voxels were observed in right SMG (PFt), bilateral STG, right MTG, left insula, left hippocampus as well as left VMPFC (see Figure S4 A).
DTI methods
Diffusion imaging data
Participants
The DTI data were collected from a sample of 30 healthy volunteers (12 females, mean age 28 years, range 23–42 years; 29 right-handed participantsFootnote 1).
Data acquisition
The MRI data from all 30 subjects were acquired on a 3T Siemens TIM Trio scanner (Siemens, Erlangen, Germany). For DTI a total of 70 scans with 69 slices was acquired using a diffusion-sensitive spin-echo EPI sequence with CSF-suppression [61 diffusion encoding gradient directions (b factor = 1,000 s/mm2) and 9 scans without diffusion weighting (b value = 0 s/mm2), voxel size = 2 × 2 × 2 mm3, matrix size = 104 × 104, TR = 11.8 s, TE = 96 ms, TI = 2.3 s]. During reconstruction, scans were corrected for motion and distortion artifacts based on a reference measurement (Zaitsev et al. 2006). For spatial processing of the DTI data an additional high-resolution T1 anatomical scan was obtained (160 slices, voxel size = 1 × 1 × 1 mm3, TR = 2.2 s, TE = 2.6 ms, FOV = 240 × 240 × 160 mm3).
Definition and analysis of seed regions
Seed regions for probabilistic fiber tracking were extracted from the t-maps of the parametric fMRI analyses of the two fMRI tasks. Peak voxels in the major activation clusters of both studies were identified at an FWE cluster-threshold corrected p cluster-corr < 0.05 with a cluster size of k = 15 voxels. In a next step, each of the coordinates identified in the re-analyses were transferred to the native space of each participant’s DTI data, using the inverse normalization parameters obtained from the segmentation procedure of the T1 anatomical scan and enlarged to a sphere with radius 4 mm, each containing 33 seed voxels. These spheres were taken as seed regions for the probabilistic fiber tracking procedure.
Probabilistic diffusion tensor-based fiber tracking
DTI data were analyzed using the method of pathway extraction developed by Kreher et al. (2008; see also Saur et al. 2008, 2010) implemented in the Matlab-based DTI and Fiber Toolbox (http://www.uniklinik-freiburg.de/mr/live/arbeitsgruppen/diffusion/fibertools_en.html). The diffusion tensor (DT) was calculated (Basser et al. 1994) from the movement and distortion-corrected diffusion-weighted imaging dataset. Subsequently, probabilistic maps were calculated for each seed region by Monte Carlo simulation of random walks similar to the probabilistic index of connectivity (PICo) method (Parker et al. 2003). The tracking procedure differed from the classical PICo method by empirically extracting the orientation density function from the diffusion tensor and by preserving the main traversing direction (in relation to the first eigenvector) of each propagated trajectory during the random walk (see Kreher et al. 2008 for details). In the probabilistic maps, the visiting frequency of a voxel reflects the degree of connectivity to the seed region. The number of random walks was set to 105 and the maximum fiber length to 150 voxels. The tracking area was restricted to a white matter mask to avoid tracking across anatomical borders. To ensure contact of the cortical seed regions with white matter, a rim of grey matter was included in the mask.
Region-to-region anatomical connectivity between two seed regions was computed by combining two probability maps of interest (Kreher et al. 2008). At the computational level, this combination implies a multiplication, which takes the main traversing direction of the random walk into account. Walks starting from two seed regions may either face in opposing directions (connecting fibers) or they may merge and point in the same direction (merging fibers). Within the pathway connecting both seeds, the proportion of connecting fibers should exceed the proportion of merging fibers. Using the directional information (obtained by extended PICo) during the multiplication procedure, merging fibers are suppressed, while connecting fibers are preserved (Kreher et al. 2008). This method enables the extraction of the most probable direct pathway between two seed regions without using a priori knowledge about the presumed course. The resulting values of the combined maps represent voxel-wise estimation of the probability index that a voxel is part of the connecting fiber bundle of interest (in short: probability index forming part of the bundle of interest, PIBI).
Post-processing of probability maps
Using SPM8 (http://www.fil.ion.ucl.ac.uk/spm) the combined maps were scaled to the range from 0 to 1, and spatially normalized into standard MNI space. To remove random artifacts, maps were thresholded using a PIBI value of 0.0148. This value was generated empirically from the distribution observed in a large collection of pre-processed combined probability maps (see Saur et al. 2008). The resulting maps of each region-to-region connection were then summed across subjects (Newton et al. 2006). Within these across-subject maps, the voxel intensity ranged from 1 to 30 indicating spatial overlap between subjects. Here, all maps are displayed with an arbitrary threshold of 10 to eliminate noise. 3D volume renderings of pathways were visualized with an application from the Freiburg Brain Imaging Center based on the free software OpenDX (http://www.opendx.org). Resulting tracts were identified based on atlases on connectivity of the human cortex (Bürgel et al. 2006; Catani and Thiebaut de Schotten 2012; Oishi et al. 2011) as well as the macaque cortex (Schmahmann and Pandya 2006).
DTI results
Seed regions
To define meaningful seed regions, we compared the empirical evidence from the re-analyses of the fMRI data with propositions of the TCM and its latest amendments (Arsalidou and Taylor 2011; Dehaene and Cohen 1995; Dehaene et al. 2003). According to the TCM the identification of Arabic digits recruits the VNF (situated in the fusiform gyrus). Since this area was found active in the re-analyses of both tasks, it was incorporated as a seed region.
For number magnitude processing, the TCM proposes bilateral activation of IPS, PSPL/pIPS, and the fusiform gyrus, all of which were identified in the re-analyses of Wood et al. (2008) and Klein et al. (2009, see Tables 1, 2)—and thus included as seed regions for fiber tracking.
For fact retrieval, the TCM proposes activation of the left AG and of left-hemispheric perisylvian language areas, which we also found in the re-analyses of both tasks and thus incorporated as seed regions: for the addition task STG, MTG, and insula; for the NBT the anterior and posterior MTG and the SMG.
Furthermore, frontal areas, which are not specified in more detail in the TCM, are recruited in complex numerical tasks. In the re-analyses we were able to identify the following frontal regions, which were included as seed regions: triangular part of the inferior frontal gyrus (IFGtri, Area 45), BA 47, supplementary motor area (SMA), and FEF (the latter two were already recommended by Arsalidou and Taylor to be included in the TCM 2011). The re-analysis of the NBT data also revealed significant activation in Area 44 (opercular part of IFG, IFGoper). Thus, we included these regions in the fiber tracking for the NBT task. The coordinates reported by Arsalidou and Taylor (2011) for magnitude processing fell within the spheres we defined as seed regions for our trackings for both parietal as well as prefrontal coordinates. Thus, our trackings for the magnitude-related processing network (see below) consider the results of the meta-analysis by Arsalidou and Taylor (2011). Moreover, our data complement the findings of Arsalidou and Taylor (2011), because our prefrontal seed-points reflect a specification of the involvement of cortical areas summarized as DLPFC by these authors.
Based on hypotheses concerning additional processes involved in the two numerical tasks, which have not yet been specified anatomically in the TCM (see above), we added further seed regions. In particular, we included areas, which were repeatedly associated with the recognition of familiar objects and procedures, such as retro-splenial cortex (for a review, see Vann et al. 2009) and an area known to be involved in retrieval from long-term memory: the hippocampus (see Montaldi and Mayes 2010; Rugg and Yonelinas 2003 for reviews). For a detailed description of the coordinates of seed-points and trackings, see Table 3.
Finally, because (1) the main regions supposed to be involved in the verbal representation of numbers are restricted to the left hemisphere and (2) the main regions assumed to contribute to magnitude processing are supposed to be situated bi-hemispherically (which is also true for the visual number form area, when Arabic digits are involved), we concentrated our analyses on the left hemisphere only.
Connectivity analysis
Number bisection task
The overlay of magnitude and fact retrieval-related processing suggests two largely separate networks. These networks not only differ in localization of the seed regions (Fig. 1a; Table 1) but also in the connections between them (Fig. 1b–d: 3D presentation and axial slices; see also Figure S5 for more details and different views). The connections involve dorsal and ventral pathways, only overlapping to a very limited degree in the vicinity of the AG and the extreme and/or external capsule system (EC/EmC) system. Importantly, both networks—even though seemingly largely anatomically distinct—work as functionally integrated circuitry for mental calculation, because they were identified applying parametric analyses to fMRI data of the same task. However, whether or not the overlap observed in Fig. 1d indicates the actual connection between the two networks cannot be decided based on the present DTI data (cf. Wedeen et al. 2012).

Number magnitude-related (red) and fact retrieval-related (blue) network in number bisection. a Seed regions associated with number magnitude processing (red) or fact retrieval (blue). b 3D volume rendering with PIBI values >0.0148 the identified pathways for the more difficult magnitude-related conditions in red (left side). On the right side, a detailed view on the course of the fiber tracts is provided in axial orientation. c Pathways reflecting the fact retrieval-related conditions in blue (as 3D volume rendering with PIBI values >0.0148 on the left side and on axial slices on the right). d Overlay on the number magnitude-related (red) and fact retrieval-related (blue) network (as 3D volume rendering and on axial slice orientation). Two anatomically largely distinct dorsal vs. ventral fiber pathway profiles for the processing of number magnitude- and fact retrieval-related problems can be observed, differing not only in localization of activation but also in the connections between associated cortical areas and overlapping to a very limited degree only in the vicinity of the angular gyrus and EC/EmC system. Additionally, the connection between visual number form area (VNF) and the number magnitude representation (IPS/pIPS) is displayed in red. EC/EmC external/extreme capsule, IPS intraparietal sulcus, pIPS posterior intraparietal sulcus, PIBI probability index forming part of the bundle of interest
In particular, areas associated with magnitude-related processing were connected by both dorsal and ventral pathways. Fibers from SMA, IFGoper (Area 44), and IFGtri (Area 45) ran rostrally in ventral pathways of the EC/EmC system first, before merging into lateral and medio-dorsal parts of the SLF, leading towards the parietal seed-points IPS and pIPS. In contrast, FEF seemed to be connected via SLF only. The connection of the visual number form area in the fusiform gyrus to the left IPS was arching around the occipital horn of the lateral ventricle and probably sharing fibers with the inferior longitudinal fascicule (ILF), the inferior fronto-occipital fascicle (IOF) and/or the posterior parts of the posterior thalamic radiation.
For areas associated with arithmetic fact retrieval, predominantly ventral pathways were observed. AG was connected to posterior MTG, the VMPFC, and the hippocampus exclusively via ventral pathways encompassing the middle longitudinal fascicle (MdLF) and the EC/EmC. The short connections from SMG to AG travelled an almost identical ventral course as the connections from anterior MTG to the AG, possibly also including association fibers. Additionally, a dorsal connection via the cingulate bundle (and probably touching small parts of the corpus callosum) was observed linking RC with VMPFC. The connections between the AG and the RC (via callosal fibers) as well as between the AG and the insula (via SLF) ran predominantly dorsal. Nevertheless, the ventrally oriented MdLF fiber system was the main fiber pathway connecting frontal and parietal areas of the fact-retrieval network. The connection of the AG to the hippocampus ran via the RC first and then arching around the occipital horn of the lateral ventricle from superior to inferior until reaching the hippocampus, possibly sharing fibers from the retro-lenticular part of the posterior thalamic radiation. Between RC and hippocampus an additional connection was observed starting from the posterior thalamic radiation, arching in medial direction over the corpus callosum, then running to inferior and then rostral, probably sharing sparse medial cingulate fibers, and finally reaching from anterior and inferior caudally into the hippocampus.
Exact/approximate addition task
Similar to the NBT, evaluation of the overlay of magnitude and fact retrieval-related processing suggests two largely separate networks. The networks differed in localization of the seed regions (Fig. 2a; Table 2) and the connections between them (Fig. 2b–d: 3D presentation and axial slices; see Figure S6 for more details and different views). Both networks involved dorsal and ventral pathways but only overlapped to a very limited degree in the white matter next to the AG and the EC/EmC system. Again, based on the present data it cannot be decided whether or not the overlap observed in Fig. 2d indicated a direct connection between the two networks.

Number magnitude- (red) and fact retrieval-related (blue) network in exact/approximate arithmetic a Seed regions associated with number magnitude processing (red) or fact retrieval (blue). b 3D volume rendering with PIBI values >0.0148 (left side) the identified pathways for the more difficult magnitude-related conditions in red. A detailed view on the course of the fiber tracts is provided in axial orientation on the right side. c Pathways reflecting the fact retrieval-related conditions in blue (as 3D volume rendering with PIBI values >0.0148 on the left side and on axial slices on the right). d Overlay on the number magnitude-related (red) and fact retrieval-related (blue) network in exact and approximate arithmetic. Comparable with the results for NBT, two anatomically largely distinct dorsal vs. ventral fiber pathway profiles for the processing of number magnitude- and fact retrieval-related problems can be observed, differing not only in localization of activation but also in the connections between associated cortical areas and overlapping to a very limited degree only in the vicinity of the angular gyrus and the EC/EmC system. Additionally, the connection between visual number form area (VNF) and the number magnitude representation (IPS/pIPS) is displayed in red. EC/EmC external/extreme capsule, IPS intraparietal sulcus, pIPS posterior intraparietal sulcus, PIBI probability index forming part of the bundle of interest
Areas associated with magnitude-related processing were connected via both dorsal (SLF) and ventral pathways (more superior and lateral part of the EC/EmC system). The majority of fibers from FEF and IFGtri (Area 45) ran in lateral and medio-dorsal parts of the SLF to the parietal seed-points IPS and pIPS. However, ventral connections were also observed. Rostrally, connections from IPS to BA 47 and SMA also travelled ventrally encompassing the lateral and superior part of the EC/EmC system. The connection of the VNF area in the fusiform gyrus to the left IPS was again arching around the occipital horn of the lateral ventricle and probably sharing fibers with the posterior parts of the posterior thalamic radiation.
For areas associated with fact retrieval-related processing, the fibers connecting STG and MTG showed considerable anatomical consistency across the middle longitudinal fascicle (MdLF), probably also sharing fibers with the ILF and the SLF, and converged into the subinsular white matter near the claustrum. This bundle seemed to encompass the EC/EmC system in a more inferior part as compared to the fibers involved in magnitude-related processing. Furthermore, similar to the NBT, a dorsal connection via the cingulate bundle was observed linking RC with VMPFC. Moreover, the pathway to the insula travelled a more dorsal route via the SLF. Similar to the NBT, the connection of the AG to the hippocampus ran via the RC first and from the RC to the hippocampus in two different fibers around the occipital horn of the lateral ventricle and in medial direction over the corpus callosum.
Discussion
The current study aimed at extending the TCM (Dehaene et al. 2003; Arsalidou and Taylor 2011) by specifying the neural connections between cortical areas involved in numerical cognition with a particular focus on the important representational differentiation of the TCM: the representation of number magnitude and the verbal representation of arithmetic fact knowledge. To pursue these objectives, we performed probabilistic fiber tracking on seed regions identified by re-analyzing fMRI data of a number bisection task (NBT; Wood et al. 2008) and an exact/approximate addition task (Klein et al. 2009), as well as theoretical considerations suggesting the involvement of prefrontal and (para)hippocampal areas.
First, we found the visual number form area (VNF) to be involved (NBT and exact/approximate addition) and to be connected via similar fibers to the IPS in both tasks. Second, the numerical magnitude representation in the bilateral IPS was identified in both tasks. In line with our hypotheses, the IPS was not only connected via dorsal pathways to the VNF, but also via dorsal and ventral pathways to specific prefrontal areas. Third, in both tasks we observed that areas associated with fact retrieval such as the left AG were activated together with additional areas associated with retrieval from long-term memory. We found these areas to be connected both via dorsal and ventral fibers within a fronto-parietal network. Fourth, in line with the propositions of the TCM, activations in prefrontal cortex were identified in both tasks, which were also connected via similar pathways. In the following, we will first discuss these findings before integrating them into a schematic update of the TCM.
Fiber pathways in numerical cognition
The re-analysis of two different numerical tasks converged on a set of activation peaks and fiber pathways for number magnitude processing and fact retrieval which were largely identical across tasks. In line with the results of Klein et al. (2013), at least two different networks seem to be involved in numerical cognition: a network underlying magnitude-related processes and an anatomically largely distinct network subserving verbally mediated fact retrieval (Figs. 1, 2). With respect to the TCM the magnitude-related network reflects an amendment to the analog magnitude representation by its associated connectivity pattern, while the fact retrieval-related network is an extension of the verbal representation specifying its connectivity (see Fig. 3). Importantly, number magnitude manipulations and fact retrieval both seemed to be processed via dorsal and ventral pathways. However, these pathways were not identical across functional networks. In particular, there was only very limited overlap in white matter next to the AG and the EC/EmC system. For magnitude-related processing, the dorsal pathways involved the SLF-system for the connection of intraparietal cortex with frontal areas, while the ventral connections from IPS to BA 47 of the IFG and to the SMA predominantly followed a route via the lateral and superior part of the EC/EmC system. These tracking results for the network of magnitude-related processing fit nicely with the results of a recent meta-analysis by Arsalidou and Taylor (2011) for mental arithmetic. Both parietal as well as prefrontal coordinates identified by Arsalidou and Taylor (2011) to reflect cortical areas involved in magnitude processing fell within the spheres we defined as seed regions for our trackings. For fact retrieval-related processing, a dorsal connection via the cingulate bundle connected retro-splenial cortex (RC) with medial frontal cortex (VMPFC), while the ventral pathways encompassed the EC/EmC system. Additionally, in fact retrieval, the AG was connected to the hippocampus via retro-splenial cortex. In the present study, we focused on fiber pathways connecting seed regions identified to be involved in two different numerical tasks, which also meet the coordinates described by Arsalidou and Taylor (2011) in their recent meta-analysis. However, in numerical tasks with high demands on working memory an additional involvement of the middle frontal gyrus (MFG) is often observed. Nevertheless, this MFG involvement does not seem to be specific to number processing itself (Pesenti et al. 2000). Further studies are needed to investigate, which fiber tracts besides the SLF connect the MFG with number-specific parietal cortical areas.

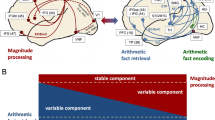
Dorsal (red) and ventral (green) processing pathways suggested to be included in the TCM (Dehaene et al. 2003) Based on the current results, two anatomically largely distinct dorsal (purple lines) vs. ventral (green lines) fiber pathway profiles are assumed for magnitude-related processing (a) and for (verbally mediated) fact retrieval (b). The colour changing arrow between a and b illustrates the idea that, even though distinct anatomically, these networks operate together as a functionally integrated circuit in numerical cognition, but with differential involvement depending on task characteristics. In a, a dashed line depicts the dorsal pathway from the visual number form (VNF) situated in the fusiform gyrus to the magnitude representation in the IPS. In b, areas on the surface of the brain are depicted in dark blue. c The interplay between magnitude-related processing and arithmetic fact retrieval. Basic fact retrieval-related processing is assumed to be independent of the difficulty of the arithmetic task, depicted as the blue rectangle at the bottom. However, in easier tasks a larger variable component of fact retrieval may be recruited (blue triangle), whereas in more difficult tasks more and more magnitude-related processing is assumed (red triangle). On the other side, an invariant component of mandatorily involved magnitude-related processing is assumed as well, depicted as the red rectangle at the top of the figure
It seems to be the case that both networks—though seemingly anatomically largely distinct—work as functionally integrated circuitry for mental calculation. This view is supported by the fact that in both tasks the activation peaks for magnitude-related and retrieval-related processing were identified applying the same parametric predictors (cf. Klein et al. 2013). Therefore, the distinction between magnitude-related processing and fact retrieval-related processing may not be a question of “either/or”, but may rather indicate that number magnitude information and arithmetic facts are recruited concomitantly, since we observed significant concordant and discordant fMRI changes. Generally we did not only find more activation in magnitude-related mental arithmetic; instead, we found different localizations of the activation peaks for verbally mediated and magnitude-related processes in mental arithmetic. This further corroborates our interpretation that the observed difference between the underlying networks may reflect qualitative processing differences rather than a merely quantitative effect of task difficulty.
Yet, one might argue that our approach will, by necessity, identify non-overlapping seed regions, because positive and negative contrasts for parametric regressors were evaluated. This leads to the identification of two non-overlapping networks. Taking the TCM by Dehaene and colleagues (1995, 1997, 2003) as a basis, this was exactly what was to be expected. In the TCM distinct areas are associated with either verbally mediated fact retrieval or magnitude manipulation. Therefore, the constellation of two different functional networks, which do not share a seed-point, was actually a necessary prerequisite for our fiber tracking. When assuming two content-wise different networks, as proposed by the TCM, one must not mix up functional regions from these two networks. Otherwise the requirements for probabilistic fiber tracking would be violated. One might still obtain a solution from the fiber tracking algorithm for an anatomical network, but it would not be functionally reasonable. Furthermore, it is worth emphasizing that in both networks both parietal and prefrontal seeds were incorporated. As described in detail by Caspers et al. (2011, e.g., Figure 7, p. 371) both the angular gyrus as well as the intraparietal cortex are connected to a variety of prefrontal, temporal and parietal seeds (see also Seghier 2013, who observed exactly the same connectivity for the angular gyrus as we found for the IPS). Against this background the current results appear even more interesting. Even though, both, the angular gyrus as well as large parts of the parietal cortex are connected to almost all prefrontal areas, we observed differential connectivity of the angular gyrus and the IPS with frontal areas within the respective networks of fact retrieval and magnitude manipulations.
Specificities of arithmetic fact retrieval
In both tasks we were able to identify similar areas, which have been proposed to be associated with arithmetic fact retrieval (i.e., left AG and left-hemispheric perisylvian language areas such as STG and MTG, Dehaene and Cohen 1997; Dehaene et al. 2003). However, we also identified additional neural activation associated with the retrieval of arithmetic facts from long-term memory in the hippocampus (Montaldi and Mayes 2010) as well as the recognition of familiarity in RC (see Vann et al. 2009 for a review). Interestingly, in both tasks the connection of the AG to the hippocampus ran via the RC first, only then arching around the occipital horn of the lateral ventricle from superior to inferior until reaching the hippocampus. This means that the AG and the hippocampus were not only connected via similar pathways in both tasks. Even more important, they were not connected directly but via the RC only. This is of particular interest, as the RC was also found to be activated in both tasks. Therefore, the RC may not only be an important structure in itself, probably bundling fibers, but it may also be involved in identifying familiar aspects. Familiarity information is then submitted to the hippocampus, which is supposed to integrate familiarity (e.g., from RC and AG) with recollection (e.g., from parahippocampal areas) to retrieve (arithmetic) fact information (Montaldi and Mayes 2010). From this triangle of fact retrieval found in both tasks (AG, RC, and hippocampus), only the AG was considered in the TCM so far (Dehaene et al. 2003). The AG indeed seems to work closely integrated with further perisylvian language areas as proposed by Dehaene and Cohen (1997). However, the whole network supporting fact retrieval seems to extend to the VMPFC as well (see Atique et al. 2011; Baetens et al. 2013; Spunt et al. 2011).
Qin et al. (2014) recently provided a comprehensive overview of the involvement of the hippocampus during arithmetic learning and arithmetic fact retrieval. Hippocampus activation was observed to be largest during learning retrieval-based solutions in children. In adolescents and adults, hippocampal activation was decreased but showed “higher interproblem stability” (p. 1267). This further corroborates the idea that the hippocampus is highly and actively engaged in learning and encoding of problems in close functional connection with frontal areas (Qin et al. 2014); however, as soon as the hippocampus is only needed for retrieval from long-term memory (e.g., in adults), its activation is less prominent but more stable. This is in line with other recent evidence. For instance, Cho et al. (2012) showed that children with better fluency in arithmetic fact retrieval revealed larger hippocampal fMRI signal. In this vein, Supekar et al. (2013) were able to show that larger hippocampal volume predicted learning improvements.
These results are consistent with the update for the TCM proposed in the current study. In adults, not only the angular gyrus, but also the hippocampus and the RC were observed to be engaged in arithmetic fact retrieval. The connection of the hippocampus with the AG in both tasks (NBT, exact/approximate task) was via the RC first, which is involved in the retrieval of familiarity (Montaldi and Mayes 2010) and well connected with the hippocampus via two separate bundles. The actual “arithmetic fact retrieval” is then an integration of familiarity and recollection processes in the hippocampus (Montaldi and Mayes 2010). However, while these retrieval processes are situated in parietal areas, the actual encoding of the facts seems to require much more effort in the hippocampus and a close connection to frontal areas (Qin et al. 2014; Supekar et al. 2013). Therefore, support of the (verbal) representation of arithmetic facts, as proposed by the TCM, clearly needs to be extended to the hippocampus (and probably parahippocampus and RC).
One might argue that the connectivity of the retro-splenial cortex (RC) with the AG and the VMPFC seems to resemble the connectivity pattern of the so-called default-mode network. This network is supposed to show higher activity and connectivity during resting state than during a cognitive task (e.g., Rosenberg-Lee et al. 2011; Wu et al. 2009; see Raichle et al. 2001 for more other cognitive domains). However, increased connectivity during resting state does not necessarily mean that this network is not recruited in active tasks—rather, there is increasing evidence to the contrary. Areas already well connected in the resting state are perfect candidates as members of a fast and automatized network in active tasks. In particular, there is now evidence that the RC response (e.g., during tests of spatial navigation with memorized cues as well as autobiographical memory retrieval) is significantly above resting state baseline, reflecting true engagement of the RC in performing such tasks (e.g., Epstein 2008; Maguire 2001; Svoboda et al. 2006). Additionally, activation of the hippocampus in our fMRI data suggests that participants actively retrieved facts from long-term memory in the easier task conditions. Furthermore, an example for the easiest condition in Klein et al. (2009) would be, “23 + 46 = 69 or 83” (easy because no carry needed, problem size of the addends’ digits is rather small, and the distance between target and distractor is large), which had to be solved within 3.5 s. While this easy condition was answered up to 200 ms faster than the most difficult one (e.g., “46 + 39 = 82 or 76”, difficult because carry needed, correct result not given, and small distractor distance), participants nevertheless struggled with the time limit of 3.5 s. This is hard to reconcile with the assumption that participants were “in resting state” while they solved these problems. Thereby, these data provide evidence against the default-mode network hypothesis—making the interpretation that the activity between areas involved in the easier conditions merely reflects stronger involvement of the default-mode network unlikely.
In summary, the present study suggests that the network subserving fact retrieval in the left AG and perisylvian language areas (as described by Dehaene et al. 2003) needs to be extended to a network connecting the AG with RC, VMPFC, and the hippocampus. The hippocampus is involved in retrieval from long-term memory (Baddeley 1966; Cansino et al. 2002 for fMRI data; Montaldi and Mayes 2010; Rugg and Yonelinas 2003 for a review), while the retro-splenial cortex and the VMPFC have repeatedly been associated with the recognition of familiar objects or procedures (Vann et al. 2009)—cognitive processes vitally involved in the retrieval of arithmetic fact knowledge associated with the AG.
An update for the TCM
The present results suggest an amendment to the TCM: as regards the representation of number magnitude, we suggest to extend its anatomo-functional instantiation towards a network including not only the IPS but also additional cortical areas as well as the connectivity between these areas (Fig. 3a). In particular, four prefrontal areas were engaged in both numerical tasks: Broca’s Area (IFGtri, Area 45) as well as BA 47, SMA, and FEF were found active consistently in both numerical tasks. Additionally, they were also connected via similar pathways. We propose to include IFGtri (Area 45), BA 47, and the SMA as additional prefrontal substrates associated with number magnitude processing. The bilateral SMA has been suggested to play a supportive role for multi-componential cognitive functions, such as mathematical operations and carrying out supporting arithmetic operations (Kong et al. 2005; Krueger et al. 2011), while Area 45 and BA 47 seem to be associated with the application of rules (Weiller et al. 2011). Because it is not clear whether the FEF activation indeed reflects functionally relevant orientation on the mental number line rather than eye movements when re-checking the problems presented, we do not include the FEF at this point. We are confident that the additional areas incorporated in our model are not specific for the tasks studied, but are essential for number processing in general (see also Krueger et al. 2011 for the case of the SMA).
With respect to the verbally mediated representation of arithmetic facts, we also suggest to amend the anatomo-functional association by both, additional cortical areas as well as their connections (Fig. 3b). In particular, the present study suggests that the verbal representation needs to be extended to a network subserving fact retrieval including connections of the AG to the RC and further on via dorsal and ventral fiber pathways to subcomponents of the mentalizing network (VMPFC) and the hippocampus. RC and VMPFC may subserve the automatic recognition of familiar or anticipated arithmetic facts, while the hippocampus not only reflects retrieval of arithmetic facts from long-term memory (e.g., Klein et al. 2013), but has also been associated with the recognition of familiarity as well (Rugg and Yonelinas 2003 for a review).
Mental exact/approximate addition and NBT: differences and commonalities
We evaluated the neural networks recruited in two substantially different numerical tasks and observed not only common, but also differing activation peaks. For instance, in the NBT, but not the exact/approximate task, Area 44 of the inferior frontal gyrus, which is assumed to subserve verbal working memory (Weiller et al. 2011), was observed to be activated. However, this may just reflect differences between the two tasks. Performing the number bisection task for a triplet of two-digit numbers with a large range (e.g., 31_39_47) may most likely require more processing steps (e.g., calculating the difference between the first and the second number, adding this difference to the second number and comparing the result to the third) with intermediate results to be kept in mind, rather than the mere addition of two two-digit numbers. Additionally, the influence of fact retrieval (i.e., the advantage for multiplicative triplets such as 21_24_27) is much more implicit than in active calculation, as required in exact/approximate addition (such as 2 + 3 = 5 in 42 + 32 = 75). Put differently, the number bisection task not only requires more but also different additional cognitive components and/or strategies than exact/approximate addition. In fact, having a closer look at these two numerical tasks, it is even more notable that we found so many similar or even identical grey matter activation spots as well as fiber pathways.
Interrelation between the two networks
As has been outlined before (Klein et al. 2013), we suggest the magnitude-related network and the verbally mediated fact retrieval network to operate in a closely integrated fashion. Nevertheless, the exact way, in which the two networks work together, and the question through which anatomical structures this cooperation is specified, awaits further clarification. We suggest invariant as well as variable components of both number magnitude processing and fact retrieval (cf. Fig. 3c) to be activated whenever one encounters a numerical problem irrespective of the task or its difficulty. On the one hand, it has been shown repeatedly (e.g., Eger et al. 2003; Klein et al. 2010) that the IPS is activated automatically whenever we see or hear numerical stimuli. On the other hand, Rusconi et al. (2006) observed that in a number-matching task the product of two digits was also activated automatically. These stable components of number magnitude and arithmetic facts are complemented by variable ones, whose activation depends on task requirements. With increasing task difficulty the influence of fact retrieval processes may decrease, whereas the influence of magnitude-related processes may increase complimentarily. However, we assume processes of fact retrieval to also take effect in difficult arithmetic. This proposition is corroborated by participants breaking down complex problems into more tractable sub-problems, the solutions of which are retrieved from long-term memory (e.g., splitting 32 + 43 in 3 + 4 and 2 + 3 = 5; e.g., LeFevre et al. 1996). In this way, our model is not only capable of accounting for fact-retrieval deficits often exhibited by brain-damaged patients, by assuming an interruption of the pathways underlying arithmetic fact retrieval (e.g., Zaunmueller et al. 2009). This also offers an explanation for the often severe problems in more difficult arithmetic tasks observed after brain damage. Because a stable component of arithmetic fact retrieval is also involved in more difficult problems, loss of arithmetic fact knowledge will influence difficult arithmetic as well. Furthermore, we want to emphasize that we do not claim that the brain networks identified in the current study (i.e., cortical areas and their structural connectivity) are specific to numerical tasks only (see Willmes et al. 2014). For instance, the fact retrieval network (or at least parts of it) may be involved in non-numerical tasks, which require retrieval of factual knowledge from long-term memory (e.g., Addis et al. 2004; Montaldi and Mayes 2010; Piefke et al. 2003).
As regards the anatomical correlates of the interaction of the two networks, the present study did not aim to find the exact location of this interaction. Thus, one may only speculate in which structures the two networks might interact. From an anatomical point of view, this might most probably be between the AG and IPS. IPS and the AG are not only anatomically close (IPS delineates the upper border of AG), but are also closely connected via association fibers and most probably via U-fibers as well (Caspers et al. 2011). Moreover, there is a rich body of literature suggesting that the upper part of the AG is central for mental arithmetic (e.g., Anderson 2005; Anderson et al. 2003). However, this idea remains to be substantiated by future research.
In a next step, it would be interesting to investigate the interrelation between the two networks in a more dynamic context. In the present study, we looked for stable structural connectivity. In a training study, it seems reasonable to assume that tasks, which are predominantly solved by number magnitude manipulations in the beginning, may be solved by arithmetic fact retrieval with increasing proficiency (cf. Ischebeck et al. 2007 for on-line monitoring of training effects in an fMRI session). In this context, it would also be desirable to examine not only the structural, but also the effective connectivity among these networks. For instance, employing multivariate dynamic causal modeling (DCM), it would be required to further specify the direction and strength of the connections identified in the present study.
Notes
We re-ran the analysis of tracts omitting the left-handed participant. However, since this did neither alter the results substantially nor even visibly, we decided to keep all participants.
References
Addis DR, Moscovitch M, Crawley AP, McAndrews MP (2004) Recollective qualities modulate hippocampal activation during autobiographical memory retrieval. Hippocampu 14(6):752–762
Anderson JR (2005) Human symbol manipulation within an integrated cognitive architecture. Cogn Sci 29(3):313–341
Anderson JR, Qin Y, Sohn MH, Stenger VA, Carter CS (2003) An information-processing model of the BOLD response in symbol manipulation tasks. Psychon Bull Rev 10(2):241–261
Arsalidou M, Taylor MJ (2011) Is 2 + 2 = 4? Meta-analyses of brain areas needed for numbers and calculations. NeuroImage 54:2382–2393
Ashcraft MH, Kirk EP (2001) The relationships among working memory, math anxiety, and performance. J Exp Psychol Gen 130:224–237
Atique B, Erb M, Gharabaghi A, Grodd W, Anders S (2011) Task-specific activity and connectivity within the mentalizing network during emotion and intention mentalizing. Neuroimage 55(4):1899–1911
Baddeley AD (1966) The influence of acoustic and semantic similarity on long-term memory for word sequences. Q J Exp Psychol 18(4):302–309
Baetens K, Ma N, Steen J, Van Overwalle F (2014) Involvement of the mentalizing network in social and non-social high construal. Soc Cogn Affect Neurosci 9:817–824
Basser PJ, Mattiello J, Le Bihan D (1994) Estimation of the effective self-diffusion tensor from the NMR spin echo. J Magn Res B 103:247–254
Bürgel U, Amunts K, Hoemke L, Mohlberg H, Gilsbach JM, Zilles K (2006) White matter fiber tracts of the human brain: three-dimensional mapping at microscopic resolution, topography and intersubject variability. Neuroimage 29(4):1092–1105
Cansino S, Maquet P, Dolan RJ, Rugg MD (2002) Brain activity underlying encoding and retrieval of source memory. Cereb Cortex 10:1048–1056
Caspers S, Geyer S, Schleicher A, Mohlberg H, Amunts K, Zilles K (2006) The human inferior parietal cortex: cytoarchitectonic parcellation and interindividual variability. Neuroimage 33(2):430–448
Caspers S, Eickhoff SB, Rick T, von Kapri A, Kuhlen T, Huang R, Shah NJ, Zilles K (2011) Probabilistic fibre tract analysis of cytoarchitectonically defined human inferior parietal lobule areas reveals similarities to macaques. Neuroimage 58(2):362–380
Catani M, Thiebaut de Schotten M (2012) Atlas of Human Brain Connections. Oxford University Press
Cho S, Metcalfe AWS, Young CB, Ryali S, Geary DC, Menon V (2012) Hippocampal-prefrontal engagement and dynamic causal interactions in the maturation of children’s fact retrieval. J Cogn Neurosci 24:1849–1866
Dehaene S, Cohen L (1995) Towards an anatomical and functional model of number processing. Math Cogn 1:83–120
Dehaene S, Cohen L (1997) Cerebral pathways for calculation: double dissociation between rote verbal and quantitative knowledge of arithmetic. Cortex 33:219–250
Dehaene S, Piazza M, Pinel P, Cohen L (2003) Three parietal circuits for number processing. Cogn Neuropsychol 20:487–506
Delazer M, Domahs F, Bartha L, Brenneis C, Lochy A, Trieb T, Benke T (2003) Learning complex arithmetic—a fMRI study. Cogn Brain Res 18:76–88
Eickhoff S, Stephan KE, Mohlberg H, Grefkes C, Fink GR, Amunts K, Zilles K (2005) A new SPM toolbox for combining probabilistic cytoarchitectonic maps and functional imaging data. NeuroImage 25(4):1325–1335
Eger E, Sterzer P, Russ MO, Giraud AL, Kleinschmidt A (2003) A supramodal number representation in human intraparietal cortex. Neuron 37:719–725
Epstein RA (2008) Parahippocampal and retrosplenial contributions to human spatial navigation. Trends Cogn Sci 12:388–396
Grabner RH, Ansari D, Koschutnig K, Reishofer G, Ebner F, Neuper C (2009) To retrieve or to calculate? Left angular gyrus mediates the retrieval of arithmetic facts during problem solving. Neuropsychologia 47:604–608
Imbo I, Vandierendonck A, De Rammelaere S (2007a) The role of working memory in the carry operation of mental arithmetic: number and value of the carry. Q J Exp Psychol 60:708–731
Imbo I, Vandierendonck A, Vergauwe E (2007b) The role of working memory in carrying and borrowing. Psychol Res 71:467–483
Ischebeck A, Zamarian L, Egger K, Schocke M, Delazer M (2007) Imaging early practice effects in arithmetic. NeuroImage 36:993–1003
Klein E, Nuerk HC, Wood G, Knops A, Willmes K (2009) The exact vs. approximate distinction in numerical cognition may not be exact, but only approximate How different processes work together in multi-digit addition. Brain Cogn 2:369–381
Klein E, Moeller K, Nuerk HC, Willmes K (2010) On the cognitive foundations of basic auditory number processing. Behav Brain Funct 6:42
Klein E, Moeller K, Glauche V, Weiller C, Willmes K (2013) Processing pathways in mental arithmetic—evidence from probabilistic fiber tracking. PLoS ONE 8(1):e55455
Kong J, Wang C, Kwong K, Vangel M, Chuac E, Gollub R (2005) The neural substrate of arithmetic operations and procedure complexity. Cogn Brain Res 22:397–405
Kreher BW, Schnell S, Mader I, Ilyasov KA, Hennig J, Kiselev V, Saur D (2008) Connecting and merging fibres: pathway extraction by combining probability maps. NeuroImage 43:81–89
Krueger F, Landgraf S, van der Meer E, Deshpande G, Hu X (2011) Effective connectivity of the multiplication network: a functional MRI and multivariate Granger Causality Mapping study. Hum Brain Mapp 32(9):1419–1431. doi:10.1002/hbm.21119
LeFevre JA, Sadesky GS, Bisanz J (1996) Selection of procedures in mental addition: reassessing the problem size effect in adults. J Exp Psychol Learn Mem Cogn 22:216–230
Maguire EA (2001) The retrosplenial contribution to human navigation: a review of lesion and neuroimaging findings. Scand J Psychol 42:225–238
Miller EK (2000) The prefrontal cortex and cognitive control. Nat Rev Neurosci 1:58–65
Montaldi D, Mayes AR (2010) The role of recollection and familiarity in the functional differentiation of the medial temporal lobes. Hippocampus 20(11):1291–1314
Newton JM, Ward NS, Parker GJM, Deichmann R, Alexander DC, Friston K, Frackowiak RSJ (2006) Non-invasive mapping of corticofugal fibres from multiple motor areas—relevance to stroke recovery. Brain 129:1844–1858
Oishi K, Faria AV, Zijl PCM, Mori S (2011) MRI atlas of the human white matter, 2nd edn. Academic Press, New York
Parker GJM, Haroon HA, Wheeler-Kingshott CAM (2003) A framework for a streamline-based probabilistic index of connectivity (PICo) using a structural interpretation of MRI diffusion measurements. J Magn Res Imaging 18:242–254
Pesenti M, Thioux M, Seron X, De Volder A (2000) Neuroanatomical substrates of Arabic number processing, numerical comparison and simple addition: a PET study. J Cogn Neurosci 12(3):461–479
Piefke M, Weiss PH, Zilles K, Markowitsch HJ, Fink GR (2003) Differential remoteness and emotional tone modulate the neural correlated of autobiographical memory. Brain 126:650–668
Qin S, Cho S, Chen T, Rosenberg-Lee M, Geary DC, Menon V (2014) Su Hippocampal-neocortical functional reorganization underlies children’s cognitive development. Nat Neurosci 17:1263–1269
Raichle ME, MacLeod AM, Snyder AZ, Powers WJ, Gusnard DA, Shulman GL (2001) A default mode of brain function. Proc Natl Acad Sci USA 98(2):676–682
Rosenberg-Lee M, Chang TT, Young CB, Wu S, Menon V (2011) Functional dissociations between four basic arithmetic operations in the human posterior parietal cortex: a cytoarchitectonic mapping study. Neuropsychologia 49(9):2592–2608
Rugg MD, Yonelinas AP (2003) Human recognition memory: a cognitive neuroscience perspective. Trends Cogn Sci 7(7):313–319
Rusconi E, Galfano G, Rebonato E, Umiltà C (2006) Bidirectional links in the network of multiplication facts. Psychol Res 70(1):32–42
Saur D, Kreher BW, Schnell S, Kuemmerer D, Kellermeyer P, Vry MS, Umarova RM, Musso M, Glauche V, Abel S, Huber W, Rijntjes M, Hennig J, Weiller C (2008) Ventral and dorsal pathways for language. Proc Natl Acad Sci USA 105:18035–18040
Saur D, Schelter B, Schnell S, Kratochvil D, Küpper H, Kellmeyer P, Kümmerer D, Klöppel S, Glauche V, Lange R, Mader W, Feess D, Timmer J, Weiller C (2010) Combining functional and anatomical connectivity reveals brain networks for auditory language comprehension. NeuroImage 49:3187–3197
Scheperjans F, Eickhoff SB, Hömke L, Mohlberg H, Hermann K, Amunts K, Zilles K (2008) Probabilistic maps, morphometry, and variability of cytoarchitectonic areas in the human superior parietal cortex. Cereb Cortex 18(9):2141–2157
Schmahmann JD, Pandya DN (2006) Fiber pathways of the brain. Oxford Univ Press, New York
Seghier ML (2013) The angular gyrus: multiple functions and multiple subdivisions. Neuroscientist 19(1):43–61
Spunt RP, Satpute AB, Lieberman MD (2011) Identifying the what, why, and how of an observed action: an fMRI study of mentalizing and mechanizing during action observation. J Cogn Neurosci 23(1):63–74
Supekar K, Swigart AG, Tenison C, Jolles DD, Rosenberg-Lee M, Fuchs L, Menon V (2013) Neural predictors of individual differences in response to math tutoring in primary-grade school children. Proc Natl Acad Sci USA 110:8230–8235
Svoboda E, McKinnon MC, Levine B (2006) The functional neuroanatomy of autobiographical memory: a meta-analysis. Neuropsychologia 44:2189–2208
Uddin LQ, Supekar K, Amin H, Rykhlevskaia E, Nguyen DA, Greicius MD, Menon V (2010) Dissociable connectivity within human angular gyrus and intraparietal sulcus: evidence from functional and structural connectivity. Cereb Cortex 20(11):2636–2646
Vann SD, Aggleton JP, Maguire EA (2009) What does the retrosplenial cortex do? Nat Rev Neurosci 10:792–802
Wedeen VJ, Rosene DL, Wang R, Dai G, Mortazavi F, Hagmann P, Kaas JH, Tseng WY (2012) The geometric structure of the brain fiber pathways. Science 335(6076):1628–1634
Weiller C, Bormann T, Saur D, Musso M, Rijntjes M (2011) How the ventral pathway got lost—and what its recovery might mean. Brain Lang 118:29–39
Willmes K, Moeller K, Klein E (2014) Where numbers meet words—a common ventral network for semantic classification. Scand J Psychol 55:202–211
Wood G, Nuerk HC, Moeller K, Geppert B, Schnitker R, Weber J, Willmes K (2008) All for one but not one for all: how multiple number representations are recruited in one numerical task. Brain Res 1187:154–166
Wu SS, Chang TT, Majid A, Caspers S, Eickhoff SB, Menon V (2009) Functional heterogeneity of inferior parietal cortex during mathematical cognition assessed with cytoarchitectonic probability maps. Cereb Cortex 12:2930–2945
Zaitsev M, Hennig J, Ilyasov KA (2006) Geometric distortions applied to diffusion tensor imaging. Annual Meeting of the international society for magnetic resonance in medicine, Seattle
Zaunmueller L, Domahs F, Dressel K, Lonnemann J, Klein E, Ischebeck A, Willmes K (2009) Rehabilitation of arithmetic fact retrieval via extensive practice: a combined fMRI and behavioural case-study. Neuropsychol Rehabil 19:422–443
Acknowledgments
This research was supported by the German Research Foundation (DFG) to Klaus Willmes (Wl 1804/6-1), Hans-Otto Karnath (KA 1258/11-1), Korbinian Moeller (MO 2525/2-1), and André Knops (KN 959/2-1) as well as by the Leibniz-Competition Fund (SAW) providing funding to Elise Klein (SAW-2014-IWM-4). We are indebted to the “Brain Imaging Facility of the Interdisciplinary Centre for Clinical Research within the Faculty of Medicine at the RWTH Aachen University” for their precious help in acquiring the MRI/fMRI data.
Author information
Authors and Affiliations
Corresponding author
Electronic supplementary material
Below is the link to the electronic supplementary material.
Rights and permissions
About this article

Cite this article
Klein, E., Suchan, J., Moeller, K. et al. Considering structural connectivity in the triple code model of numerical cognition: differential connectivity for magnitude processing and arithmetic facts. Brain Struct Funct 221, 979–995 (2016). https://doi.org/10.1007/s00429-014-0951-1
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00429-014-0951-1