
Abstract
Glutamine metabolism is emerging as one aspect of dysregulated metabolism of tumors. Triple-negative breast cancer (TNBC) cells are glutamine dependent, whereas luminal-type cells tend to be glutamine independent. Therefore, TNBC patients might benefit from therapies targeting glutamine metabolism. To investigate the clinical significance of glutamine metabolism, we examined expression and prognostic significance of glutaminase in tumor cells and tumor-infiltrating lymphocytes (TILs) in TNBC. We retrieved 658 surgically resected TNBCs and analyzed glutaminase expression in tumor cells and TILs by immunohistochemical staining. Glutaminase expression was observed in 237 cases (36.0%) in tumor cells and 104 cases (15.5%) in TILs. Although glutaminase expression in tumor cells was significantly associated with a low level of TILs (p = 0.018), glutaminase expression in TILs was significantly higher in cases with a high level of TILs (p = 0.031). Glutaminase expression in tumor cells was significantly associated with poor disease-free survival in patients with lymph node metastasis and high levels of TILs (p = 0.020). In addition, it was an independent poor prognostic factor (hazard ratio = 10.643, 95% confidence interval = 1.999–56.668; p = 0.006). Glutaminase expression in tumor cells was observed in a subset of TNBC patients. It was significantly associated with a low level of TILs and poor disease-free survival in TNBCs presenting with lymph node metastasis and high levels of TILs.
Similar content being viewed by others
Avoid common mistakes on your manuscript.
Introduction
Cancer cells depend on energy for survival, proliferation, and invasion [1]. Features of dysregulated tumor metabolism, such as aerobic glycolysis, play a critical role in oncogenesis and represent potential diagnostic markers or therapeutic targets [2–4]. Recent studies indicate that glutamine metabolism is another key feature of dysregulated metabolism in tumors. Glutamine is needed to supply energy to cancer cells by generating cellular ATP and protein synthesis, as a metabolic intermediate for nucleotide synthesis and for c-MYC-mediated oncogenesis [5–10]. However, cancer cell lines show significant variability in response to glutamine deprivation indicating that not all cancer cells need exogenous glutamine for their survival [11]. In breast cancer, basal-type cells are glutamine dependent, whereas luminal-type cells tend to be glutamine independent [1]. Therefore, patients with triple-negative breast cancer (TNBC), which currently have few treatment options, might benefit from therapies that target glutamine metabolism. The conversion of glutamine to glutamate is mediated by glutaminolysis, a catabolic process mediated by two distinct glutaminase proteins (GLS1 and GLS2). Glutaminase mediates a signaling cascade associated with the c-MYC oncogene and presumably plays a critical role in glutamine metabolism [8, 9, 12, 13].
Tumor-infiltrating lymphocytes (TILs) are associated with good prognosis and predict response to therapy in TNBC [14–16]. TILs also need adequate nutrients and metabolism to function [17]. T cells show different metabolic demands according to their developmental stage. Naïve T cells are metabolically quiescent relative to effector and memory T cells. To promote cell growth and proliferation and ensure effective cell function, effector T cells become metabolically active and increase levels of glycolysis and glutaminolysis [17]. Metabolic competition between cancer cells and cells in the tumor microenvironment, such as TILs, might inhibit the protective role of T cells and subsequently drive cancer progression [18]. Chang et al. demonstrated that glucose consumption by tumors metabolically restricts T cells and promotes tumor progression in a mouse sarcoma model [18]. However, the correlation of glutaminase expression in cancer cells and TILs with respect to metabolic competition and the prognostic significance of glutaminase expression in TNBC has not been investigated. In this study, we examined expression of glutaminase in consecutive breast cancer and TNBC cohorts. We also evaluated the expression of glutaminase in TILs in TNBCs and analyzed its prognostic significance.
Materials and methods
Patient and tissue specimens
We evaluated two groups of patients with primary breast cancer. One group comprised 157 consecutively enrolled patients with breast cancer, who underwent surgical resection at the Asan Medical Center in Seoul, Korea in 1998 and from whom formalin-fixed, paraffin-embedded tissue samples were available for analysis. In this cohort, 97 of the 157 patients received six cycles of an adjuvant methotrexate-based regimen (cyclophosphamide, 500 mg/m2; methotrexate, 40 mg/m2; and 5-fluorouracil, 500 mg/m2) or an anthracycline-based regimen (cyclophosphamide, 500 mg/m2; adriamycin, 50 mg/m2; and 5-fluorouracil, 500 mg/m2). Five patients received adjuvant chemotherapy with an unknown regimen. In addition, 16.5 and 75.8% patients received radiotherapy and hormone therapy, respectively. The mean follow-up period was 13.4 years. The second group comprised 682 TNBC patients who underwent surgical resection between 2004 and 2010 at the Asan Medical Center, from whom tissue samples were available. In this group, 473 of the 682 patients did not present with lymph node metastasis. They received four cycles of adjuvant anthracycline and cyclophosphamide (AC, adriamycin, 60 mg/m2, and cyclophosphamide, 600 mg/m2). The remaining 209 patients presented with lymph node metastasis and were treated with four cycles of AC followed by either four cycles of paclitaxel (175 mg/m2) or four cycles of docetaxel (75 mg/m2). Five hundred fifty-one patients (80.8%) received radiotherapy, and one patient received hormone therapy. The median follow-up period was 78.3 months. All of the patients were chemotherapy naïve and radiotherapy naïve preoperatively. Clinical information was obtained from medical records and pathology reports from the surgery. Exemption from informed consent after de-identification of information was approved by the Institutional Review Board of the Asan Medical Center.
Histological evaluation
Hematoxylin and eosin (H&E)-stained slides were reviewed by two pathologists (H.J.L. and G.G.). Slides were histopathologically analyzed for the presence of TILs (defined as the percentage of the stroma of invasive carcinoma infiltrated by lymphocytes in 10% increments) [14], histological grade, tumor size, pT stage, pN stage, and lymphovascular invasion. TIL presence was dichotomized into high or low using 60% as cutoff, in agreement with the definition of lymphocyte-predominant breast cancer [19, 20]. Histological grade was assessed using the modified Bloom–Richardson classification [21].
Tissue microarray construction and immunohistochemistry
From formalin-fixed, paraffin-embedded tissue samples, a tissue microarray was constructed using an arraying instrument, as previously described [22]. Of each sample, three 1-mm-diameter cores were included in the array, to minimize tissue loss and mitigate the effects of tumor heterogeneity. Tissue microarray sections were stained using an automatic immunohistochemical staining device (Benchmark XT; Ventana Medical Systems, Tucson, AZ). Antibodies to target the estrogen receptor (ER, 1:100, Leica Biosystems, Newcastle, UK), progesterone receptor (PR, 1:200, Leica Biosystems), HER2 (1:8, Ventana Medical Systems), and glutaminase (1:100, Abcam, Cambridge, UK) were used. ER and PR levels were scored as positive if at least 1% tumor nuclei stained positive [23]. The hormone receptor (HR)-positive group was defined as ER- or PR-positive tumors. HER2-overexpressing tumors were defined as those with scores of 3+ by immunohistochemistry or gene amplification by fluorescence in situ hybridization (FISH) or silver in situ hybridization (SISH) [24].
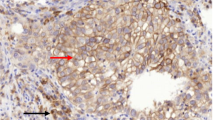
The intensity of glutaminase expression in the tumors was scored on a scale of 0–3 (Fig. 1). Cytoplasmic expression of glutaminase was also evaluated using an immunoreactivity score, which was generated by multiplying intensity and percentage of positive cells (0–300); scores >10 were classified as positive. The intensity of glutaminase staining in TILs was classified as none, weak, moderate, or strong (Fig. 1). The cases with moderate or strong intensity were considered positive [25].

Representative images of glutaminase immunohistochemical staining of tumor cells and TILs
Statistical analysis
All statistical analyses were conducted using SPSS statistical software version 18 (SPSS, Chicago, IL). The Chi-square test, log-rank test, and forward conditional Cox proportional hazards regression model were used to evaluate the data. All tests were two-sided, and p > 0.05 was considered statistically significant.
Results
Glutaminase expression in consecutive breast cancer cases
We analyzed glutaminase expression in the tumor cells of each subtype in a consecutive series of breast cancer cases. Glutaminase was expressed in 15 consecutive breast cancer cases (9.6%), 14 of which were TNBC (14 of 40 TNBC cases, 35%, p < 0.001, Table 1). There was one glutaminase-positive case in the HR-positive (weak ER expression, ER Allred score 4) and HER2-negative group. All HER2-positive cases were negative for glutaminase expression.
Glutaminase expression in the TNBC cohort and the correlation of glutaminase expression with clinical factors
As glutaminase expression was more prevalent in TNBC samples, we analyzed glutaminase expression in a large cohort of TNBC cases. Evaluation of immunohistochemical staining for glutaminase was possible on 658 cases regarding tumor cells and 673 cases regarding TILs. Clinicopathological characteristics of TNBC patients are summarized in Table 2. Glutaminase expression was positive in 237 cases (36.0%) in tumor cells and 104 cases (15.5%) in TILs. While positive glutaminase expression in tumor cells was significantly associated with a low level of TILs (p = 0.018), glutaminase expression in TILs was significantly higher in cases with a high level of TILs (p = 0.031, Table 2). There were no significant differences in histological grade, pT stage, lymphovascular invasion, lymph node metastasis, and radiotherapy between glutaminase-negative and glutaminase-positive cases.
Prognostic significance of glutaminase expression in TNBC
In the survival analysis, a higher pT stage (p = 0.002), the presence of lymphovascular invasion, metastasis, and low TIL levels (all, p < 0.001) were significantly associated with poor disease-free survival (Table 3). Lymph node metastasis and low level of TILs (both, p < 0.001) were independent poor prognostic factors. Subgroup analysis was done according to lymph node metastasis and levels of TILs. The tumor cell glutaminase expression was significantly associated with poor disease-free survival in patients with lymph node metastasis and a high level of TILs (p = 0.020, Table 4, Fig. 2a). In multivariate analysis, glutaminase expression in tumor cells was an independent poor prognostic factor (hazard ratio = 10.643, 95% confidence interval = 1.999–56.668; p = 0.006) along with the presence of lymphovascular invasion (hazard ratio = 16.372, 95% confidence interval = 1.920–139.629; p = 0.011). In other subgroups, glutaminase expression was not associated with disease-free survival (Tables 4 and 5, Fig. 2b, c, and d).

Disease-free survival analysis of TNBC subgroups according to tumoral glutaminase expression. a In TNBC patients with lymph node metastasis and high levels of TILs, glutaminase expression is associated with poor disease-free survival (p = 0.020). b–d In TNBC patients not presenting with lymph node metastasis or high levels of TILs, glutaminase expression is not significantly associated with disease-free survival. (b TNBC patients with lymph node metastasis and low TIL levels, p = 0.800; c without lymph node metastasis and high levels of TILs, p = 0.964; d without lymph node metastasis and low levels of TILs, p = 0.569)
Discussion
Glutamine metabolism is important for cancer cell survival [5, 26]. Glutamine plays a role in cancer cell proliferation by participating in ATP synthesis and generating intermediates for the synthesis of macromolecules [5]. Using immunohistochemical staining with an antibody against glutaminase, we demonstrate that glutaminase is almost exclusively expressed in TNBCs compared with that in other types of breast cancer. Kim et al. also noted that tumor glutaminase expression is higher in TNBC than in other subtypes of breast cancer, and in HR-positive tumors, glutaminase tends to be negative [25], which was in accordance with our results. A previous study reported that in luminal-type breast cancer, glutamine is synthesized by glutamine synthetase, but not in basal-like type breast cancer [1]. Thus, basal-like breast cancer or TNBC show a high level of glutaminase expression and therefore appear more glutamine dependent than other subtypes of breast cancer [1]. TNBC might show a high rate of glutamine metabolism, because TNBC usually have features correlated with high metabolic activity, such as high histologic grade, increased mitosis, and tumor necrosis [25].
Glutaminase expression in tumor cells was significantly correlated with a low level of TILs, suggesting that active consumption of glutamine in tumor cells precludes effective utilization of glutamine by TILs and hinders proliferation of TILs in TNBCs with a high expression of glutaminase. We evaluated the prognostic significance of glutaminase expression in TNBCs and found that tumor glutaminase expression is significantly associated with poor disease-free survival in TNBCs with lymph node metastasis and a high level of TILs. Although a high level of TILs was associated with better clinical outcome in patients with TNBC, not all patients with a high level of TILs remained free of recurrence or alive [27]. Few prognostic indicators are associated with TNBCs with a high level of TILs. Our findings suggest that glutaminase expression in tumor cells might be a valuable prognostic marker of TNBCs presenting with a high level of TILs. Our findings are also consistent with the concept of metabolic competition in the tumor microenvironment, as a high level of glutaminase expression in tumor cells might result in glutamine deprivation in TILs, thereby hindering the protective role of T cells in cancer progression [18].
Poor prognosis of TNBC with lymph node metastasis and a high level of TILs might be associated with hypoxia-inducible factor 1 (HIF1) promoting invasion, growth, and metastasis of TNBC cells [28–30]. High glutaminase activity induces a high level of extracellular glutamate, which inhibits intracellular cysteine uptake via the xCT glutamate-cystine antiporter [28]. Depletion of intracellular cysteine directly inhibits degradation of HIF1α resulting in intracellular HIF1α accumulation [28].
In developing new modalities of cancer treatment, observations on cancer metabolism beyond the Warburg effect, such as discovery of targetable metabolomic pathway and identification of somatic mutations in metabolic enzymes, might be relevant [31–36]. However, it would be important to selectively target metabolism of cancer cells because TILs also need increased glycolysis and glutaminolysis for cell survival and function and compete with cancer cells for metabolite consumption [17, 18, 37].
In summary, glutaminase expression in tumor cells was almost exclusively present in TNBCs and significantly correlated with a low level of TILs. High glutaminase in TNBCs presenting with lymph node metastasis and high levels of TILs was significantly associated with poor disease-free survival.
References
Kung HN, Marks JR, Chi JT (2011) Glutamine synthetase is a genetic determinant of cell type-specific glutamine independence in breast epithelia. PLoS Genet 7:e1002229. doi:10.1371/journal.pgen.1002229
Hsu PP, Sabatini DM (2008) Cancer cell metabolism: Warburg and beyond. Cell 134:703–707. doi:10.1016/j.cell.2008.08.021
Tennant DA, Duran RV, Gottlieb E (2010) Targeting metabolic transformation for cancer therapy. Nat Rev Cancer 10:267–277. doi:10.1038/nrc2817
Warburg O (1956) On respiratory impairment in cancer cells. Science 124:269–270
DeBerardinis RJ, Cheng T (2010) Q’s next: the diverse functions of glutamine in metabolism, cell biology and cancer. Oncogene 29:313–324. doi:10.1038/onc.2009.358
Wise DR, Thompson CB (2010) Glutamine addiction: a new therapeutic target in cancer. Trends Biochem Sci 35:427–433. doi:10.1016/j.tibs.2010.05.003
Yuneva M, Zamboni N, Oefner P, Sachidanandam R, Lazebnik Y (2007) Deficiency in glutamine but not glucose induces MYC-dependent apoptosis in human cells. J Cell Biol 178:93–105. doi:10.1083/jcb.200703099
Wise DR, DeBerardinis RJ, Mancuso A, Sayed N, Zhang XY, Pfeiffer HK, Nissim I, Daikhin E, Yudkoff M, McMahon SB, Thompson CB (2008) Myc regulates a transcriptional program that stimulates mitochondrial glutaminolysis and leads to glutamine addiction. Proc Natl Acad Sci U S A 105:18782–18787. doi:10.1073/pnas.0810199105
Gao P, Tchernyshyov I, Chang TC, Lee YS, Kita K, Ochi T, Zeller KI, De Marzo AM, Van Eyk JE, Mendell JT, Dang CV (2009) c-Myc suppression of miR-23a/b enhances mitochondrial glutaminase expression and glutamine metabolism. Nature 458:762–765. doi:10.1038/nature07823
Yang C, Sudderth J, Dang T, Bachoo RM, McDonald JG, DeBerardinis RJ (2009) Glioblastoma cells require glutamate dehydrogenase to survive impairments of glucose metabolism or Akt signaling. Cancer Res 69:7986–7993. doi:10.1158/0008-5472.CAN-09-2266
van den Heuvel AP, Jing J, Wooster RF, Bachman KE (2012) Analysis of glutamine dependency in non-small cell lung cancer: GLS1 splice variant GAC is essential for cancer cell growth. Cancer Biol Ther 13:1185–1194. doi:10.4161/cbt.21348
Wang JB, Erickson JW, Fuji R, Ramachandran S, Gao P, Dinavahi R, Wilson KF, Ambrosio AL, Dias SM, Dang CV, Cerione RA (2010) Targeting mitochondrial glutaminase activity inhibits oncogenic transformation. Cancer Cell 18:207–219. doi:10.1016/j.ccr.2010.08.009
Kaadige MR, Looper RE, Kamalanaadhan S, Ayer DE (2009) Glutamine-dependent anapleurosis dictates glucose uptake and cell growth by regulating MondoA transcriptional activity. Proc Natl Acad Sci U S A 106:14878–14883. doi:10.1073/pnas.0901221106
Loi S, Sirtaine N, Piette F, Salgado R, Viale G, Van Eenoo F, Rouas G, Francis P, Crown JP, Hitre E, de Azambuja E, Quinaux E, Di Leo A, Michiels S, Piccart MJ, Sotiriou C (2013) Prognostic and predictive value of tumor-infiltrating lymphocytes in a phase III randomized adjuvant breast cancer trial in node-positive breast cancer comparing the addition of docetaxel to doxorubicin with doxorubicin-based chemotherapy: BIG 02-98. J Clin Oncol 31:860–867. doi:10.1200/JCO.2011.41.0902
Adams S, Gray RJ, Demaria S, Goldstein L, Perez EA, Shulman LN, Martino S, Wang M, Jones VE, Saphner TJ, Wolff AC, Wood WC, Davidson NE, Sledge GW, Sparano JA, Badve SS (2014) Prognostic value of tumor-infiltrating lymphocytes in triple-negative breast cancers from two phase III randomized adjuvant breast cancer trials: ECOG 2197 and ECOG 1199. J Clin Oncol. doi:10.1200/JCO.2013.55.0491
Ahn SG, Jeong J, Hong S, Jung WH (2015) Current issues and clinical evidence in tumor-infiltrating lymphocytes in breast cancer. J Pathol Transl Med 49:355–363. doi:10.4132/jptm.2015.07.29
Mockler MB, Conroy MJ, Lysaght J (2014) Targeting T cell immunometabolism for cancer immunotherapy; understanding the impact of the tumor microenvironment. Front Oncol 4:107. doi:10.3389/fonc.2014.00107
Chang CH, Qiu J, O’Sullivan D, Buck MD, Noguchi T, Curtis JD, Chen Q, Gindin M, Gubin MM, van der Windt GJ, Tonc E, Schreiber RD, Pearce EJ, Pearce EL (2015) Metabolic competition in the tumor microenvironment is a driver of cancer progression. Cell 162:1229–1241. doi:10.1016/j.cell.2015.08.016
Denkert C, Loibl S, Noske A, Roller M, Muller BM, Komor M, Budczies J, Darb-Esfahani S, Kronenwett R, Hanusch C, von Torne C, Weichert W, Engels K, Solbach C, Schrader I, Dietel M, von Minckwitz G (2010) Tumor-associated lymphocytes as an independent predictor of response to neoadjuvant chemotherapy in breast cancer. J Clin Oncol 28:105–113. doi:10.1200/JCO.2009.23.7370
Salgado R, Denkert C, Demaria S, Sirtaine N, Klauschen F, Pruneri G, Wienert S, Van den Eynden G, Baehner FL, Penault-Llorca F, Perez EA, Thompson EA, Symmans WF, Richardson AL, Brock J, Criscitiello C, Bailey H, Ignatiadis M, Floris G, Sparano J, Kos Z, Nielsen T, Rimm DL, Allison KH, Reis-Filho JS, Loibl S, Sotiriou C, Viale G, Badve S, Adams S, Willard-Gallo K, Loi S, International TWG (2015) The evaluation of tumor-infiltrating lymphocytes (TILs) in breast cancer: recommendations by an International TILs Working Group 2014. Ann Oncol 26:259–271. doi:10.1093/annonc/mdu450
Lakhani SREI, Schnitt SJ, Tan PH, van de Vijver MJ (eds) (2012) WHO classification of tumours of the breast. International Agency for Research on Cancer, Lyon
Lee HJ, Seo AN, Park SY, Kim JY, Park JY, Yu JH, Ahn JH, Gong G (2014) Low prognostic implication of fibroblast growth factor family activation in triple-negative breast cancer subsets. Ann Surg Oncol 21:1561–1568. doi:10.1245/s10434-013-3456-x
Hammond ME, Hayes DF, Wolff AC, Mangu PB, Temin S (2010) American Society of Clinical Oncology/College of American Pathologists guideline recommendations for immunohistochemical testing of estrogen and progesterone receptors in breast cancer. J Oncol Pract 6:195–197. doi:10.1200/JOP.777003
Wolff AC, Hammond ME, Hicks DG, Dowsett M, McShane LM, Allison KH, Allred DC, Bartlett JM, Bilous M, Fitzgibbons P, Hanna W, Jenkins RB, Mangu PB, Paik S, Perez EA, Press MF, Spears PA, Vance GH, Viale G, Hayes DF (2013) Recommendations for human epidermal growth factor receptor 2 testing in breast cancer: American Society of Clinical Oncology/College of American Pathologists Clinical Practice guideline update. Arch Pathol Lab Med. doi:10.5858/arpa.2013-0953-SA
Kim S, Kim Do H, Jung WH, Koo JS (2013) Expression of glutamine metabolism-related proteins according to molecular subtype of breast cancer. Endocr Relat Cancer 20:339–348. doi:10.1530/ERC-12-0398
Collins CL, Wasa M, Souba WW, Abcouwer SF (1997) Regulation of glutamine synthetase in human breast carcinoma cells and experimental tumors. Surgery 122:451–463 discussion 463-454
Lee HJ, Park IA, Song IH, Shin SJ, Kim JY, Yu JH, Gong G (2016) Tertiary lymphoid structures: prognostic significance and relationship with tumour-infiltrating lymphocytes in triple-negative breast cancer. J Clin Pathol 69:422–430. doi:10.1136/jclinpath-2015-203089
Briggs KJ, Koivunen P, Cao S, Backus KM, Olenchock BA, Patel H, Zhang Q, Signoretti S, Gerfen GJ, Richardson AL, Witkiewicz AK, Cravatt BF, Clardy J, Kaelin WG Jr (2016) Paracrine induction of HIF by glutamate in breast cancer: EglN1 senses cysteine. Cell 166:126–139. doi:10.1016/j.cell.2016.05.042
Chen X, Iliopoulos D, Zhang Q, Tang Q, Greenblatt MB, Hatziapostolou M, Lim E, Tam WL, Ni M, Chen Y, Mai J, Shen H, Hu DZ, Adoro S, Hu B, Song M, Tan C, Landis MD, Ferrari M, Shin SJ, Brown M, Chang JC, Liu XS, Glimcher LH (2014) XBP1 promotes triple-negative breast cancer by controlling the HIF1alpha pathway. Nature 508:103–107. doi:10.1038/nature13119
Chaturvedi P, Gilkes DM, Wong CC, Kshitiz LW, Zhang H, Wei H, Takano N, Schito L, Levchenko A, Semenza GL (2013) Hypoxia-inducible factor-dependent breast cancer-mesenchymal stem cell bidirectional signaling promotes metastasis. J Clin Invest 123:189–205. doi:10.1172/JCI64993
Sreekumar A, Poisson LM, Rajendiran TM, Khan AP, Cao Q, Yu J, Laxman B, Mehra R, Lonigro RJ, Li Y, Nyati MK, Ahsan A, Kalyana-Sundaram S, Han B, Cao X, Byun J, Omenn GS, Ghosh D, Pennathur S, Alexander DC, Berger A, Shuster JR, Wei JT, Varambally S, Beecher C, Chinnaiyan AM (2009) Metabolomic profiles delineate potential role for sarcosine in prostate cancer progression. Nature 457:910–914. doi:10.1038/nature07762
Vander Heiden MG (2011) Targeting cancer metabolism: a therapeutic window opens. Nat Rev Drug Discov 10:671–684. doi:10.1038/nrd3504
Parsons DW, Jones S, Zhang X, Lin JC, Leary RJ, Angenendt P, Mankoo P, Carter H, Siu IM, Gallia GL, Olivi A, McLendon R, Rasheed BA, Keir S, Nikolskaya T, Nikolsky Y, Busam DA, Tekleab H, Diaz LA Jr, Hartigan J, Smith DR, Strausberg RL, Marie SK, Shinjo SM, Yan H, Riggins GJ, Bigner DD, Karchin R, Papadopoulos N, Parmigiani G, Vogelstein B, Velculescu VE, Kinzler KW (2008) An integrated genomic analysis of human glioblastoma multiforme. Science 321:1807–1812. doi:10.1126/science.1164382
Dang L, White DW, Gross S, Bennett BD, Bittinger MA, Driggers EM, Fantin VR, Jang HG, Jin S, Keenan MC, Marks KM, Prins RM, Ward PS, Yen KE, Liau LM, Rabinowitz JD, Cantley LC, Thompson CB, Vander Heiden MG, Su SM (2009) Cancer-associated IDH1 mutations produce 2-hydroxyglutarate. Nature 462:739–744. doi:10.1038/nature08617
Ward PS, Patel J, Wise DR, Abdel-Wahab O, Bennett BD, Coller HA, Cross JR, Fantin VR, Hedvat CV, Perl AE, Rabinowitz JD, Carroll M, Su SM, Sharp KA, Levine RL, Thompson CB (2010) The common feature of leukemia-associated IDH1 and IDH2 mutations is a neomorphic enzyme activity converting alpha-ketoglutarate to 2-hydroxyglutarate. Cancer Cell 17:225–234. doi:10.1016/j.ccr.2010.01.020
Figueroa ME, Abdel-Wahab O, Lu C, Ward PS, Patel J, Shih A, Li Y, Bhagwat N, Vasanthakumar A, Fernandez HF, Tallman MS, Sun Z, Wolniak K, Peeters JK, Liu W, Choe SE, Fantin VR, Paietta E, Lowenberg B, Licht JD, Godley LA, Delwel R, Valk PJ, Thompson CB, Levine RL, Melnick A (2010) Leukemic IDH1 and IDH2 mutations result in a hypermethylation phenotype, disrupt TET2 function, and impair hematopoietic differentiation. Cancer Cell 18:553–567. doi:10.1016/j.ccr.2010.11.015
Swamy M, Pathak S, Grzes KM, Damerow S, Sinclair LV, van Aalten DM, Cantrell DA (2016) Glucose and glutamine fuel protein O-GlcNAcylation to control T cell self-renewal and malignancy. Nat Immunol 17:712–720. doi:10.1038/ni.3439
Acknowledgments
This research was supported by the Basic Science Research Program through the National Research Foundation of Korea (NRF) funded by the Ministry of Science, ICT & Future Planning (2015R1C1A1A02036484) and the Asan Institute for Life Sciences, Asan Medical Center, Seoul, Korea (2016-732).
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
This study was approved by the Institutional Review Board of Asan Medical Center and confirmed to the provisions of the Declaration of Helsinki.
Conflict of interest
The authors declare that they have no conflict of interest.
Additional information
Joo Young Kim and Sun-Hee Heo contributed equally to this article.
Rights and permissions
About this article

Cite this article
Kim, J.Y., Heo, SH., Choi, S.K. et al. Glutaminase expression is a poor prognostic factor in node-positive triple-negative breast cancer patients with a high level of tumor-infiltrating lymphocytes. Virchows Arch 470, 381–389 (2017). https://doi.org/10.1007/s00428-017-2083-5
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00428-017-2083-5