
Abstract
Human movements spontaneously entrain to auditory rhythms, which can help to stabilise movements in time and space. The properties of auditory rhythms supporting the occurrence of this phenomenon, however, remain largely unclear. Here, we investigate in two experiments the effects of pitch and tempo on spontaneous movement entrainment and stabilisation. We examined spontaneous entrainment of hand-held pendulum swinging in time with low-pitched (100 Hz) and high-pitched (1600 Hz) metronomes to test whether low pitch favours movement entrainment and stabilisation. To investigate whether stimulation and movement tempi moderate these effects of pitch, we manipulated (1) participants’ preferred movement tempo by varying pendulum mechanical constraints (Experiment 1) and (2) stimulation tempo, which was either equal to, or slightly slower or faster (± 10%) than the participant’s preferred movement tempo (Experiment 2). The results showed that participants’ movements spontaneously entrained to auditory rhythms, and that this effect was stronger with low-pitched rhythms independently of stimulation and movement tempi. Results also indicated that auditory rhythms can lead to increased movement amplitude and stabilisation of movement tempo and amplitude, particularly when low-pitched. However, stabilisation effects were found to depend on intrinsic movement variability. Auditory rhythms decreased movement variability of individuals with higher intrinsic variability but increased movement variability of individuals with lower intrinsic variability. These findings provide new insights into factors that influence auditory–motor entrainment and how they may be optimised to enhance movement efficiency.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
People spontaneously move in synchrony with auditory rhythms, musical rhythms in particular. Such auditory–motor entrainment happens intentionally but also often spontaneously (Burger, Thompson, Luck, Saarikallio, & Toiviainen, 2014; Demos, Chaffin, Begosh, Daniels, & Marsh, 2012; Néda, Ravasz, Brechet, Vicsek, & Barabási, 2000; Nozaradan, Petretz, & Keller, 2016; Phillips-Silver, Aktipis, & Bryant, 2010; Repp, 2005; Repp & Su, 2013; Torre, Varlet, & Marmelat, 2013; Van Dyck et al., 2015). The tendency for auditory rhythms to make us move has opened a window to understanding links between auditory and motor processes in the brain. This form of entrainment has also received interest in clinical contexts, as it can be used to stimulate and modulate the motor system of patients with movement disorders simply by presenting auditory rhythms (Hove & Keller, 2015; Malcom, Massie, & Thaut, 2009; McIntosh, Brown, Rice, & Thaut, 1997; Thaut et al., 1996; Thaut, McIntosh, & Rice, 1997). Auditory–motor entrainment can thus help to improve the locomotion of patients with Parkinson’s disease or stroke, for instance, by stabilising movements in time and space. However, not all auditory rhythms induce movement entrainment and stabilisation equally. In fact, it remains unclear which properties of auditory rhythms facilitate spontaneous motor entrainment. Here, we investigate in two experiments the mechanisms underlying spontaneous auditory–motor entrainment and movement stabilisation, and more specifically, the role played by the pitch and tempo of auditory rhythms.
Auditory rhythms induce movements and attract produced movements towards synchrony in a variety of contexts (Burger et al., 2014; Demos et al., 2012; Keller & Rieger, 2009; Peckel, Pozzo, & Bigand et al., 2014; Van Dyck et al., 2015). Underlying these effects, previous research has revealed shared functions and neural substrates between the auditory and motor systems, enabling mutual influence between auditory perception and movement production (Fujioka, Trainor, Larger, & Ross, 2012; Grahn & Brett, 2007; Repp & Knoblich, 2007; Zatorre, Chen, & Penhune, 2007). It has been suggested that spontaneous or unintentional movement synchronisation to auditory rhythms is governed by the dynamical entrainment processes of coupled oscillators (Demos et al., 2012; Kelso, 1995; Large, 2008, 2000; Néda et al., 2000). However, the investigation of spontaneous sensory–motor entrainment has so far been restricted mostly to studies involving visual rhythms, in particular, rhythmic movements of other people (Coey, Varlet, Richardson, 2012; Richardson, Marsh, Isenhower, Goodman, & Schmidt, 2007; Schmidt & Richardson, 2008; Schmidt & O’Brien, 1997; Varlet, Bucci, Richardson, & Schmidt, 2015). Spontaneous visuo-motor entrainment can occur when observing people walking or dancing, or when viewing simple oscillating stimuli on a computer screen, showing that motor and visual systems are also closely coupled.
These studies have demonstrated that the behavioural dynamics exhibited, which are characterised by spontaneous and intermittent synchronisation towards in-phase relations (movement proceeding in the same direction) and anti-phase relations (movement proceeding in opposite directions), can be explained by the well-established Haken–Kelso–Bunz coupled oscillator model (Coey, Varlet, Schmidt, & Richardson, 2011; Haken, Kelso, & Bunz, 1985; Richardson et al., 2007; Schmidt & O’Brien, 1997; Schmidt & Richardson, 2008; Varlet, Novembre, & Keller, 2017). As predicted by the model, these studies also revealed that the occurrence and strength of entrainment depend on the difference in tempo between the visual rhythm and an individual’s preferred movement (Fuchs, Jirsa, Haken, & Kelso, 1996; Lopresti-Goodman, Richardson, Silva, & Schmidt, 2008; Varlet, Coey, Schmidt, & Richardson, 2012). Entrainment is strongest when the tempo of the external rhythm is similar to the preferred movement tempo, decreases with increasing difference between the two tempi, and vanishes when the difference of tempo is too large (above around 15% difference) to be compensated by visuo-motor coupling (Lopresti-Goodman et al., 2008).
Studies involving the synchronisation of movements to sounds have revealed that auditory–motor entrainment is characterised by similar dynamics. For instance, Néda et al. (2000) showed that the hand clapping of an audience becomes spontaneously synchronised without the need for a conductor. Studies have also shown that the tempo difference between an individual’s movement and auditory rhythms may be important for stable auditory–motor synchronisation (Bardy, Hoffmann, Moens, Leman, & Dalla Bella, 2015; Repp, 2006; Styns, van Noorden, Moelants, & Leman 2007). Synchronisation when walking, running or cycling is most likely to occur when auditory rhythmic stimuli are presented at preferred movement tempo (Bardy et al., 2015; Leman et al., 2013; Styns et al., 2007). It has also been shown in piano duet studies that the coordination between two players is more stable when they have similar preferred movement tempo (Zamm, Pfordresher, & Palmer, 2015; Zamm, Wellman, & Palmer, 2016).
Previous research using systematic manipulations of the properties of visual rhythms has demonstrated that not all rhythms, even if they have the same tempo, are equal in inducing spontaneous entrainment (Varlet et al., 2012, 2014, 2015). The occurrence and strength of visuo-motor entrainment is modulated by the amplitude and velocity profiles of observed rhythmic movements, for instance (Varlet et al., 2012, 2014). As with visual rhythms, there is evidence that auditory rhythms, and musical rhythms specifically, do not all have the same impact on listeners’ movements (Burger, Thompson, Luck, Saarikallio, & Toivianinen, 2013; Leman et al., 2013; Leow, Parrott, & Grahn, 2014; Stupacher, Hove, Novembre, Schütz-Bosbach, & Keller, 2013). Recent studies have shown that ‘groove’ rhythms can modulate neurophysiological responses in a listener’s motor system (Stupacher et al., 2013) and can strongly impact upon an individual’s body sway dynamics (Ross, Warlaumont, Abney, Rigoli, & Balasubramaniam, 2016). There is evidence suggesting that the pitch (related to the fundamental frequency of the sound) may play a key role in such effects (Hove, Keller, & Krumhansl, 2007; Hove, Marie, Bruce, & Trainor, 2014; Van Dyck et al., 2013).
Recent studies have shown that musical groove is often associated with greater power fluctuation over time (spectral flux) in lower acoustic frequency bands (below 200 Hz), leading to greater effects on movements (Burger et al., 2013; Stupacher et al., 2016). It has been shown in finger-tapping tasks that movement is attracted more strongly to lower-pitched sounds than higher-pitched sounds when the two are presented together in separate streams (Hove et al., 2007; Hove et al., 2014; Keller & Repp, 2005). Van Dyck et al. (2013) found that dance movement became more synchronised to musical rhythms when the bass drum was amplified. Consistent with these findings, a recent study that examined synchronisation between paired individuals playing music together on custom-built user-friendly devices (electronic ‘music boxes’) also found that attraction and adaptation were stronger towards lower-pitched sounds (Novembre, Varlet, Muawiyath, Stevens, & Keller, 2015).
With regard to mechanisms, Hove et al. (2014) demonstrated facilitation of temporal processing for lower-pitched sounds at a neural level, as indicated by stronger brain responses to lower-pitched rhythms compared with higher-pitched rhythms, effects that may originate from the nonlinear dynamics of the human cochlear in the inner ear (Hove et al., 2014). It has also been proposed that lower-pitched rhythms may be effective at stimulating the vestibular system, which would contribute to greater impact on the listener’s auditory and motor systems (Phillips-Silver & Trainor, 2005, 2008; Todd & Lee, 2015; Todd, Rosengren, & Colebatch, 2008, 2009; Trainor, Gao, Lei, Lehtovaara, & Harris, 2009). Together, these results suggest that entrainment and stabilisation of human rhythmic movements could be strengthened by lower-pitched rhythms.
Research in the music domain suggests that the effects of pitch on auditory–motor entrainment might be moderated by the tempo of the auditory rhythms. In music, different dimensions of sound—including pitch, loudness, rhythm, and tempo—interact to convey musical meaning and affect via associations with physical space and bodily motion in the natural environment (Eitan & Granot, 2006; Scherer & Oshinsky, 1977; Zbikowsky, 2002). With regard to interactions specifically between pitch and tempo, high-pitched sequences are perceived to be faster, and are preferred to be played at faster tempi, than low-pitched sequences (Boltz, 2011; Collier & Hubbard, 2004; Tamir-Ostrover & Eitan, 2015). This tendency for lower-pitched sounds to be preferentially associated with slower rates while higher-pitched sounds are associated with faster rates is reflected in music performance conventions (Broze & Huron, 2013). For instance, the pulse or beat in music is most often carried in lower-pitched sounds, such as the bass and drums in a jazz rhythm section or an accompaniment in the left hand part of a piano piece (Large, 2000; Snyder & Krumhansl, 2001). These lower-pitched sounds typically proceed at slower rates (fewer events per time unit) than higher-pitched sounds in the melody, which raises the possibility that entrainment supported by lower-pitched sounds will occur preferentially for rhythms at slower tempi. Conversely, perceptual acuity and brain responses are greater for higher-pitched sounds than lower-pitched sounds when listening to melodies (Fujioka, Trainor, Large, & Ross, 2012; Marie & Trainor, 2013), suggesting that higher-pitched sounds might benefit auditory–motor entrainment at faster tempi.
Interactions between pitch and tempo may, furthermore, be related to the influence of biomechanical properties of the body on links between perception and action. Research investigating body movements induced by music has shown that fluctuations in lower-pitched sounds of music carrying the rhythmic pulse or beat entrain head movements but not hand movements, which could be explained by greater inertial load and thus slower preferred tempo of head movements (Burger et al., 2013). Conversely, this research also demonstrates that hand movements, which have lower inertial load and thus faster preferred tempi, may entrain more strongly with higher-pitched sounds. These findings suggest that the effects of pitch on auditory–motor entrainment might also be moderated by the biomechanical properties of the body segments being entrained, in particular, their preferred movement tempi.
The current study investigates these questions related to interactions between pitch and tempo in two experiments that examine the occurrence of spontaneous auditory–motor entrainment and movement stabilisation with various types of sound sequence. The task requires participants to listen to auditory metronomes with different pitches and tempi while swinging a handheld pendulum with different mechanical constraints to manipulate the individual’s preferred tempo. We hypothesized that low-pitched rhythms would induce stronger entrainment and movement stabilisation (reduced tempo and amplitude variability) than high-pitched rhythms in general. Nevertheless, specific effects of low- and high-pitched rhythms were also expected to be moderated by the preferred tempo of the participant’s movement (i.e., slow or fast, Experiment 1) and/or whether the rhythm was presented at slightly slower or faster tempo (Experiment 2).
Moreover, we also examined across the two experiments the influence of individual intrinsic movement variability on the occurrence of movement period and amplitude stabilisation. As detailed above, movement stabilisation induced by entrainment is receiving growing interest in the literature as a means to improve motor performances (Hove & Keller, 2015), but conditions required for an individual to benefit from the presentation of auditory rhythms remain largely unclear. Indeed, increased movement stability induced by entrainment or synchronisation has been shown so far predominantly in patients suffering from motor disorders (Malcom et al., 2009; McIntosh et al., 1997; Thaut et al., 1996), that is, in individuals who exhibit relatively large tempo and amplitude variability. However, benefits of auditory rhythms on individuals’ movement with lower intrinsic variability, such as athletes for instance, might be more mixed. It is possible that the occurrence of entrainment for movements that are intrinsically more stable, in particular, for stimuli slightly slower or faster than an individual’s preferred tempo, may lead to less regular movements with higher variability in time and space compared to conditions without external rhythmic stimulation. We, therefore, expected that the effects of auditory rhythms on movement variability would interact with participant’s intrinsic movement variability in such a way that stabilisation would preferentially occur for movements intrinsically more variable, whereas weaker stabilisation effects, and potentially even destabilisation effects, would occur for movements intrinsically less variable.
Experiment 1
Experiment 1 investigated entrainment and movement stabilisation in participants swinging a handheld pendulum with either high or low inertial load while listening to either low-pitched (i.e., 100 Hz) or high-pitched (i.e., 1600 Hz) metronome presented at each individual’s preferred movement tempo. Previous research has shown that a pendulum’s inertial load changes an individual’s preferred movement tempo such as high inertial load leads to slower preferred tempo and low inertial load leads to faster preferred tempo (Coey et al., 2011; Kugler & Turvey, 1987; Schmidt & O’Brien, 1997). Although stronger entrainment was expected with the lower-pitched metronome in general, enhanced entrainment of the low inertial pendulum swinging (faster tempo) with high-pitched metronome was considered possible. Movement stabilisation (decreased variability in tempo and amplitude) compared to conditions without auditory stimulation, was also expected to be stronger for low-pitched metronome in general, though variations depending on the pendulum swung by participants and individual participant’s intrinsic movement variability were also deemed possible.
Methods
Participants
Twenty-three psychology undergraduates and one staff member (N = 24) from Western Sydney University volunteered to participate in the experiment (18 females and 6 males aged from 18 to 36 years; M = 22.13, SD = 4.57). Our sample size was chosen based on an a priori power analysis to detect medium effect sizes (f = 0.25) with at least 80% power, in line with effect sizes previously reported in sensorimotor entrainment studies (e.g., Romero, Coey, Schmidt, & Richardson, 2012; Varlet et al., 2012). The undergraduates received credit in partial fulfilment of course requirements. All 24 participants were right-handed, had normal hearing and normal or corrected-to-normal vision. All participants provided written informed consent prior to the experiment, which was approved by the Western Sydney University Ethics Committee.
Apparatus
A chair was positioned in front of a 22-inch BenQ computer monitor that was used to display visual stimuli for a letter detection task (described below). The chair had a custom support on the right arm on which the participant’s forearm was positioned to perform the pendulum-swinging task.
Auditory rhythmic stimuli were generated in Praat software (Boersma & Weenink, 2014) and corresponded to pure sine tones with either 100 or 1600 Hz pitch. The duration of the tones was 150 ms including a 10 ms fade in and fade out. The tones were equalised for perceived loudness following the Cambridge loudness model (Moore, Glasberg, & Baer, 1997) and presented via Sennheiser HD 280 pro headphones (Sennheiser, Wedemark, Germany) at a comfortable intensity held constant for all participants (91.4 dB for 100 Hz and 89.6 dB for 1600 Hz).
Two different wooden pendulums were used for the pendulum-swinging task. A pendulum with low inertial load had a length of 45 cm and a 50 g mass attached at its base, resulting in an eigenfrequency of 1 Hz. A pendulum with high inertial load had a length of 48 cm and a 225 g mass attached at its base resulting in an eigenfrequency of 0.9 Hz. Previous research has shown that, in pendulum-swinging tasks, participants spontaneously adopt a preferred tempo very close to the eigenfrequency of the pendulum, and that this is true for a wide range of pendulum eigenfrequencies when manipulated by modifying pendulum’s length and weight (Kugler & Turvey, 1987; Schmidt & Turvey, 1992; Schmidt & O’Brien, 1997). A 1 × 1 × 1.5 cm motion-tracking sensor was attached to each pendulum to record the oscillations produced by the participant at a sampling rate of 240 Hz with 0.01 mm spatial resolution via a Polhemus LIBERTY motion tracker (Polhemus Ltd., VT, USA). Auditory stimuli presented to the participant were recorded together with movement data on a PC computer for off-line analyses.
To avoid intentional synchronisation and visuomotor interference due to eye movements, a letter detection concentration task was presented to participants on the monitor (Varlet et al., 2015). A fixation cross was displayed at the centre of the monitor throughout each trial, alternating with letters occurring briefly for 0.67 s at random time intervals between 4 and 18 s, which the participant had to read aloud.
Procedure
On arrival, the participant was provided with an information sheet that described the task as a letter detection task with auditory and movement perturbations, requiring her/him to swing a hand-held pendulum at her/his preferred tempo while reading aloud letters that flashed at random times at the centre of the monitor. This cover story was intended to ensure that the synchronisation of participants’ movements was spontaneous. Written informed consent was then invited (and in all cases obtained).
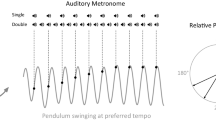
The participant, who was seated in the chair in front of the monitor, was then asked to swing the hand-held pendulum parallel to the sagittal plane from the right wrist joint (ulnar–radial flexion–extension), as in previous visuomotor synchronisation studies (see Fig. 1; Lopresti-Goodman et al., 2008; Varlet et al., 2015). The participant was instructed to practice swinging each pendulum until a comfortable preferred movement tempo was established. The participant’s preferred movement tempo was then recorded for each pendulum in a counterbalanced order in three trials of 60 movement cycles in the absence of auditory stimulation. Instructions for the task were displayed in full view on the computer monitor. The task instructed the participant to swing the nominated pendulum at the preferred movement tempo, while reading out the letters that appeared at the centre of the screen.

Illustration of the pendulum-swinging task performed by participants in Experiment 1 and 2. The top panel illustrates the pendulum swinging produced by a representative participant over 30 s and the bottom panel represents the corresponding relative phase angles (between − 180° and 180°). The black dots illustrate the auditory metronome presented
The participant was then provided with headphones to present auditory rhythmic stimuli before experimental trials were started. Two independent variables, Stimulus Pitch [low-pitch (100 Hz), high-pitch (1600 Hz), and control (mute)], and Pendulum (Slow and Fast) were manipulated across 30 trials, representing five blocks of six trials with one trial for each of the six conditions: slow/low-pitch, slow/high-pitch, slow/control, fast/low-pitch, fast/high-pitch, and fast/control. In each block, the first three trials were performed with one pendulum and the last three trials with the other pendulum in a counterbalanced order, while the order of the different stimulus conditions was randomized. Each high-pitch and low-pitch trial consisted of 8 s without auditory stimulus followed by 45 cycles of the auditory stimulus, presented at the preferred movement tempo of the participant. Each control trial was the same duration, 8 s followed by 45 cycles at the participant’s preferred tempo, but without any auditory stimulus presented. The experimenter waited for the participant to begin swinging the given pendulum at her/his preferred movement tempo and then initiated each trial (which started with 8 s of silence) manually at a random phase of the participant’s oscillation. Short breaks between trials were taken as required.
Each participant completed all trials in one 1-h session. On completion of the trials, each participant was asked to complete a questionnaire to obtain demographic information before being debriefed on the purposes of the study prior to leaving.
Design and analysis
The first 10 s of each trial were discarded to eliminate transient fluctuations in the movement. The movement time series data was centered around zero and filtered using a bidirectional Butterworth band pass filter with 0.1 and 15 Hz cut-off frequencies to remove slow trends and high frequency noise (Varlet et al., 2012; Varlet, Schmidt, & Richardson, 2016). One trial for each of two participants was removed from the analyses because the incorrect pendulum was used.
The length of the mean resultant vector of the relative phase angles between the participant’s movement and stimulus onsets was computed using circular statistics to obtain the degree of synchrony, with 0 corresponding to no synchrony (no phase locking) and 1 corresponding to complete synchrony (perfect phase locking; Batschelet, 1981; Pikovsky et al., 2003, Varlet et al., 2016). The relative phase angles capture the relation between the participant’s movement and the metronome, and the length of the mean resultant vector indicates how consistent this relation is. If relative phase angles tend to remain the same, then all vectors tend to go in the same direction, indicating consistent phase relation and the occurrence of entrainment. With spontaneous entrainment, synchrony is only intermittent and characterised by nonstationary relative phase angle time series with continuous drifts, and therefore, mean and standard deviation measures are limited and the strength of entrainment is better indexed using the mean resultant vector (Pikovsky et al., 2003). The relative phase angles were obtained by computing the continuous phase of participants’ movements using a Hilbert Transform and extracting phase values at stimulus onsets. The phase angles extracted were artefact-free, as the first and last stimulus onsets were sufficiently distant from the beginning and end of the continuous phase calculated with the Hilbert transform. The phase synchrony in the control trials was calculated in the same manner except that auditory stimuli were mute (hence virtual time points were used to represent stimulus onsets). The ‘stimulus onsets’ for the mute metronome condition were, therefore, the same as those for the 100 and 1600 Hz metronome conditions, which allowed the level of incidental synchrony that could occur per chance to be computed. Significantly higher synchrony in 100 and 1600 Hz conditions than this control synchrony indicates the occurrence of more consistent phase relations, and thus, entrainment. Mean resultant vector values were standardized using a Fisher Transformation before statistical analyses.
The time between the points of maximum angular extension, as defined by the maxima of the movement time series, was also computed to determine the average of the participant’s preferred movement period (tempo), and the corresponding coefficient of variation (CV = SD/mean × 100), indexing the magnitude of the tempo variability inherent to her/his movement. The distance between maximum angular flexion and extension of each cycle, as defined by the difference between two consecutive minima and maxima of the movement time series, was used to compute the average movement amplitude and the corresponding coefficient of variation. Movement period and amplitude were computed based on turning points in movement time series for consistency with previous research (Schmidt, Richardson, Arsenault, & Galantucci, 2007; Varlet et al., 2012, 2016). The average and CV of each participant’s movement period and amplitude were computed within each trial and then averaged across the trials.
Repeated-measures analyses of variance (ANOVAs) were conducted on each of the five dependent variables—Movement Synchrony, Movement Period (Mean and CV) and Movement Amplitude (Mean and CV) with factors of Stimulus Pitch [Low-pitch (100 Hz), High-pitch (1600 Hz), and Control (mute)], and Pendulum (Slow and Fast). The Greenhouse–Geisser correction was used when conditions of sphericity were not met. Post-hoc analyses were conducted using Bonferroni post hoc tests.
To examine whether the effects of auditory stimulation on participants’ movement variability depended on her/his intrinsic level of variability, we computed stabilisation index values corresponding to the CV values in the Low-pitch and High-pitch conditions after subtraction of the control (mute) condition. Negative values indicate the occurrence of stabilisation and positive values indicate the occurrence of destabilisation compared to the control condition. These stabilisation index values were computed for both movement period and amplitude (CVs) and then submitted to multiple linear regression analyses with the independent variables Intrinsic Variability (i.e., CV in control condition), Stimulus Pitch [Low-pitch (100 Hz), High-pitch (1600 Hz)], and Intrinsic Variability × Stimulus Pitch interaction term. Linear regression analyses were conducted separately on each pendulum, as the participant’s intrinsic variability differed depending on the pendulum swung.
Results
Movement synchrony
The ANOVA performed on the movement synchrony yielded a significant main effect of Stimulus Pitch, F(2, 46) = 7.69, MSE = 0.02, p = 0.004, ηg2 = 0.03 and no significant effect of Pendulum, F(1, 23) = 0.05, MSE = 0.04, p = 0.83, ηg2 < 0.001, and interaction between those two factors, F(2, 46) = 0.97, MSE = 0.01, p = 0.37, ηg2 = 0.003. Post hoc tests indicated that synchrony in the low-pitch condition was significantly higher than the control condition, t(46) = 3.86, p = 0.001, d = 0.45, and the high-pitch condition, t(46) = 2.53, p = 0.04, d = 0.26 (see Fig. 2). No significant difference was found between the high-pitch and control conditions, t(46) = 1.33, p = 0.57, d = 0.18, showing that entrainment preferentially occurred with low-pitch stimuli.

Movement synchrony as a function of the different stimulus pitch conditions [i.e., low-pitch (100 Hz), high-pitch (1600 Hz) and Mute] in Experiment 1. The error bars represent the standard error of the mean (O’Brien & Cousineau, 2014)
Movement period
The ANOVA performed on the mean movement period of participants revealed a significant main effect of Pendulum, F(1, 23) = 260.82, MSE = 0.002, p < 0.0001, ηg2 = 0.21, showing that, as expected, participants’ movement tempo was slower with greater inertial load (i.e., Slow Pendulum). Participants’ movement period was equal to 1.10 s (SD = 0.12) with the Slow Pendulum and to 0.97 s (SD = 0.13) with the Fast Pendulum. In line with previous research, participants’ preferred frequency was very close to the pendulum’s eigenfrequency − 0.91 Hz for the Slow Pendulum that had an eigenfrequency of 0.9 Hz and 1.03 Hz for the Fast Pendulum that had an eigenfrequency of 1 Hz (Kugler & Turvey, 1987; Schmidt & Turvey, 1992; Schmidt & O’Brien, 1997). The ANOVA did not reveal significant main effect of Stimulus Pitch, F(2, 46) = 0.68, MSE < 0.001, p = 0.49, ηg2 < 0.001, or interaction between Stimulus Pitch and Pendulum, F(2, 46) = 0.40, MSE < 0.001, p = 0.64, ηg2 < 0.001.
The ANOVA performed on the CV of participants’ movement period also only indicated a significant main effect of Pendulum, F(1, 23) = 76.44, MSE = 0.032, p < 0.0001, ηg2 = 0.18, showing that participants’ timing variability was greater with the Fast Pendulum (M = 3.32%, SD = 0.99) compared with the Slow Pendulum (M = 2.49%, SD = 0.77). No significant main effect of Stimulus Pitch, F(2, 46) = 0.88, MSE = 0.12, p = 0.40, ηg2 = 0.002, or interaction between Stimulus Pitch and Pendulum were found, F(2, 46) = 1.81, MSE = 0.14, p = 0.18, ηg2 = 0.004.
The multiple linear regression analysis conducted on the period stabilisation index values for the Slow Pendulum indicated that the model with the predictors Intrinsic Variability, Stimulus Pitch, and Intrinsic Variability × Stimulus Pitch did not reach significance, F(3, 44) = 2.39, p = 0.08, R2 = 0.14, and that none of the predictors tested were significant (all p values > 0.05). When the interaction term was removed, the model reached significance, F(2, 45) = 3.56, p = 0.037, R2 = 0.14, and Intrinsic Variability became a significant predictor, t(45) = − 2.31, p = 0.025. The multiple linear regression analysis conducted on the period stabilisation index values for the Fast Pendulum revealed that the model with Intrinsic Variability, Stimulus Pitch, and Intrinsic Variability × Stimulus Pitch was significant, F(3, 44) = 15.55, p < 0.0001, R2 = 0.51, and that Intrinsic Variability was a significant predictor, t(44) = − 5.20, p < 0.0001. These results show that tempo variability with both Slow Pendulum and Fast Pendulum decreased for participants with high intrinsic variability but increased for participants with low intrinsic variability as seen in Fig. 3.

Movement period and amplitude stabilisation index values as a function of individual participant’s intrinsic movement variability (CV exhibited in the control condition without auditory stimulus) and the type of pendulum swung in Experiment 1. Negative values indicate movement stabilisation and positive values indicate movement destabilisation. Lines of best fit are included when a significant effect of the covariate Intrinsic Variability was found. Separate lines for low-pitch (100 Hz) and high-pitch (1600 Hz) conditions are shown when the interaction between Intrinsic Variability and Stimulus Pitch was significant
Movement amplitude
The ANOVA performed on the amplitude of participants’ movements did not show significant effects of Pendulum, F(1, 23) = 1.68, MSE = 8.39, p = 0.21, ηg2 < 0.001, Stimulus Pitch, F(2, 46) = 0.09, MSE = 2.61, p = 0.86, ηg2 < 0.001, or the interaction between those two factors, F(2, 46) = 1.34, MSE = 1.62, p = 0.27, ηg2 < 0.001.
The ANOVA performed on the CV of movement amplitude indicated a significant main effect of Pendulum, F(1, 23) = 46.76, p < 0.0001, MSE = 1.31, ηg2 = 0.04. Greater amplitude variability was exhibited with the Fast Pendulum (M = 9.48%, SD = 3.31) compared with the Slow Pendulum (M = 8.17%, SD = 3.06). No significant main effect of Stimulus Pitch, F(2, 46) = 0.92, MSE = 1.42, p = 0.40, ηg2 = 0.002, or interaction between Pendulum and Stimulus Pitch, F(2, 46) = 1.01, MSE = 2.17, p = 0.36, ηg2 = 0.002, were revealed.
The multiple linear regression analyses on the amplitude stabilisation index values for the Slow Pendulum indicated that the model with the predictors Intrinsic Variability, Stimulus Pitch, and Intrinsic Variability × Stimulus Pitch was significant, F(3, 44) = 16.61, p < 0.0001, R2 = 0.53, and that Intrinsic Variability, t(44) = − 6.57, p < 0.0001; Stimulus Pitch, t(44) = − 2.75, p = 0.009; and Intrinsic Variability × Stimulus Pitch, t(44) = 2.84, p = 0.007, were all significant predictors. As seen in Fig. 3, these results show that the amplitude variability decreased for participants with high intrinsic variability but increased for participants with low intrinsic variability, and that these effects were stronger when low-pitched stimuli were presented. The multiple linear regression analyses conducted on the amplitude stabilisation index values for the Fast Pendulum yielded no significant models and predictors whether or not the interaction term was included (all p values > 0.05).
Discussion
Experiment 1 investigated the effects of pitch in auditory rhythms on the occurrence of auditory–motor entrainment and movement stability, and whether these effects are modulated by mechanical constraints that influence the preferred tempo of movements. Participants swung a handheld pendulum with either high (inducing slower tempo) or low inertial load (inducing faster tempo) while listening to either low-pitched (i.e., 100 Hz) or high-pitched (i.e., 1600 Hz) metronome presented at their preferred movement tempo.
In line with previous research that investigated pendulum-swinging entrainment with visual rhythms, the results indicated that movement synchrony in the presence of auditory metronomes was greater than incidental synchrony occurring in mute control trials without auditory stimulation (Richardson et al., 2007; Schmidt & O’Brien, 1997; Varlet et al., 2016). This finding demonstrates that although participants were only instructed to maintain their preferred movement tempo, they spontaneously entrained to the metronome. The synchrony values of around 0.3 exhibited in Experiment 1 corroborate values previously reported in visuo-motor entrainment studies, showing that movement synchrony was only intermittent rather than continuously phase locked, as is typically observed in spontaneous synchronisation situations (Repp, 2005, 2006; Richardson et al., 2007; Varlet et al., 2014, 2016).
The results also indicated that movement synchronisation was stronger for low-pitched rhythms compared to high-pitched rhythms. These findings are in line with our predictions and previous research that suggested stronger influence of low-pitched rhythms on an individual’s motor system (Hove et al., 2007; Hove et al., 2014; Novembre et al., 2015; Todd & Lee, 2015). It is particularly noteworthy that the presentation of an auditory metronome altered the stability of participants’ movement tempo and amplitude, and that these changes appeared stronger with low-pitched rhythms. The results also demonstrated that the direction of these changes depended on individual differences in the intrinsic variability of participants’ movement. Movement variability decreased for participants with higher intrinsic movement variability, whereas movement variability increased for participants with lower intrinsic movement variability. This new finding is noteworthy as it shows that the effects of auditory rhythm on an individual’s movement stability are not necessarily beneficial and that analyses at the group level might not reflect the true effects of a particular type of stimulation on movement stability.
For movement tempo (period) variability, these effects occurred when swinging both pendulums. However, for movement amplitude variability, the effects only occurred when swinging the pendulum associated with intrinsically more stable movements due to high inertial load. Furthermore, for movement amplitude variability, these effects were found to be stronger with low-pitched rhythms compared to high-pitched rhythms, showing that low-pitch rhythms do not only tend to facilitate entrainment but also the magnitude of (de)stabilisation effects. This specificity could be due to greater difference between low and high intrinsic variability participants when using this pendulum, although further exploration is necessary in the future to better understand these differences and their links with mechanical constraints on movement.
Experiment 1 demonstrates that the presentation of auditory rhythms leads to auditory–motor entrainment and modifies movement stability, and that these effects are strengthened by low-pitched rhythms compared to high-pitched rhythms. However, the effects of low-pitched rhythms occurred equally strongly for the pendulums with high and low inertial loads, which does not support the hypothesis that the effects on the movement of large and small body segments, characterised by slower and faster preferred tempo, would be preferentially facilitated by low and high-pitched rhythms, respectively. Although an interaction between pitch and tempo was not found in Experiment 1, such an interaction may be possible with a wider range of tempi.
Indeed, auditory rhythms were only presented in Experiment 1 at participants’ preferred tempi, while entrainment in everyday situations, including for clinical purposes, often occurs with rhythms at slower or faster tempo (Bardy et al., 2015; Burger et al., 2013; Hove & Keller, 2015; Leman et al., 2013). It is possible that entrainment to slower and faster rhythms may be facilitated by different pitches, due to natural preference for slower rhythms when low-pitched and faster rhythms when high-pitched. Experiment 2 aimed to determine whether entrainment to slower rhythms is stronger for low-pitched sequences and entrainment to faster rhythms is stronger for high-pitched rhythms.
Experiment 2
Experiment 2 investigated the entrainment and movement stabilisation while participants swung a handheld pendulum and listened to either a low-pitched (i.e., 100 Hz) or a high-pitched (i.e., 1600 Hz) metronome, which was presented either at the participant’s preferred movement tempo or preferred tempo ± 10%. It was expected that movement entrainment and stabilisation would be stronger with the low-pitched metronome and at the participant’s preferred movement tempo. It was also expected that movement entrainment to the faster metronome would be selectively facilitated by high-pitched metronome while entrainment to slower metronome would be facilitated by low-pitched metronome, given preferred pitch–tempo relations in everyday auditory rhythms, musical rhythms in particular.
Methods
Participants
Twenty-three Western Sydney University psychology undergraduates and one staff member (N = 24; all different from Experiment 1) volunteered to participate in the experiment (16 females and 8 males aged from 18 to 30 years; M = 20.33, SD = 2.84). Our sample size was chosen based on an a priori power analysis to detect medium effect sizes (f = 0.25) with at least 80% power, in line with effect sizes reported in Experiment 1 and previous sensorimotor entrainment studies (e.g., Romero, Coey, Schmidt, & Richardson, 2012; Varlet et al., 2012). The undergraduates received credit to partially fulfil course requirements. All 24 participants were right-handed, had normal hearing and normal or corrected-to-normal vision. All participants provided written informed consent prior to the experiment, which was approved by the Western Sydney University Ethics Committee.
Apparatus
The apparatus was the same as in Experiment 1 with the exception that only one intermediate inertial load pendulum with a length of 45 cm and a 75 g mass attached at its base, resulting in an eigenfrequency of 0.94 Hz, was used.
Procedure
The procedure was the same as in Experiment 1 with the exception that only one pendulum was used and metronome stimuli were presented at the participant’s preferred movement tempo and also at his or her preferred tempo ± 10%. Two independent variables, Stimulus Pitch [low-pitch (100 Hz), high-pitch (1600 Hz), and control (mute)], and Stimulus Tempo (− 10%, Preferred and + 10%) were manipulated across 35 trials, representing seven conditions: − 10%/low-pitch, Preferred/low-pitch, + 10%/low-pitch, − 10%/high-pitch, Preferred/high-pitch, + 10%/high-pitch, and control/mute. The same control/mute trials were used to compute the level of incidental movement synchrony that could be expected to occur due to chance in each of the six other conditions. The order of the trials was randomized in five blocks of seven trials, with each block containing one trial of each condition.
Design and analysis
As in Experiment 1, repeated-measures ANOVAs were conducted on each of the five dependent variables computed—Movement Synchrony, Movement Period (Mean and CV) and Movement Amplitude (Mean and CV)—with factors of Stimulus Pitch [Low-pitch (100 Hz), High-pitch (1600 Hz), and Control (mute)], and Stimulus Tempo (− 10%, Preferred and + 10%). Multiple linear regression analyses were also conducted on movement period and amplitude stabilisation index values with the independent variables Intrinsic Variability [Control (mute)], Stimulus Pitch [Low-pitch (100 Hz) and High-pitch (1600 Hz)], Stimulus Tempo (− 10%, Preferred and + 10%), and all corresponding 2-way and 3-way interaction terms to examine whether the effects of properties of the metronome on movement variability depended on participants’ intrinsic level of variability.
Results
The analyses revealed that one participant continuously synchronised with the stimulus (synchrony values very close to 1 throughout the experimental trials) and one participant did not consistently swing the pendulum throughout the trials. Both were, therefore, removed, leaving twenty-two participants in the dataset.
Movement synchrony
The ANOVA performed on movement synchrony revealed a significant main effect of Stimulus Pitch, F(2, 42) = 6.15, MSE = 0.01, p = 0.005, ηg2 = 0.02. Post-hoc comparisons indicated that compared with the control condition, synchrony was significantly higher in the low-pitch condition, t(42) = 3.39, p = 0.0045, d = 0.26, but not in the high-pitch condition, t(42) = 2.46, p = 0.054, d = 0.19, and no significant differences between the high-pitch and low-pitch conditions, t(42) = 0.93, p = 1, d = 0.07. Note that the effect of high-pitch was close to reaching significance, suggesting that some (perhaps intermittent) entrainment may have also occurred despite appearing weaker than in the low-pitch condition. The ANOVA also indicated a significant main effect of Stimulus Tempo, F(2, 42) = 17.12, MSE = 0.08, p < 0.0001, ηg2 = 0.35 but no interaction between Stimulus Tempo and Stimulus Pitch, F(4, 84) = 0.73, MSE = 0.01, p = 0.53, ηg2 = 0.005, showing that entrainment occurred for all stimulation tempi although the differences between low-pitch and high-pitch conditions and the control condition appear to be greater for stimuli presented at preferred and faster tempi (see Fig. 4).

Movement synchrony as a function of the different stimulus tempo and pitch conditions [i.e., low-pitch (100 Hz), high-pitch (1600 Hz) and Mute] in Experiment 2. The error bars represent the standard error of the mean (O’Brien & Cousineau, 2014)
Movement period
Mean movement period across participants was 1.08 s (SD = 0.09), corresponding to a preferred frequency of 0.93 Hz, which was close to the pendulum’s eigenfrequency of 0.94 Hz in accordance with previous studies (Kugler & Turvey, 1987; Schmidt & Turvey, 1992; Schmidt & O’Brien, 1997). The ANOVA performed on movement period indicated no main effect of Stimulus Pitch, F(2, 42) = 0.35, MSE = 0.002, p = 0.64, ηg2 < 0.001, Stimulus Tempo, F(2, 42) = 2.98, MSE < 0.001, p = 0.062, ηg2 < 0.001, or interaction between those two factors, F(4, 84) = 0.97, MSE < 0.001, p = 0.40, ηg2 < 0.001. Note that the main effect of Stimulus Tempo was close to significance, suggesting that participants’ movement period tended to become shorter with faster stimulation tempi and longer with slower stimulation tempi. Mean period was 1.082 s (SD = 0.09) for fast, 1.087 s (SD = 0.09) for preferred, and 1.089 s (SD = 0.10) for slow stimulation frequencies.
The ANOVA performed on the CV of movement period did not indicate significant main effects of Stimulus Pitch, F(2, 42) = 0.55, MSE = 0.22, p = 0.55, ηg2 = 0.001, Stimulus Tempo, F(2, 42) = 1.78, MSE = 0.18, p = 0.19, ηg2 = 0.003, or the interaction between Stimulus Tempo and Stimulus Pitch, F(4, 84) = 2.04, MSE = 0.14, p = 0.11, ηg2 = 0.004.
The multiple linear regression analyses on the period stabilisation index values indicated that the model was significant when all predictors were included, F(11, 120) = 2.78, p = 0.003, R2 = 0.20, but that none of the predictors were significant (all p values > 0.05). When the 3-way interaction term Intrinsic Variability × Stimulus Pitch × Stimulus Tempo was removed, the model remained significant, F(7, 124) = 3.75, p = 0.001, R2 = 0.17, the predictor Intrinsic Variability became significant, t(124) = − 2.28, p = 0.025, and there was a significant interaction between Intrinsic Variability and whether or not the metronome was presented at slower tempo − 10%, t(124) = − 2.65, p = 0.009. Simple linear regression analyses conducted on each of the stimulus tempo with the independent variable Intrinsic Variability confirmed this interaction. Regressions were significant for Preferred, F(1, 42) = 11.71, p = 0.001, R2 = 0.22, and for + 10%, F(1, 42) = 10.56, p = 0.002, R2 = 0.20, but not for − 10%, F(1, 42) = 0.07, p = 0.80, R2 = 0.002, showing that metronomes led to increased tempo variability for participants who were intrinsically more stable and decreased variability for participants who were intrinsically less stable when they were presented at participant’s preferred tempo or slightly faster, as seen in Fig. 5. No other significant effects were found (all p values > 0.05).

Movement period and amplitude stabilisation index values as a function of individual participant’s intrinsic movement variability (CV exhibited in the control condition without auditory stimulus) in Experiment 2. Negative values indicate movement stabilisation and positive values indicate movement destabilisation. Lines of best fit are included when a significant effect of the covariate Intrinsic Variability was found. Separate lines for the different tempo conditions are shown when the interaction between Intrinsic Variability and Stimulus Tempo was significant
Movement amplitude
The ANOVA performed on the amplitude of participants’ movements yielded a significant main effect of Stimulus Pitch, F(2, 42) = 7.27, MSE = 3.51, p = 0.003, ηg2 = 0.002, showing that movement amplitude increased with auditory stimulation compared to control trials. Post-hoc comparisons indicated significant differences between control (M = 28.32, SD = 11.30) and high-pitch (M = 29.18, SD = 11.39), t(42) = 2.79, p = 0.02, d = 0.08, and low-pitch (M = 29.44, SD = 11.97), t(42) = 3.65, p = 0.002, d = 0.10, and no significant difference between high-pitch and low-pitch, t(42) = 0.85, p = 1, d = 0.02. The ANOVA also yielded a main effect of Stimulus Tempo close to significance, F(2, 42) = 3.38, MSE = 2.21, p = 0.06, ηg2 < 0.001, suggesting that participants’ movement amplitude tended to decrease with faster stimulation tempi. Post-hoc comparisons indicated differences between the slow stimulation tempo (M = 29.31, SD = 11.73) and preferred tempo (M = 28.80, SD = 11.50), t(42) = − 2.29, p = 0.08, d = 0.04, and faster tempo (M = 28.82, SD = 11.42), t(42) = − 2.21, p = 0.10, d = 0.04, close to significance. The ANOVA did not indicate a significant interaction between Stimulus Pitch and Stimulus Tempo, F(4, 84) = 0.88, MSE = 2.29, p = 0.45, ηg2 < 0.001.
The ANOVA performed on the CV of participants’ movement amplitudes revealed a significant main effect of Stimulus Pitch, F(2, 42) = 5.59, MSE = 2.37, p = 0.01, ηg2 = 0.01, showing that movement amplitude variability decreased with auditory stimulation, especially low-pitched (see Fig. 6). Post-hoc comparisons indicated a significant difference between low-pitch and control [t(42) = − 3.21, p = 0.0075, d = 0.24] and a difference between high-pitch and control close to significance [t(42) = − 2.40, p = 0.062, d = 0.17]. The ANOVA revealed no significant main effect of Stimulus Tempo, F(2, 42) = 0.84, MSE = 1.21, p = 0.43, ηg2 < 0.001, and no interaction between Stimulus Pitch and Stimulus Tempo, F(4, 84) = 0.20, MSE = 1.35, p = 0.90, ηg2 < 0.001.

Movement amplitude variability as a function of the different stimulus pitch conditions [i.e., low-pitch (100 Hz), high-pitch (1600 Hz) and Mute] in Experiment 2. The error bars represent the standard error of the mean (O’Brien & Cousineau, 2014)
The multiple linear regression analyses conducted on the amplitude stabilisation index values indicated that the model with Intrinsic Variability, Stimulus Pitch, Stimulus Tempo, and all corresponding interaction terms was significant, F(11, 120) = 3.86, p < 0.0001, R2 = 0.26, and that only the predictor Intrinsic Variability was significant, t(120) = − 3.70, p = 0.0003. The same results were found when the 3-way interaction term was removed from the analysis and when the 3-way and 2-way interaction terms were removed from the analyses. This shows that amplitude variability of participants who were intrinsically more variable decreased while the amplitude variability of participants who were intrinsically less variable increased as seen in Fig. 5.
Discussion
Experiment 2 investigated the effects of both pitch and tempo on the occurrence of movement entrainment and stability. The specific hypotheses that were examined concerned whether auditory rhythms delivered at an individual’s preferred movement tempo lead to stronger movement entrainment and greater stability, and whether effects induced by rhythms presented slightly slower and faster than an individual’s preferred tempo are strengthened by low and high pitches, respectively.
Corroborating Experiment 1, the results of Experiment 2 showed that participants’ pendulum swinging spontaneously entrained to auditory rhythms and that this entrainment was stronger with low-pitched (i.e., 100 Hz) compared to high-pitched rhythms (i.e., 1600 Hz). It can be noted that the difference between control and high-pitch condition was bordering on statistical significance, suggesting that while entrainment was generally unreliable for the high-pitch metronome in Experiment 2, it may not have been entirely absent (i.e., intermittent entrainment may have occurred for portions of trials). This borderline effect encourages further studies to confirm the difference of entrainment strength between high-pitched and low-pitched rhythms. The results also revealed stronger entrainment when rhythms were presented at the participant’s preferred tempo in line with previous studies that investigated frequency (tempo) detuning effects in visuo-motor entrainment studies (Lopresti-Goodman et al., 2008; Richardson et al., 2007; Schmidt & O’Brien, 1997). However, decreased synchrony compared to the preferred tempo condition was principally observed for conditions with slower rhythms, which can be explained by a tendency for participants’ preferred tempo to increase throughout the experiment. Contrasting with our predictions, the results did not show any interaction between the effects of pitch and tempo on entrainment. Entrainment to slower rhythms was not strengthened by low-pitch and entrainment to faster rhythms was not strengthened by high-pitch.
The results also revealed changes in the mean movement period and amplitude exhibited by participants. Participants’ movement tempo tended to gravitate towards the stimulation tempo, showing the ability of auditory rhythms to drive an individual’s movement towards slower or faster tempi. The results also showed that the mean amplitude of participants increased when auditory rhythms were presented compared to control trials without stimulation, which is in line with increased stride lengths previously reported with auditory metronomes in locomotion studies (Lim et al., 2005; McIntosh et al., 1997; Thaut et al., 1996).
In line with Experiment 1, effects of auditory rhythms on the period and amplitude variability of participants’ movements were observed. The movement period variability of participants who were intrinsically more variable decreased in the presence of auditory rhythms, and (extending the results of Experiment 1) this effect preferentially occurred for auditory rhythms delivered close to the participant’s preferred movement tempo. A decrease of amplitude variability was also found with auditory rhythms, especially with low-pitched rhythms, further demonstrating the benefits of auditory rhythms for the stabilisation of rhythmic movements in time and space. Benefits of low-pitched rhythms over high-pitched rhythms were thus observed across several variables, showing stronger (de)stabilisation effects depending on intrinsic variability and greater stabilisation effects at the group level. However, as these effects were not observed for all the tests conducted, further studies with larger sample sizes are needed to confirm the existence of low-pitch advantages on the stability of self-paced rhythmic movements.
It can also be noted that stabilisation index values were computed using the participant’s intrinsic variability recorded in the Control (Mute) condition and thus there was dependency between the dependent and independent variables in the regression analyses conducted. This issue might be avoided in future studies by assessing participant’s intrinsic variability separately from experimental trials (e.g., before and after, to obtain a baseline). Moreover, assessment of participant’s intrinsic variability before could be used to assign participants with low and high intrinsic variability to different groups for more direct investigation of the effects of intrinsic variability. More direct investigation might also be possible in future studies by comparing healthy participants with patients suffering from motor disorders (e.g., Stroke or Parkinson’s disease) to examine differences in intrinsic movement variability of greater magnitude.
General discussion
The current study investigated two experiments, the effects of pitch and tempo of auditory rhythms on the occurrence and strength of spontaneous auditory–motor entrainment and movement stabilisation. The two experiments showed that low-pitched rhythms have stronger influence on an individual’s rhythmic movement compared to high-pitched rhythms, inducing stronger entrainment as well as tempo and amplitude stabilisation. The results also showed that rhythms presented close to an individual’s preferred movement tempo facilitated movement entrainment and stability, though no interaction between pitch and tempo was revealed across the two experiments—low-pitched rhythms had greater influence on participants’ movement irrespective of the tempo of auditory stimulation.
Stronger effects of low-pitched rhythms on movement
A key finding of the current study is the occurrence of strengthened movement entrainment and greater stability when listening to a low-pitched (100 Hz) compared to a high-pitched (1600 Hz) auditory metronome. The low-pitched metronome strengthened participants’ movement entrainment, in such a way that they moved more consistently in time with the metronome. These results show that low-pitched rhythms have a relatively strong influence on an individual’s timing and motor processes. This is in line with results previously reported by Hove and colleagues at both behavioural and neurophysiological levels (Hove et al., 2007; 2014). It can be noted, however, that our results extend previous studies that revealed low-pitch facilitation with complex and natural polyphonic sounds by showing that this effect also occurs with simple pure tone stimuli. The low-pitch facilitation observed here at the movement level is in line with the results of a recent study that found with Electroencephalography a low-pitch facilitation with simple pure tone stimuli at the neural level for the tracking of auditory rhythms (Lenc, Keller, Varlet, & Nozaradan, 2018). It has been proposed that this effect of low-pitch might be due to the nonlinear dynamics of the human cochlear in the inner ear enhancing time encoding for low-frequency sounds (Hove et al., 2014), although an alternative explanation based on perceptual tolerance for delays in processing low sounds has been recently advanced (Wojtczak et al., 2017). It has also been proposed that the vestibular system plays an important role in this low-pitch facilitation. Auditory rhythms can stimulate the vestibular system, especially when low-pitched, contributing to stronger modulation of an individual’s movement (Todd, et al., 2008, 2009; Todd & Lee, 2015). However, although stimulation of the vestibular system might still have contributed to the effects observed in the current study, it can be noted that the stimulus intensity used here was lower than the threshold for direct stimulation of the vestibular system previously reported (Todd & Cody, 2000). It would, moreover, be interesting to test a wider range of stimulus pitches. 100 Hz was used as the low pitch in the current study but the effects on entrainment and movement stabilisation might differ for other low-pitch values. Hove et al. (2014) used 196 Hz, for instance, and it would be interesting to see whether this pitch might lead to weaker or stronger effects than the ones observed here.
Contrasting with our initial predictions, neither of the two experiments revealed interactions between the pitch and tempo of the auditory stimuli. Entrainment for both slow and fast pendulum swinging was facilitated by low-pitched rhythms (Experiment 1) and stimulus rhythms both slower and faster than an individual’s preferred movement tempo induced stronger entrainment when low-pitched (Experiment 2). Despite the manipulations of the pendulum’s mechanical properties in Experiment 1 and stimulus tempo in Experiment 2, the range of tempi explored in these two experiments remained generally slow (around 1 Hz). This leaves open the possibility that interactions between pitch and tempo, and entrainment strengthened by high-pitched rhythms in particular, might occur with auditory rhythms presented at faster tempi.
It can be noted that Hove and colleagues found that finger-tapping was more strongly attracted to low-pitched sounds when low-pitched and high-pitched sounds were presented together in separate streams at 2 Hz (Hove et al. 2014), suggesting that low-pitch facilitation might occur at least up to this tempo. Furthermore, it has been shown that the involvement of the vestibular system in auditory perception, especially when auditory rhythms are low-pitched, has a strong resonance frequency around 2 Hz, which may have developed through extensive exposure to this tempo during locomotion (Todd & Lee, 2015; MacDougall & Moore, 2005). It has actually been proposed that this link to movement may explain preferences in humans for auditory rhythms around 2 Hz and why tempo in western music, often carried in low-pitched sounds, is usually around 2 Hz (Burger et al., 2014; Fraisse, 1982; Moelants, 2002; van Noorden & Moelants, 1999). The preferred movement tempo of the fingers, which are among the most dexterous human body segments, is also around 2 Hz in general (Fraisse, 1982; Large, 2008), which limits the remaining range of faster tempi where entrainment could be strengthened by high-pitch.
Movement improved in time and space
The results of the two experiments demonstrated that the presentation of auditory rhythmic stimuli, especially low-pitched stimuli, attract an individual’s rhythmic movement to different tempi, increase its amplitude, and stabilise (reduce variability of) its tempo and amplitude. These results corroborate previous studies that showed that auditory rhythmic stimulation can help to improve motor performance (Bood, Nijssen, Van Der Kamp, & Roerdink, 2013; Lim et al., 2005; McIntosh et al., 1997; Roerdink, Bank, Peper, & Beek, 2011; Thaut et al., 1996). Also particularly noteworthy is that the two experiments showed that movement stabilisation in time and space is moderated by individual differences in intrinsic movement variability. Tempo and amplitude stabilisation preferentially occur when an individual’s movement is intrinsically more variable. Tempo and amplitude destabilisation actually tend to occur when an individual’s movement is intrinsically less variable. This finding underscores the importance of taking into account individual differences in intrinsic variability when comparing the effects of different rhythms on movement stability. As revealed by our analyses, failing to take into account intrinsic variability can compromise the validity of conclusions, as analyses at group level might not capture the differences between the two types of rhythms and/or lead to misleading interpretations of their effects or the absence of effects. These results also highlight the particular relevance of auditory rhythmic stimulation for improving motor performance of patients suffering from motor disorders given their higher intrinsic movement variability (Hove & Keller, 2015; Lim et al., 2005; McIntosh et al., 1997). However, it does suggest a more limited relevance of auditory rhythms to stabilise movements of individuals with lower intrinsic variability, such as athletes running or cycling, for instance. The results also demonstrate potential limitations of investigating and optimising auditory rhythmic stimulation for motor rehabilitation based on research conducted with healthy participants who have lower intrinsic movement variability and may be more likely not to be susceptible to benefits of the stimulation.
An important aspect of the present findings is that the movement improvements were observed in a spontaneous synchronisation task. The presentation of an auditory metronome induced spontaneous synchronisation despite participants being only instructed to swing the pendulum at their preferred movement tempo. Although such spontaneous synchronisation is only intermittent and weaker than intentional synchronisation, the stability of participants’ movement was improved in time and space. Entrainment processes underlying spontaneous synchronisation are sufficiently strong to enhance individual motor performance. This finding opens new promising perspectives for using auditory cueing for motor rehabilitation. Indeed, although entrainment processes can be involved in intentional synchronisation tasks as well, intentional synchronisation tasks often require the involvement of error correction and predictive mechanisms that are cognitively, in particular attentionally, more demanding (Pecenka, Engel, & Keller, 2013; Repp & Keller, 2004; Van Der Steen & Keller, 2013). This can be an obstacle in motor rehabilitation, as patients often have attention limitations in addition to motor deficits. The results of the present two experiments suggest that movement improvements are possible in cognitively less demanding rehabilitation protocols via entrainment, which invites further comparisons in the future between spontaneous and intentional synchronisation protocols.
Perspectives for further movement entrainment and improvement
Our results encourage future research to further investigate the effects of pitch of auditory rhythms on the occurrence and strength of auditory–motor entrainment. Greater effects of low-pitched rhythms compared with high-pitched rhythms were found across two experiments and different dependent variables. It will be important to confirm these effects in other studies and determine how robust these effects are within a larger range of low-pitched rhythms. More specifically, it will also be of particular interest to test whether the effects of pitch interact with other properties of auditory rhythms. Other sound properties could be manipulated in future research to examine whether they moderate the low-pitch benefits observed in these two experiments. Manipulating the duration and intensity of the sounds may be particularly relevant given that louder and longer sounds may favour the positive effects of low-pitched rhythms on movement. Todd and Cody (2000) have shown for instance that the involvement of the vestibular system with low-pitched sounds preferentially occurs for loudness above 90 dBA, which is louder than the low-pitch stimuli presented in the current study (i.e., 72 dBA), as mentioned before. Further research is needed, but our results might suggest limited contribution of the vestibular system in the low-pitch enhanced entrainment and stabilisation observed under conditions tested in the present study. The way these low-pitched stimulations are delivered would also be interesting to manipulate in future studies. The use of loudspeakers, especially in enclosed spaces, might favour greater effects of low-pitched rhythms compared to when delivered via headphones, as such free-field, high intensity delivery may result in larger vibrations, and thus, stronger involvement of the vestibular system.
The link between pitch and tempo could be further explored—not in terms of mean tempo (mean inter-onset interval), as examined in Experiment 1 and 2, but in terms of tempo variability (inter-onset interval variability). Indeed, recent research has shown that synchronisation might be facilitated when presenting metronomes with biological variability, characterised by short- and long-term correlations of inter-onset intervals, compared to perfectly regular metronomes (Dotov et al., 2017; Varlet et al., 2016). Tempo is also continuously modulated in music, which can influence the effects induced on listeners’ movements (Leow et al., 2014; Ross et al., 2016; Stupacher et al., 2013). Investigating the role played by the pitch of the sounds in these tempo-changing auditory sequences is also a potentially fruitful avenue for future research, as pitch variation can add higher-order structure and thus stabilise performance (Large, 2008; Large & Palmer, 2002).
Furthermore, although auditory metronomes presented in Experiment 1 and 2 did not provide any spatial information, as they were presented simultaneously and with equal intensity on both sides of the participant’s headphones, the results showed that participants’ movement was improved not only in time but also in space. Specifically, larger movement amplitude and decreased amplitude variability were observed with auditory stimulation. It would be interesting to investigate in future studies whether these spatial effects are strengthened using metronomes containing also spatial information (e.g., left vs. right or distal vs. proximal) and how this enhancement could be moderated by the pitch of the sounds.
To conclude, this study provides new insights into the processes underpinning the occurrence and strength of spontaneous auditory–motor entrainment and movement stabilisation. The findings revealed influences of pitch and tempo of auditory rhythms on movement entrainment and stabilisation, showing that not all auditory rhythms in the environment modulate an individual’s motor system equally. Moreover, the results demonstrate that movement stabilisation in time and space induced by auditory rhythmic stimulation also occurs in spontaneous synchronisation situations, and preferentially for movements that are intrinsically more variable. This highlights the importance of investigating movement and stimulus properties together to better understand auditory–motor entrainment and sensorimotor performance more generally.
References
Bardy, B. G., Hoffmann, C. P., Moens, B., Leman, M., & Dalla Bella, S. (2015). Sound-induced stabilization of breathing and moving. Annals of the New York Academy of Sciences, 1337, 94–100.
Batschelet, E. (1981). Circular statistics in biology (Vol. 371). London: Academic Press.
Boersma, P., & Weenink, D. (2014). Praat: Doing phonetics by computer (Version 5. 3. 79) [Computer program].
Boltz, M. G. (2011). Illusory tempo changes due to musical characteristics. Music Perception, 28, 367–386.
Bood, R. J., Nijssen, M., Van Der Kamp, J., & Roerdink, M. (2013). The power of auditory-motor synchronization in sports: enhancing running performance by coupling cadence with the right beats. PloS One, 8, e70758.
Broze, Y., & Huron, D. (2013). Is higher music faster? Pitch–speed relationships in Western compositions. Music Perception, 31, 19–31.
Burger, B., Thompson, M. R., Luck, G., Saarikallio, S., & Toiviainen, P. (2013). Influences of rhythm-and timbre-related musical features on characteristics of music-induced movement. Frontiers in Psychology, 4, 183.
Burger, B., Thompson, M. R., Luck, G., Saarikallio, S. H., & Toiviainen, P. (2014). Hunting for the beat in the body: on period and phase locking in music-induced movement. Frontiers in Human Neuroscience, 8, 903.
Coey, C. A., Varlet, M., & Richardson, M. J. (2012). Coordination dynamics in a socially situated nervous system. Frontiers in Human Neuroscience, 6, 164.
Coey, C. A., Varlet, M., Schmidt, R. C., & Richardson, M. J. (2011). Effects of movement stability and congruency on the emergence of spontaneous interpersonal coordination. Experimental Brain Research, 211, 483–493.
Collier, W. G., & Hubbard, T. L. (2004). Musical scales and brightness evaluations: effects of pitch, direction, and scale mode. Musicae Scientiae, 8, 151–173.
Demos, A. P., Chaffin, R., Begosh, K. T., Daniels, J. R., & Marsh, K. L. (2012). Rocking to the beat: Effects of music and partner’s movements on spontaneous interpersonal coordination. Journal of Experimental Psychology: General, 141, 49–53.
Dotov, D. G., Bayard, S., de Cock, V. C., Geny, C., Driss, V., Garrigue, G., Bardy, B., & Bella, D., S (2017). Biologically-variable rhythmic auditory cues are superior to isochronous cues in fostering natural gait variability in Parkinson’s disease. Gait and Posture, 51, 64–69.
Eitan, Z., & Granot, R. Y. (2006). How music moves: Musical parameters and images of motion. Music Perception, 23, 221–247.
Fraisse, P. (1982). Rhythm and tempo. In D. Deutsch (Ed.), The psychology of music (pp. 149–180). Orlando: Academic Press.
Fuchs, A., Jirsa, V. K., Haken, H., & Kelso, J. S. (1996). Extending the HKB model of coordinated movement to oscillators with different eigenfrequencies. Biological Cybernetics, 74, 21–30.
Fujioka, T., Trainor, L. J., Large, E. W., & Ross, B. (2012). Internalized timing of isochronous sounds is represented in neuromagnetic beta oscillations. Journal of Neuroscience, 32, 1791–1802.
Fujioka, T., Trainor, L. J., Ross, B., Kakigi, R., & Pantev, C. (2005). Automatic encoding of polyphonic melodies in musicians and nonmusicians. Journal of Cognitive Neuroscience, 17, 1578–1592.
Grahn, J. A., & Brett, M. (2007). Rhythm and beat perception in motor areas of the brain. Journal of Cognitive Neuroscience, 19, 893–906.
Haken, H., Kelso, J. S., & Bunz, H. (1985). A theoretical model of phase transitions in human hand movements. Biological Cybernetics, 51, 347–356.
Hove, M. J., & Keller, P. E. (2015). Impaired movement timing in neurological disorders: rehabilitation and treatment strategies. Annals of the New York Academy of Sciences, 1337, 111–117.
Hove, M. J., Keller, P. E., & Krumhansl, C. L. (2007). Sensorimotor synchronization with chords containing tone-onset asynchronies. Attention, Perception, and Psychophysics, 69, 699–708.
Hove, M. J., Marie, C., Bruce, I. C., & Trainor, L. J. (2014). Superior time perception for lower musical pitch explains why bass-ranged instruments lay down musical rhythms. Proceedings of the National Academy of Sciences of the United States of America, 111, 10383–10388.
Keller, P. E., & Repp, B. H. (2005). Staying offbeat: Sensorimotor syncopation with structured and unstructured auditory sequences. Psychological Research Psychologische Forschung, 69, 292–309.
Keller, P. E., & Rieger, M. (2009). Special issue - Musical movement and synchronization. Music Perception: An Interdisciplinary Journal, 26, 397–400.
Kelso, J. S. (1995). Dynamic patterns: The self-organization of brain and behavior. MIT press.
Kugler, P. N., & Turvey, M. T. (1987). Information, natural law, and the self-assembly of rhythmic movement. Routledge.
Large, E. W. (2000). On synchronizing movements to music. Human Movement Science, 19, 527–566.
Large, E. W. (2008). Resonating to musical rhythm: Theory and experiment. In S. Grondin (Ed.), The psychology of time (pp. 189–231). Cambridge: Emerald.
Large, E. W., & Palmer, C. (2002). Perceiving temporal regularity in music. Cognitive Science, 26, 1–37.
Leman, M., Moelants, D., Varewyck, M., Styns, F., van Noorden, L., & Martens, J. P. (2013). Activating and relaxing music entrains the speed of beat synchronized walking. PloS One, 8, e67932.
Lenc, T., Keller, P. E., Varlet, M., & Nozaradan, S. (2018). Neural tracking of the musical beat is enhanced by low-frequency sounds. Proceedings of the National Academy of Sciences of the United States of America, 115, 8221–8226.
Leow, L. A., Parrott, T., & Grahn, J. A. (2014). Individual differences in beat perception affect gait responses to low-and high-groove music. Frontiers in Human Neuroscience, 8, 811.
Lim, I., van Wegen, E., de Goede, C., Deutekom, M., Nieuwboer, A., Willems, A., Jones, D., Rochester, L., & Kwakkel, G. (2005). Effects of external rhythmical cueing on gait in patients with Parkinson’s disease: a systematic review. Clinical Rehabilitation, 19, 695–713.
Lopresti-Goodman, S. M., Richardson, M. J., Silva, P. L., & Schmidt, R. C. (2008). Period basin of entrainment for unintentional visual coordination. Journal of Motor Behavior, 40, 3–10.
MacDougall, H. G., & Moore, S. T. (2005). Marching to the beat of the same drummer: the spontaneous tempo of human locomotion. Journal of Applied Physiology, 99, 1164–1173.
Malcolm, M. P., Massie, C., & Thaut, M. (2009). Rhythmic auditory-motor entrainment improves hemiparetic arm kinematics during reaching movements: a pilot study. Topics in Stroke Rehabilitation, 16, 69–79.
Marie, C., & Trainor, L. J. (2013). Development of simultaneous pitch encoding: infants show a high voice superiority effect. Cerebral Cortex, 23, 690–669.
McIntosh, G. C., Brown, S. H., Rice, R. R., & Thaut, M. H. (1997). Rhythmic auditory-motor facilitation of gait patterns in patients with Parkinson’s disease. Journal of Neurology, Neurosurgery and Psychiatry, 62, 22–26.
Moelants, D. (2002). Preferred tempo reconsidered. In C. Stevens, D. Burnham, G. McPherson, E. Schubert, and J. Renwick (Ed.), Proceedings of the 7th international conference on music perception and cognition (pp. 580–583). Adelaide: Causal Production.
Moore, B. C., Glasberg, B. R., & Baer, T. (1997). A model for the prediction of thresholds, loudness, and partial loudness. Journal of the Audio Engineering Society, 45, 224–240.
Néda, Z., Ravasz, E., Brechet, Y., Vicsek, T., & Barabási, A. L. (2000). Self-organizing processes: The sound of many hands clapping. Nature, 403, 849–850.
Novembre, G., Varlet, M., Muawiyath, S., Stevens, C. J., & Keller, P. E. (2015). The E-music box: an empirical method for exploring the universal capacity for musical production and for social interaction through music. Royal Society Open Science, 2, 150286.
Nozaradan, S., Peretz, I., & Keller, P. E. (2016). Individual differences in rhythmic cortical entrainment correlate with predictive behavior in sensorimotor synchronization. Scientific Reports, 6, 20612.
O’Brien, F., & Cousineau, D. (2014). Representing error bars in within-subject designs in typical software packages. Quantitative Methods for Psychology, 10, 56–67.
Pecenka, N., Engel, A., & Keller, P. E. (2013). Neural correlates of auditory temporal predictions during sensorimotor synchronization. Frontiers in Human Neuroscience, 7, 380.
Peckel, M., Pozzo, T., & Bigand, E. (2014). The impact of the perception of rhythmic music on self-paced oscillatory movements. Frontiers in Psychology, 5, 1037.
Phillips-Silver, J., Aktipis, C. A., & Bryant, G. A. (2010). The ecology of entrainment: Foundations of coordinated rhythmic movement. Music Perception: An Interdisciplinary Journal, 28, 3–14.
Phillips-Silver, J., & Trainor, L. J. (2005). Feeling the beat: movement influences infant rhythm perception. Science, 308, 1430–1430.
Phillips-Silver, J., & Trainor, L. J. (2008). Vestibular influence on auditory metrical interpretation. Brain and Cognition, 67, 94–102.
Pikovsky, A., Rosenblum, M., & Kurths, J. (2003). Synchronization: a universal concept in nonlinear sciences (Vol. 12). Cambridge: Cambridge university press.
Repp, B. H. (2005). Sensorimotor synchronization: a review of the tapping literature. Psychonomic Bulletin and Review, 12, 969–992.
Repp, B. H. (2006). Does an auditory distractor sequence affect self-paced tapping? Acta Psychologica, 121, 81–107.
Repp, B. H., & Keller, P. E. (2004). Adaptation to tempo changes in sensorimotor synchronization: Effects of intention, attention, and awareness. Quarterly Journal of Experimental Psychology Section A, 57, 499–521.
Repp, B. H., & Knoblich, G. (2007). Action can affect auditory perception. Psychological Science, 18, 6–7.
Repp, B. H., & Su, Y. H. (2013). Sensorimotor synchronization: a review of recent research (2006–2012). Psychonomic Bulletin and Review, 20, 403–452.
Richardson, M. J., Marsh, K. L., Isenhower, R. W., Goodman, J. R., & Schmidt, R. C. (2007). Rocking together: Dynamics of intentional and unintentional interpersonal coordination. Human Movement Science, 26, 867–891.
Roerdink, M., Bank, P. J., Peper, C. L. E., & Beek, P. J. (2011). Walking to the beat of different drums: Practical implications for the use of acoustic rhythms in gait rehabilitation. Gait and Posture, 33, 690–694.
Romero, V., Coey, C., Schmidt, R. C., & Richardson, M. J. (2012). Movement coordination or movement interference: Visual tracking and spontaneous coordination modulate rhythmic movement interference. PLoS One, 7, e44761.
Ross, J. M., Warlaumont, A. S., Abney, D. H., Rigoli, L. M., & Balasubramaniam, R. (2016). Influence of musical groove on postural sway. Journal of Experimental Psychology: Human Perception and Performance, 42, 308–319.
Scherer, K. R., & Oshinsky, J. S. (1977). Cue utilization in emotion attribution from auditory stimuli. Motivation and Emotion, 1, 331–346.
Schmidt, R. C., & O’Brien, B. (1997). Evaluating the dynamics of unintended interpersonal coordination. Ecological Psychology, 9(3), 189–206.
Schmidt, R. C., & Richardson, M. J. (2008). Dynamics of interpersonal coordination. In A. Fuchs & V. K. Jirsa (Eds.), Coordination: Neural, behavioral and social dynamics (pp. 281–308). Heidelberg: Springer.
Schmidt, R. C., Richardson, M. J., Arsenault, C., & Galantucci, B. (2007). Visual tracking and entrainment to an environmental rhythm. Journal of Experimental Psychology: Human Perception and Performance, 33(4), 860–870.
Schmidt, R. C., & Turvey, M. T. (1992). Long-term consistencies in assembling coordinated rhythmic movements. Human Movement Science, 11(3), 349–376.
Snyder, J., & Krumhansl, C. L. (2001). Tapping to ragtime: Cues to pulse finding. Music Perception: An Interdisciplinary Journal, 18, 455–489.
Stupacher, J., Hove, M. J., & Janata, P. (2016). Audio features underlying perceived groove and sensorimotor synchronization in music. Music Perception: An Interdisciplinary Journal, 33, 571–589.
Stupacher, J., Hove, M. J., Novembre, G., Schütz-Bosbach, S., & Keller, P. E. (2013). Musical groove modulates motor cortex excitability: a TMS investigation. Brain and Cognition, 82, 127–136.
Styns, F., van Noorden, L., Moelants, D., & Leman, M. (2007). Walking on music. Human Movement Science, 26, 769–785.
Tamir-Ostrover, H., & Eitan, Z. (2015). Higher is faster: Pitch register and tempo preferences. Music Perception, 33, 179–198.
Thaut, M. H., McIntosh, G. C., & Rice, R. R. (1997). Rhythmic facilitation of gait training in hemiparetic stroke rehabilitation. Journal of the Neurological Sciences, 151, 207–212.
Thaut, M. H., McIntosh, G. C., Rice, R. R., Miller, R. A., Rathbun, J., & Brault, J. M. (1996). Rhythmic auditory stimulation in gait training for Parkinson’s disease patients. Movement Disorders, 11, 193–200.
Todd, N. P., & Cody, F. W. (2000). Vestibular responses to loud dance music: A physiological basis of the “rock and roll threshold”? The Journal of the Acoustical Society of America, 107, 496–500.
Todd, N. P., & Lee, C. S. (2015). The sensory-motor theory of rhythm and beat induction 20 years on: a new synthesis and future perspectives. Frontiers in Human Neuroscience, 9, 444.
Todd, N. P., Rosengren, S. M., & Colebatch, J. G. (2008). Tuning and sensitivity of the human vestibular system to low-frequency vibration. Neuroscience Letters, 444, 36–41.
Todd, N. P., Rosengren, S. M., & Colebatch, J. G. (2009). A utricular origin of frequency tuning to low-frequency vibration in the human vestibular system? Neuroscience Letters, 451, 175–180.
Torre, K., Varlet, M., & Marmelat, V. (2013). Predicting the biological variability of environmental rhythms: Weak or strong anticipation for sensorimotor synchronization? Brain and Cognition, 83, 342–350.
Trainor, L. J., Gao, X., Lei, J. J., Lehtovaara, K., & Harris, L. R. (2009). The primal role of the vestibular system in determining musical rhythm. Cortex, 45, 35–43.
Van Der Steen, M. C., & Keller, P. E. (2013). The ADaptation and Anticipation Model (ADAM) of sensorimotor synchronization. Frontiers in Human Neuroscience, 7, 253.
Van Dyck, E., Moelants, D., Demey, M., Deweppe, A., Coussement, P., & Leman, M. (2013). The impact of the bass drum on human dance movement. Music Perception: An Interdisciplinary Journal, 30, 349–359.
Van Dyck, E., Moens, B., Buhmann, J., Demey, M., Coorevits, E., Bella, D., S., & Leman, M. (2015). Spontaneous entrainment of running cadence to music tempo. Sports Medicine-open, 1, 15.
van Noorden, L., & Moelants, D. (1999). Resonance in the perception of musical pulse. Journal of New Music Research, 28, 43–66.
Varlet, M., Bucci, C., Richardson, M. J., & Schmidt, R. C. (2015). Informational constraints on spontaneous visuomotor entrainment. Human Movement Science, 41, 265–281.
Varlet, M., Coey, C. A., Schmidt, R. C., Marin, L., Bardy, B. G., & Richardson, M. J. (2014). Influence of stimulus velocity profile on rhythmic visuomotor coordination. Journal of Experimental Psychology: Human Perception and Performance, 40, 1849–1860.
Varlet, M., Coey, C. A., Schmidt, R. C., & Richardson, M. J. (2012). Influence of stimulus amplitude on unintended visuomotor entrainment. Human Movement Science, 31, 541–552.
Varlet, M., Novembre, G., & Keller, P. E. (2017). Dynamical entrainment of corticospinal excitability during rhythmic movement observation: A Transcranial Magnetic Stimulation study. European Journal of Neuroscience, 45, 1465–1472.
Varlet, M., Schmidt, R. C., & Richardson, M. J. (2016). Influence of internal and external noise on spontaneous visuomotor synchronization. Journal of Motor Behavior, 48, 122–131.
Wojtczak, M., Mehta, A. H., & Oxenham, A. J. (2017). Rhythm judgments reveal a frequency asymmetry in the perception and neural coding of sound synchrony. Proceedings of the National Academy of Sciences of the United States of America, 114, 1201–1206.
Zamm, A., Pfordresher, P. Q., & Palmer, C. (2015). Temporal coordination in joint music performance: Effects of endogenous rhythms and auditory feedback. Experimental Brain Research, 233, 607–615.
Zamm, A., Wellman, C., & Palmer, C. (2016). Endogenous rhythms influence interpersonal synchrony. Journal of Experimental Psychology: Human Perception and Performance, 42, 611–616.
Zatorre, R. J., Chen, J. L., & Penhune, V. B. (2007). When the brain plays music: auditory–motor interactions in music perception and production. Nature Reviews Neuroscience, 8, 547–558.
Zbikowsky, L. (2002). Conceptualizing music: Cognitive structure, theory, and analysis. New York: Oxford University Press.
Acknowledgements
This work was supported by an Australian Research Council Discovery project (DP170104322) and an Australian Research Council Future Fellowship grant awarded to P.K. (FT140101162).
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interest
Manuel Varlet declares that he has no conflict of interest. Rohan Williams declares that he has no conflict of interest. Peter Keller declares that he has no conflict of interest.
Ethical approval
All procedures performed in studies involving human participants were in accordance with the ethical standards of the institutional and/or national research committee and with the 1964 Helsinki declaration and its later amendments or comparable ethical standards.
Rights and permissions
About this article

Cite this article
Varlet, M., Williams, R. & Keller, P.E. Effects of pitch and tempo of auditory rhythms on spontaneous movement entrainment and stabilisation. Psychological Research 84, 568–584 (2020). https://doi.org/10.1007/s00426-018-1074-8
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00426-018-1074-8