
Abstract
Theories assuming an effect-based coding of action predict that motor responses become activated by the perception of the responses’ sensory effects. In accordance with this prediction it was found that responding to a visual target is faster and more accurate when the target is briefly preceded by the visual effect of the required response. Most importantly, this effect-induced response priming was independent of prime perceptibility and it occurred even when the prime was not consciously discriminable. Beyond ruling out alternative interpretations of earlier induction studies in terms of deliberate response biases, this suggests that effect codes evoke their associated motor patterns in a highly automatic manner not affording conscious mediation. The results accord with a functional dissociation between the consciousness-mediated implementation and the consciousness-independent realization of action goals.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Response priming by subliminal action effects
How do humans mentally control their body movements? This question continues to be a central and challenging one in movement sciences. A rather radical suggestion on this issue has been prompted by introspective researchers of the 19th century (Harleß, 1861; James, 1890; Lotze, 1852). They proposed that movements are exhaustively coded in terms of their sensory effects (rather than by abstract motor commands or motor programs, Keele, 1968; Schmidt, 1975). From this perspective even an action as simple as pressing a response button is represented by its various sensory consequences, like the tactile sensation from hitting the response button, or the auditory ‘click’ from pressing it down. Selecting a certain action is mediated by recollecting these perceptual effects, which then activate the particular motor pattern that normally brings these anticipated re-afferences about.
This “ideo-motor” (IM) view of action control allows for some interesting experimental predictions. Most notably it implies that motor patterns should become activated by externally stimulating the effect codes that represent them. In other words, actions should be induced by perceiving their effects. Several demonstrations of such effect-based action induction, both anecdotal and experimental ones, have been reported. Many of them relied on perceived body motion (as for example in imitation studies), which can be construed as the most proximal and natural effect of acting (e.g., Iacoboni et al., 1999; Meltzoff & Moore, 1977; Stürmer, Aschersleben, & Prinz, 2000).
Action induction by effect perception is, however, not confined to proximal and natural action effects. To illustrate this, consider a recent series of studies by Hommel and coworkers (1996; Elsner & Hommel, 2001). Initially, participants freely chose to carry out left or right key presses. Each response produced one of two artificial effects (e.g., left key press→low tone, right key press→high tone). In a subsequent choice reaction task, the responses no longer produced any tone effects. However, the tones now served as stimuli, which required either a left or a right key press. Responses were faster (and more accurate) when the stimulus tone matched the former effect of the required response, and was thus IM compatible (e.g., low tone→left key press, high tone→right key press) than when it did not match, and was thus IM incompatible (e.g., low tone→right key press, high tone→left key press). This suggests that even distal and artificial sensory effects become associated with the motor patterns that effectuate them, whereby these effects acquire the power to automatically induce these motor patterns when perceived later on (cf. also Beckers, De Houwer, & Eelen, 2002; Zießler & Nattkemper, 2002).
The present study explored one particular aspect of effect-based action induction: Can motor responses become activated by subliminal effect perception? Answering this question would help to clarify the functional nature of effect-based response activation. It is often implied that this activation is highly automatic, more or less independent of deliberate control. Empirical evidence on this issue, however, is inconclusive. For example, some studies found that effect-based action induction is independent of concurrent dual task requirements (Greenwald, 1972; Greenwald & Shulman, 1973; Greenwald, 2003), whereas others found that it is not (Klapp, Port-Graham, & Hoijfield, 1991; Lien, Proctor, & Allen, 2002). Demonstrating that effect codes can bias behavior without conscious mediation would strongly support the automatic nature of effect-based response activation.
At first glance, influences of subliminal effects on behavior might seem counterintuitive. Normally we are aware of our goals, so why should action effects affect the motor system unconsciously? Indeed James (1890) spoke of “images” that guide our actions (i.e., conscious, percept-like goal representations). Yet by relying on introspection James and his contemporaries could barely derive any other conclusion. Experimental brands of IM theory are far more careful with regard to the role of consciousness in action control. For example, Hommel, Müsseler, Aschersleben, and Prinz (2001) note that “awareness can sometimes emerge in perception (as it can in any other cognitive function) without, however, playing a functional role itself” (p. 915). And indeed ample evidence suggests that the perceptual processing mediating action control remains unconscious and neurally distinct from the perceptual processing that mediates phenomenal experience. This is evidenced, for example, by the resistance of action against visual illusions, and by dissociated consequences of brain injury for object recognition and visually guided action (e.g., Goodale & Humphrey, 1998). In view of such evidence it is certainly tenable that sensory effects might induce motor responses unconsciously too.
To explore this issue the participants in the present study went through three different experimental phases. In a first acquisition phase, they freely chose between left-hand or right-hand key presses. Each action produced one of two visual effects, either an up-pointing or a down-pointing arrow on a presentation display. Every effort was made to promote the acquisition of stable associations between these manual responses and their visual effects. Firstly, during acquisition the effects were clearly perceptible. Secondly, participants were explicitly informed about the response-effect (R-E) mapping. Thirdly, to direct attention to the specific R-E mapping, and to promote its encoding, each action occasionally produced an irregular effect (i.e., the normal effect of the alternative response), and participants were urged to indicate these deviations as quickly as possible.
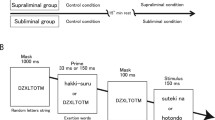
Following this R-E acquisition phase, participants entered a test phase, where they performed choice reactions to color stimuli, e.g., they pressed a left response button to a green target and a right button to a blue target or vice versa (cf. Fig. 1). The target stimuli were preceded by either an up-pointing or a down-pointing prime arrow that had served as a response effect in the acquisition phase. The outline of the prime arrows fitted exactly into a cutting in the center of the imperative color targets, which produces very efficient, “meta-contrast” masking when the prime is presented only briefly (Neumann & Klotz, 1994; Vorberg, Mattler, Heinecke, Schmidt, & Schwarzbach, 2003). All combinations of effect arrows and color targets were equally frequent. Thus, in half of the cases the arrow was the (former) effect of the afforded response (i.e., IM compatible), whereas it was the effect of the alternative (i.e., not afforded) response in the other half of the cases (i.e., IM incompatible). Response activation by the masked arrows would be indicated by faster and more accurate responses in IM-compatible trials than in IM-incompatible ones. After finishing these response priming blocks, a third task affording an unspeeded identification of primes had to performed to assess prime perceptibility.

Basic procedure of the study. In an initial response effect (R-E) acquisition phase participants carried out left/right key presses which contingently produced up-pointing/down-pointing arrows as visual effects. Afterwards participants performed a choice reaction task to the color of a target. The target was preceded by the arrows that had served as response effects in the former acquisition phase
To explore the role of effect awareness for response priming, two groups of participants were tested. One group received primes presented very briefly (14 ms) rendering the primes essentially invisible (Kunde, 2003a). The other group received primes presented slightly longer (56 ms), rendering these primes clearly discriminable. The crucial question was whether we would find compatibility influences and whether these would emerge even when primes were indiscriminable.
Method
Participants
Thirty-two undergraduates from the University of Würzburg participated in fulfillment of a course requirement or for a fee of 8 Euro. The entire experiment took about 45 min.
Apparatus and stimuli
An IBM-compatible computer with a 17-inch VGA display was used for stimulus presentation and response sampling. Stimulus presentation was synchronized with the vertical retraces of a 70-Hz monitor, resulting in a vertical refresh duration of approximately 14.5 ms. The stimuli were presented on a white background. The viewing distance was approximately 60 cm. Responses were executed with the index fingers of both hands on external response buttons (1.7 × 1.7 cm). A third button was used for the ‘veto’ responses in the acquisition phase and depressed with the right foot (see below).
Procedure
Acquisition phase
Each acquisition trial started with a gray central fixation cross measuring .5 × .5 cm. Participants were free to press the left or right response key upon fixation cross presentation, with the constraints that both keys should be pressed approximately equally frequently and in random order. After every 32 trials a brief visual message informed participants how often each key had been pressed. When no response was detected after 1,000 ms following cross onset, the message ‘please react faster’ was displayed for 500 ms. Pressing a key replaced the fixation cross by an up-pointing or down-pointing gray effect arrow of 200 ms duration and measuring .5 × .5 cm. For half the participants a left response (regularly) produced an up-pointing arrow, and a right response (regularly) produced a down-pointing arrow, whereas this R-E mapping was reversed for the other half of the participants.Footnote 1 This ‘regular’ R-E mapping was announced to the participants in advance. However, in 12.5% of the trials the responses produced an ‘irregular’ effect, which was the ‘regular’ effect of the alternative response. In order to promote the encoding and retention of the regular R-E mapping, participants were to indicate the detection of such an event within a time window of 1,000 ms following arrow onset by depressing of a response button with the right foot (a so called ‘veto response’). A correct (and timely) veto response produced the message ‘good job!’ on the screen for 500 ms. If no response was detected within the prescribed time window the message ‘attention, irregular effect!’ was displayed. A false veto response produced the message ‘careful, effect was correct!’ All feedback messages were displayed in German. Five hundred milliseconds after the offset of the effect arrow (or any of the aforementioned feedback messages) the fixation cross for the next trial was presented. Participants performed four blocks of 32 acquisition trials.
Test phase
Immediately following the last acquisition trial, a written instruction on the screen announced a new task. Each test trial started with a 20-ms auditory warning click (100 Hz). Two hundred milliseconds later one of the two arrows that served as response effects in the preceding acquisition phase was presented. For half of the participants the arrow was presented for one refresh cycle of the display (approximately 14 ms) whereas for the other half it was presented for four cycles (approximately 56 ms). After a 28-ms blank interval a green or blue target, which at the same time served as a mask, was presented for 200 ms. The outline of the primes fitted exactly in a cutting in the center of the target (cf. Fig. 1). The form of the target resulted from superimposing (and size-scaling by the factor 2.4) the up-pointing and down-pointing arrows that had served as response effects during acquisition. This rendered the target perceptually similar to both primes, which generally increases subliminal priming effects (e.g., Kunde, Kiesel, & Hoffmann, 2003). Half of the participants responded to a blue target with a left response and to a green target with a right response, whereas this mapping was reversed for the other participants. All combinations of target color (and thus response) and effect-prime (up or down) were equally frequent. Thus, in one half of the trials the effect-prime was IM compatible and in the other half it was IM incompatible. The responses no longer produced effects in the test phase to avoid the effects becoming associated with the response-affording targets. Doing so ensured that priming effects could be attributed to response activation rather than to enhanced target processing (cf. Elsner & Hommel, 2001, p. 232). Responses later than 1,500 ms after target onset were counted as errors. An error produced the message ‘Fehler’ (error) on the screen for 500 ms. One-thousand five-hundred milliseconds after response (or the error message) the warning click for the next trial started. The test phase consisted of six miniblocks of 16 trials that resulted from the combination of 2 primes (up or down) × 2 targets (blue or green) × 4 replications.
Prime discrimination phase
The same sequence of events as in the test trials was used to assess prime discrimination. Participants were informed about the nature and frequency of the primes. They were to decide at leisure which prime was presented in each individual trial by typing in a ‘1’ for an up-pointing arrow, or a ‘2’ for a down-pointing arrow on a standard PC keyboard. No feedback regarding accuracy was given. Participants performed 64 prime discrimination trials (2 primes × 2 masks × 16 replications).
Results
Acquisition phase
The participants followed the equiprobability instruction of the acquisition phase virtually perfectly. They performed 48.7% left responses and 47.9% right responses. In 3.4% no response was carried out in the prescribed time window. The occurrence of an irregular response effect was missed, or indicated too late, in 25.5% of the cases. False alarms were virtually absent (0.8%). The correct veto response time was 590 ms.
Prime discrimination performance
With a prime duration of 14 ms, the mean percentage of correct prime discriminations was 54.0%, and not significantly different from chance level, F < 1. Mean d’ was 0.22.Footnote 2 In contrast to a prime duration of 56 ms the mean percentage of correct prime discriminations was 69.0%,Footnote 3 which is well above the chance rate, p < .01. Mean d’ was 1.27. Thus, as intended, primes were indiscriminable with a duration of 14 ms whereas they were clearly discriminable with a prime duration of 56 ms.
Response priming
Mean response times (RTs) from correct choice reactions were submitted to an ANOVA with the repeated measure of IM prime compatibility (whether the prime was the effect of the requested response or not) and prime discriminability (discriminable [56 ms] or indiscriminable [14 ms]) as between-subjects factor. The mean RTs and error rates from the factorial combination of these factors are listed in Table 1. Responses were slower with discriminable primes than with indiscriminable ones, F(1,30) = 5.89, p < .05. The reasons for this are not entirely clear. It seems possible, however, that supraliminal primes promoted the use of a more conservative response criterion to avoid premature responses to the sudden prime onset. More importantly, responses were faster with compatible primes than with incompatible ones, F(1,30) = 5.69, p < .05. The interaction of prime discriminability and prime compatibility was far from significance (F < 1), and the compatibility effect was also significant when exclusively tested for the group with invisible primes, t(15) = 2.53, p < .05. To explore the location of the compatibility effect within the RT distribution, proportional bins (quintiles) of the rank-ordered RTs were computed for compatible and incompatible primes (cf. Ratcliff, 1979). An ANOVA including quintile (1–5) as an additional factor revealed no difference in the compatibility effect between quintiles (F < 1). From the first to fifth quintile the compatibility effect amounted to 7, 6, 8, 9, and 19 ms, and was significant in all but the fourth bin (ps < .05, one-tailed). Thus, the compatibility effect was more or less present in all parts of the RT distribution. The correlation between the RT congruency effect (i.e., RTincongruent−RTcongruent) and prime discrimination performance, as measured by d’ and computed across all participants, was r = −.11 and far from significance (p = .55).
The same analyses for error rates replicated the compatibility effect in RTs, i.e., more accurate responses with compatible primes than with incompatible ones, F(1,30) = 4.65, p < .05. This effect was more pronounced with discriminable primes, producing a nonsignificant interaction of the two factors, F(1, 30) = 2.95, p = .096.
Discussion
The present study was prompted by the idea that motor actions are cognitively coded and thus accessed by their sensory effects. This view predicts that perceptual stimulation of effect codes should induce the motor pattern that, according to experience, normally brings these effects about. In accordance with this prediction responses were faster (and more accurate) when a response-affording target was preceded by the effect of the requested response than when preceded by the effect of an alternative response. To this end, the present study replicated earlier work and extended it to a new paradigm. More importantly, however, visual effects primed their associated responses even when presented subliminally, rendering prime identity inaccessible.
On the methodological side, this finding rules out some objections against earlier induction studies. Effect-based response priming effects are normally quite small (typically ranging between 9 and 15 ms, e.g., Hommel, 1996). It may thus be argued that they originate from only a small portion of particularly delayed reactions. For example, participants may sometimes deliberately postpone incompatible responses in order to meet the (normally quite obvious) expected outcome of the experiment. Or they may (at least initially) simply be surprised about the co-occurrence of responses and effects that had never been experienced together before. Finding effect-based response priming without knowing which effect was presented clearly rules out such consciousness-mediated biases.
On the theoretical side, the present study, in accordance with existing action-induction studies, portrays a surprisingly automatic machinery of effect-based response generation. Firstly, response priming occurs even when effects are to be ignored during acquisition (Hommel, 1996). This suggests that movements and effects become automatically associated by virtue of repeated experience (but see Zießler & Nattkemper, 2002). Secondly, selecting a response seems to inevitably comprise the recollection of its contingent effects. For example, responses in choice reaction tasks are slower and less accurate when these responses produce incompatible but irrelevant effects (Kunde, 2001, 2003b; Koch & Kunde, 2002). This suggests that sensory effects serve as mental addresses for response selection, and thus become anticipated prior to overt response. Thirdly, as the present study demonstrated, effect codes induce their associated motor patterns without conscious mediation, which points to a highly automatic spreading of activation from effect codes to motor codes.
As already noted, the latter finding might seem counterintuitive when viewed from the perspective of goal-oriented action. Normally we know what we are aiming at. So why should effects evoke actions subliminally? Yet this apparent contradiction is resolved by setting apart two aspects of goal-oriented action, goal implementation and goal realization. Whereas it is taken for granted that implementing a goal is mediated by, or at least associated with, awareness, the realization of an implemented goal need not necessarily be. Indeed Lotze (1852) was already speculating about this distinction when he stated “We can only generate those psychic states [i.e., goal images, W.K.] that serve as a starting point for these psychic processes, which then unroll in accordance with our means, following laws and mediations that elude our awareness” (Lotze, 1852, p.288, translation by the author). The present study represents a nice empirical verification of this conjecture.
To avoid any misunderstanding a final note is warranted. Although we found subliminal priming after responses had become associated with consciously experienced effects, we did not explore the question whether responses can become associated with effects that participants were already unaware of during acquisition—interesting from the perspective of implicit learning. Existing evidence suggests, however, that associations with unconscious stimuli are generally very hard to acquire and confined to particular stimulus and response categories (e.g., fear-inducing stimuli and autonomous responses, Öhmann & Soares, 1998). Indeed, several informal attempts in our lab to establish associations between actions and subliminal effects of the type used here failed to yield any reliable results. Thus, awareness is apparently irrelevant for the functioning of action-effect associations that are already acquired. It is crucial, however, for the acquisition of such associations itself. This issue certainly warrants future research.
Notes
Counterbalancing the R-E mapping eliminates potential influences of coding preferences in orthogonal S-R ensembles (e.g., “right” being more closely related to “up” than to “left”, Weeks & Proctor, 1991). Still, a left response that consistently produces, say, an up-pointing arrow might be recoded as being “up” (or vice versa, the up-pointing arrow might be recoded as being “left”). This, however, is not only true for arrows but for any contingent R-E relationship (e.g., a high tone might be recoded as “left” when produced by a left key press or vice versa, Elsner & Hommel, 2001). Note that this is not an alternative account of the present priming effects as much as a rephrasing of the proposed ideo-motor account: Responses and effects become linked with each other bi-directionally, whereby they acquire each others’ properties.
The procedure by Hautus (1995) for the correction of extreme values in calculating d’ was applied.
The prime discrimination data of one participant were lost, so this mean is based on 15 participants. The response priming data of this participant were well within the range of the other subjects, and excluding this participant from analysis did not alter the overall data pattern in any significant respect.
References
Beckers, T., De Houwer, J., & Eelen, P. (2002). Automatic integration of non-perceptual action effect features: The case of associative affective Simon effect. Psychological Research, 66, 166–173.
Elsner, B., & Hommel, B. (2001). Effect anticipation and action control. Journal of Experimental Psychology: Human Perception and Performance, 27, 229–240.
Goodale, M. A., & Humphrey, G. K. (1998). The objects of action and perception. Cognition, 67, 181–207.
Greenwald, A. G. (1972). On doing two things at once: Timesharing as a function of ideomotor compatibility. Journal of Experimental Psychology, 94, 52–57.
Greenwald, A. G. (2003). On doing two things at once. III. Confirmation of perfect timesharing when simultaneous tasks are ideomotor compatible. Journal of Experimental Psychology: Human Perception and Performance, 29, 859–868.
Greenwald, A. G., & Shulman, H. G. (1973). On doing two things at once. II. Elimination of the psychological refractory period effect. Journal of Experimental Psychology, 101, 70–76.
Harleß, E. (1861). Der Apparat des Willens. Zeitschrift für Philosophie und philosophische Kritik, 38, 50–73.
Hautus, M. J. (1995). Corrections for extreme proportions and their biasing effects on estimated values of d’. Behavior Research Methods, Instruments & Computers, 27, 46–51.
Hommel, B. (1996). The cognitive representation of action: Automatic integration of perceived action effects. Psychological Research/Psychologische Forschung, 59, 176–186.
Hommel, B., Müsseler, J., Aschersleben, G., & Prinz, W. (2001). The theory of event coding (TEC): A framework for perception and action. Behavioral and Brain Sciences, 24, 869–937.
Iacoboni, M., Woods, R. P., Brass, M., Bekkering, H., Mazziotta, J. C., & Rizzolatti, G. (1999). Cortical mechanisms of human imitation. Science, 286, 2526–2528.
James, W. (1981/1890). The principles of psychology. Cambridge, MA: Harvard University Press.
Keele, S. W. (1968). Movement control in skilled motor performance. Psychological Bulletin, 70, 387–403.
Klapp, S. T., Porter-Graham, K. A., & Hoijfield, A. R. (1991). The relation of perception and motor action: Ideomotor compatibility and interference in divided attention. Journal of Motor Behavior, 23, 155–162.
Koch, I., & Kunde, W. (2002). Verbal response-effect compatibility. Memory and Cognition, 30, 1297–1303.
Kunde, W. (2001). Response-effect compatibility in manual choice reaction tasks. Journal of Experimental Psychology: Human Perception and Performance, 27, 387–394.
Kunde, W. (2003a). Sequential modulations of stimulus-response correspondence effects depend on awareness of response conflict. Psychonomic Bulletin & Review, 10, 198–205.
Kunde, W. (2003b). Temporal response-effect compatibility. Psychological Research, 67, 153–159.
Kunde, W., Kiesel, A., & Hoffmann, J. (2003). Conscious control over the content of unconscious cognition. Cognition, 88, 223–242.
Lien, M-C., Proctor, R. W., & Allen, P. A. (2002). Ideomotor compatibility in the psychological refractory period effect: Twenty-eight years of oversimplification. Journal of Experimental Psychology: Human Perception and Performance, 28, 396–409.
Lotze, R. H. (1852). Medizinische Psychologie oder die Physiologie der Seele. Leipzig, Germany: Weidmann’sche Buchhandlung.
Meltzoff, A. N., & Moore, M. K. (1977). Imitation of facial and manual gestures by human neonates. Science, 198, 75–78.
Neumann, O., & Klotz, W. (1994). Motor responses to nonreportable, masked stimuli: Where is the limit of direct parameter specification? In Umilta, C., Moscovitch, M. (Eds.), Attention and performance 15: Conscious and nonconscious information processing. Attention and performance series (pp. 123–150). Cambridge, MA: MIT Press.
Öhmann, A., & Soares, J. (1998). Emotional conditioning to masked stimuli: expectancies for aversive outcomes following nonrecognized fear-relevant stimuli. Journal of Experimental Psychology: General, 127, 69–82.
Ratcliff, R. (1979). Group reaction time distributions and analysis of distribution statistics. Psychological Bulletin, 86, 446–461.
Schmidt, R. A. (1975). A schema theory of discrete motor skill learning. Psychological Review, 82, 225–260.
Stürmer, B., Aschersleben, G., & Prinz, W. (2000). Correspondence effects with manual gestures and postures: A study of imitation. Journal of Experimental Psychology: Human Perception and Performance, 26, 1746–1759.
Vorberg, D., Mattler, U., Heinecke, A., Schmidt, T., & Schwarzbach, J. (2003). Invariant time-course of priming with and without awareness. In C. Kaernbach, E. Schröger, & H. Müller (Eds.). Psychophysics beyond sensation: Laws and invariants of human cognition. Hillsdale, NJ: Erlbaum.
Weeks, D. J., & Proctor, R. W. (1991). Salient-features coding in the translation between orthogonal stimulus and response dimensions. Journal of Experimental Psychology: General, 119, 355–366.
Zießler, M., & Nattkemper, D. (2002). Effect anticipation and action planning. In Prinz, W., & Hommel, B. (Eds.), Common mechanisms in perception and action: Attention & Performance XIX. Oxford: Oxford University Press.
Acknowledgements
The funding of this research was provided by a grant from the German Research Foundation to Joachim Hoffmann and Wilfried Kunde (Grant HO 1301/6-1&3). I thank Dieter Nattkemper, Michael Zießler, Ines Jentzsch, and an anonymous reviewer for helpful comments on an earlier draft of this paper, and Susanne Ebert and Karsten Toth for collecting data.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Kunde, W. Response priming by supraliminal and subliminal action effects. Psychological Research 68, 91–96 (2004). https://doi.org/10.1007/s00426-003-0147-4
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00426-003-0147-4