
Abstract
Main conclusion
This review summarizes recent advances in cyanobacterial terpenoid production. The challenges and opportunities of improving terpenoid production by cyanobacteria are discussed.
Terpenoids are a diverse group of natural products with a variety of commercial applications. With recent advances in synthetic biology and metabolic engineering, microbial terpenoid synthesis is being viewed as a feasible approach for industrial production. Among different microbial hosts, cyanobacteria have the potential of sustainable production of terpenoids using light and CO2. Terpene synthases and the precursor pathways have been expressed in cyanobacteria for enhanced production of various terpene hydrocarbons, including isoprene, limonene, β-phellandrene, and farnesene. However, the productivities need to be further improved for commercial production. Many barriers remain to be overcome in order to efficiently convert CO2 to terpenoids. In this review, we will summarize recent efforts on photosynthetic production of terpenoids and discuss the challenges and opportunities of engineering cyanobacteria for terpenoid bioproduction.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Cyanobacteria are photosynthetic microorganisms that only require sunlight, CO2, and trace minerals for growth. In recent years, cyanobacteria are emerging hosts for production of chemicals, because these photosynthetic microorganisms directly convert CO2 into desired compounds. By expressing heterologous pathways in cyanobacteria, CO2 is converted to many useful products (Sengupta et al. 2018; Knoot et al. 2018), such as ethanol (Dexter and Fu 2009), 2,3-butanediol (Kanno et al. 2017), ethylene (Ungerer et al. 2012), and fatty acids (Liu et al. 2011). Several synthetic biology tools were developed for cyanobacteria in the past decade. For instance, CRISPR/Cas9 (Li et al. 2016; Wendt et al. 2016) and Cpf1 (Ungerer and Pakrasi 2016) techniques enable rapid modification of genome sequences. Development of inducible promoters (Markley et al. 2015; Oliver et al. 2013; Huang and Lindblad 2013) and CRISPR interference (Gordon et al. 2016), and characterization of endogenous promoters (Liu and Pakrasi 2018; Li et al. 2018) allow controllable and tunable expression of heterologous genes. Currently, microbial production of fuels and chemicals is mainly performed by heterotrophic microorganisms such as Escherichia coli and Saccharomyces cerevisiae, which require sugar feedstocks for growth. However, terrestrial plants that produce sugars have a low energy efficiency to convert solar energy into biomass (< 0.5%) (Brenner et al. 2006), which become a major hurdle for sustainable feedstock production. Cyanobacteria have a higher efficiency (3–9%) than land plants to utilize solar energy (Dismukes et al. 2008). Direct conversion of CO2 to products is expected to increase the overall efficiency of production. Although cyanobacteria have been engineered to produce a variety of useful chemicals, the productivity requires to be further enhanced for commercial application. More research is needed to improve the efficiency of directing the photosynthetic carbon toward desired chemicals.
Terpenoids: functions, applications, and biosynthesis
Terpenoids (or isoprenoids) are the largest group of natural products with extreme diversity. More than 55,000 compounds have been identified (Breitmaier 2006). Some terpenoids are essential in primary metabolism, including photosynthesis (chlorophyll, carotenoids, and plastoquinone), respiration (ubiquinone), developmental regulation in plants (gibberellins and abscisic acid), etc. Most terpenoids are produced by plants as secondary metabolites. Isoprene is a volatile molecule emitted from numerous plant species to protect against heat stress. Isoprene enhances membrane integrity for the purpose of thermotolerance (Siwko et al. 2007), because heat can cause leakiness of thylakoid membrane and affect the efficiency of photosynthesis (Sharkey et al. 2008). Some volatile terpenoids emit floral scents, which can attract pollinators and seed-dispersing animals to facilitate plant reproduction. Moreover, plants release terpenoids with toxicity or strong odor to protect against herbivores and pathogens.
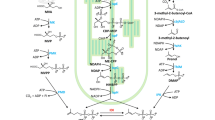
Although terpenoids are extremely diverse compounds, they are all derived from the same building blocks, isopentenyl pyrophosphate (IPP) and dimethylallyl pyrophosphate (DMAPP) (Fig. 1). One of the simplest types of terpenoids is isoprene, which is a five-carbon compound converted from DMAPP. Terpenoids are categorized into groups based on the number of isoprene units: hemiterpenes (C5), monoterpenes (C10), sesquiterpenes (C15), diterpenes (C20), triterpenes (C30), tetraterpenes (C40), and polyterpenes (> C40). In terms of nomenclature, terpenes are the hydrocarbon molecules, whereas terpenoids are terpenes modified with additional functional groups or structural rearrangement. However, the two terms are often used interchangeably.

Metabolic pathway of terpenoid biosynthesis. In cyanobacteria, linear prenyl pyrophosphates (GPP, FPP, and GGPP) are sequentially synthesized by CrtE. Terpenoid production catalyzed by terpene synthases can be from one-step or multiple-step reactions. Some of the commercially used terpenoids are shown here. GAP glyceraldehyde 3-phosphate, DXP 1-deoxy-d-xylulose 5-phosphate, DXS DXP synthase, MEP 2-C-methyl-d-erythritol-4-phosphate, DXR DXP reductase, CDP-ME 4-diphosphocytidyl-2-C-methyl-d-erythritol, IspD 2-C-methyl-d-erythritol 4-phosphate cytidyltransferase, CDP-MEP 4-diphosphocytidyl-2-C-methyl-d-erythritol 2-phosphate, IspE 4-diphosphocytidyl-2-C-methyl-d-erythritol kinase, MEcPP 2-C-methyl-d-erythritol 2,4-cyclodiphosphate, IspF 2-C-methyl-d-erythritol 2,4-cyclodiphosphate synthase, HMP-PP 4-hydroxy-3-methyl-butenyl 1-diphosphate, IspG HMB-PP synthase, IspH HMB-PP reductase, AtoB acetoacetyl-CoA thiolase, HMG-CoA 3-hydroxy-3-methyl-glutaryl-CoA, HMGS HMG-CoA synthase, HMGR HMG-CoA reductase, MK mevalonate kinase, PMK mevalonate 5-phosphate kinase, PMD mevalonate 5-pyrophosphate decarboxylase, IPP isopentenyl pyrophosphate, DMAPP dimethylallyl pyrophosphate, IDI isopentenyl diphosphate isomerase, IspS isoprene synthase, GPP geranyl pyrophosphate, GPPS GPP synthase, FPP farnesyl pyrophosphate, FPPS FPP synthase, GGPP geranylgeranyl pyrophosphate, GGPPS GGPP synthase, TPS terpene synthase
Terpenoids have a variety of commercial applications, including pharmaceuticals, nutraceuticals, flavors/fragrances, and industrial chemicals (Ajikumar et al. 2008). Artemisinin is a sesquiterpene produced by the plant Artemisia annua. Artemisinin derivatives are the first-line antimalarial drugs recommended by World Health Organization. The price and availability of artemisinin fluctuate wildly due to unstable supply of agricultural materials. With recent improvement in synthetic biology and metabolic engineering, the yeast S. cerevisiae was engineered for commercial production of artemisinic acid, the precursor of artemisinin (Paddon and Keasling 2014). Paclitaxol, a complex diterpene isolated from the Pacific yew tree (Taxus brevifolia), is a chemotherapy drug for treatment of several types of cancer. However, the low yield of paclitaxol extracted from the bark of the yew tree (< 0.01% of dry weight) is a major barrier for large-scale production (Ciddi et al. 1995). Many terpenoids are used as nutraceuticals due to their health effects to human, such as lutein, lycopene, and astaxanthin. A wide variety of monoterpenes and sesquiterpenes have characteristic fragrances which are used in the flavor and fragrance industry, such as menthol (mint odor), d-limonene (orange peel odor), α-farnesene (green apple odor), and nootkatone (grapefruit odor). Isoprene, the simplest form of terpenoid, is the main component of synthetic and natural rubber. Recently, some terpenoids (limonene, myrcene, and farnesene) or their hydrogenated forms have been determined to be compatible with diesel (Tracy et al. 2009) and aviation fuels (Chuck and Donnelly 2014), demonstrating the potential of using these terpenoids as alternative fuels.
Two distinct metabolic pathways can lead to the synthesis of IPP and DMAPP (Fig. 1). In general, the 2-C-methyl-d-erythritol 4-phosphate (MEP) pathway (also known as the 1-deoxy-d-xylulose 5-phosphate (DXP) pathway or the non-mevalonate pathway) exists in bacteria, whereas the mevalonate (MVA) pathway is mainly found in archaea and eukaryotes. Plants and algae have both pathways, in which the MVA pathway operates in the cytosol and the MEP pathway functions in the plastids. The MEP pathway consists of seven enzymatic reactions. It begins with glyceraldehyde 3-phosphate (GAP) and pyruvate to form DXP. This step is catalyzed by the rate-limiting enzyme, DXP synthase (DXS). Further, DXP is converted to MEP by DXP reductase (DXR), which requires NADPH as reducing power. Subsequently, MEP is coupled with CTP to form 4-diphosphocytidyl-2-C-methyl-d-erythritol (CDP-ME), which further undergoes phosphorylation, cyclization, and reductive dehydration to form 4-hydroxy-3-methyl-butenyl 1-diphosphate (HMB-PP). Finally, the HMB-PP is converted to IPP and DMAPP by HMB-PP reductase (IspH). The enzyme isopentenyl diphosphate isomerase (IDI) interconverts IPP and DMAPP. Although it exists in the MEP pathway, it is not essential because IspH can generate both IPP and DMAPP. The MVA pathway also contains seven reaction steps. It initiates with two condensation steps, in which three molecules of acetyl-CoA are coupled to form 3-hydroxy-3-methyl-glutaryl-CoA (HMG-CoA). Afterward, HMG-CoA is converted to mevalonate, which is then phosphorylated twice to form mevalonate pyrophosphate. IPP is generated by decarboxylation of mevalonate pyrophosphate, and the IDI catalyzes isomerization of IPP to form DMAPP.
Besides their substrate requirements, the MEP and MVA pathways differ in energy utilization and carbon yield. The net reactions of both pathways for synthesizing IPP from GAP are summarized as below:
The MEP pathway utilizes carbon more efficiently, in which 83% (5/6) of the carbon substrate is used for IPP production, whereas the MVA pathway has 56% (5/9) carbon utilization. Conversely, the MVA pathway is more efficient at energy generation, in which three ATP and three NAD(P)H are produced from three molecules of GAP, whereas the MEP pathway generates one ATP from two molecules of GAP.
The downstream process of terpene synthesis begins with sequential head-to-tail condensation of DAMPP with IPP to generate linear prenyl pyrophosphates, including geranyl pyrophosphate (GPP, C10), farnesyl pyrophosphate (FPP, C15), geranylgeranyl pyrophosphate (GGPP, C20), etc. (Fig. 1). Further, terpene synthases convert the pyrophosphate intermediates into various terpenes. Notably, the photosynthetic pigments such as carotenoids and the phytol tail on chlorophyll are synthesized from this metabolic pathway. Therefore, the terpene biosynthesis pathway is critically important for cyanobacteria.
Recent advances in cyanobacterial terpenoid production
Engineering the terpene synthases
To engineer cyanobacteria for terpenoid production, the choice of terpene synthase is critical. An ideal enzyme should exhibit superior catalytic rate and good solubility in the host cells. The abundance of a soluble enzyme and its activity are correlated with the production titer (Gao et al. 2016). For isoprene production, the commonly used isoprene synthases (ispS) are derived from Populus alba and Pueraria montana (also known as kudzu). Recently, the ispS genes from various high isoprene emission plant species were expressed in Synechococcus elongatus PCC 7942 (hereafter Synechococcus 7942). The IspS from Eucalyptus globulus produced the highest amount of isoprene, over 5- and 25-fold higher than that from P. alba and P. montana, respectively (Gao et al. 2016). Similarly, a recent work reported that IspS from E. globulus and Ipomoea batatas increased isoprene production more than 20 fold compared to the P. montana IspS (Englund et al. 2018). The monoterpene limonene is widely found in plants. The limonene synthase (Lims) from Mentha spicata is the commonly used enzyme for limonene production. It exhibits the highest production level than that from other plant species (Table 1).
Another feature of terpene synthases that needs to be taken into account is enzyme promiscuity. Many terpene synthases produce multiple products. For instance, limonene synthase from Mentha spicata primarily produces S-limonene with trace amounts of α-pinene, β-pinene, and myrcene (Colby et al. 1993). Pinene synthase from Citrus limon mainly generates β-pinene but also α-pinene, sabinene, and limonene (Lucker et al. 2002). Expressing a terpene synthase with a higher product specificity prevents the formation of unwanted products (Lin et al. 2017).
To enhance expression of terpene synthase, strong promoters such as PcpcB and Ptrc promoters are widely used to drive gene expression in cyanobacteria (Lin et al. 2017; Wang et al. 2016; Kiyota et al. 2014; Davies et al. 2014). A recent work attempted stepwise metabolic engineering of Synechococcus 7942 for limonene production, while obtaining a marginal productivity increase (Wang et al. 2016). Surprisingly, by replacing the Ptrc promoter by the pea PpsbA promoter for Lims expression, the production titer increased over 100-fold (Wang et al. 2016). Proteomic analysis confirmed the increased expression of Lims by the PpsbA promoter in the host strain (Wang et al. 2016). In addition, protein fusion is a strategy to increase protein expression. Using the highly expressed c-phycocyanin beta subunit (CpcB) protein as a leader sequence, the CpcB–IspS fusion constructs led to a significant improvement in protein abundance and isoprene productivity (Chaves et al. 2017). Although the IspS activity was attenuated, the increased expression level compensated the reduced activity of IspS, resulting in a 27-fold increase in isoprene production (Chaves et al. 2017).
Unlike plants, cyanobacteria do not have specific prenyltransferases to generate GPP and FPP, substrates for monoterpene and sesquiterpene synthases, respectively. These diphosphate intermediates are synthesized sequentially by the enzyme CrtE, which is a GGPP synthase (Fig. 1). A common strategy for enhancing monoterpene and sesquiterpene production is to express specific GPP and FPP synthases, respectively. Such strategy increases the amounts of GPP and FPP for terpenoid production (Lin et al. 2017; Wang et al. 2016; Choi et al. 2016; Englund et al. 2015). However, expression of these prenyltransferases reduces the carbon flux toward pigment synthesis, lowering the amount of photosynthetic pigments in cyanobacteria (Lin et al. 2017; Choi et al. 2016; Englund et al. 2015).
Engineering the precursor pathways
To enhance production of terpenoids, it is critical to increase the amount of the precursors (IPP and DMAPP), which can be achieved by engineering the MEP or MVA pathways. In E. coli, it is widely known that the DXS and IDI catalyze the bottleneck reactions in the MEP pathway. Overexpression of genes encoding the DXS and IDI leads to significant increase in terpene production, and this has been summarized in a recent review (Niu et al. 2017). Moreover, a previous study extensively engineered the MEP pathway in E. coli to produce taxadiene, the precursor of the potent anticancer drug paclitaxol. By overexpressing and optimizing four bottleneck genes (dxs, ispD, ispF, and idi) in the MEP pathway, the optimized strain increased taxadiene production 15,000-fold, leading to 1 g L−1 yield in fed-batch bioreactors (Ajikumar et al. 2010).
To increase the terpene precursors in cyanobacteria, most studies focused on optimizing the native MEP pathway. A recent study engineered Synechococcus 7942 for isoprene production. Overexpressing the native dxs gene increased 20% of the isoprene yield (Gao et al. 2016). Similarly, production of limonene was enhanced by overexpressing the dxs gene together with other genes involved in the upstream pathway. A previous work overexpressed genes encoding the DXS (from E. coli), IDI (from Haematococcus pluvialis), and GPPS (from Mycoplasma tuberculosis) in Anabaena sp. PCC 7120, resulting in a 6.8-fold increase in limonene production (0.52 mg L−1 in 12 days) (Halfmann et al. 2014b). By expressing the same set of genes (dxs, idi, and gpps from Synechocystis sp. PCC 6803), the limonene yield in Synechocystis 6803 increased 40% to 1 mg L−1 in 30 days (Kiyota et al. 2014). The productivity of limonene in Synechococcus 7942 was increased 2.3-fold to 76.3 μg L−1 OD −1730 day−1 by overexpressing the dxs gene from Botryococcus braunii and the gpps gene from Abies grandis (Wang et al. 2016). Moreover, increased production of the plant diterpenoid manoyl oxide was achieved by expressing the dxs gene from the plant Coleus forskohlii, which resulted in a 4.2-fold improvement in low light condition (20 μmol photons m−2 s−1) (Englund et al. 2015). However, the dxs-overexpressing strain showed no improvement in production under high light condition (100 μmol photons m−2 s−1) (Englund et al. 2015).
Compared to DXS, IDI seems to be a more important target for enhancing isoprene production in cyanobacteria. Synechococcus 7942 has a low ratio of DMAPP/IPP concentration (i.e., 0.03 in an isoprene-producing strain) (Gao et al. 2016). Such low ratio may affect the substrate availability for isoprene production, since DMAPP is the substrate for isoprene synthase. In addition, the isoprene-emitting kudzu leaves have a high DMAPP/IPP ratio (i.e., > 2) (Zhou et al. 2013), indicating that isoprene production is related to DMAPP availability. To increase the DMAPP/IPP ratio, the idi gene from S. cerevisiae was overexpressed in an isoprene-producing Synechococcus 7942, which increased the DMAPP/IPP ratio to 2.25 and resulted in a twofold improvement in isoprene production (Gao et al. 2016). However, co-expression of dxs and idi genes produced similar amount of isoprene compared to the idi-expressing strain (Gao et al. 2016), suggesting that the IDI has a more critical role than DXS for enhancing isoprene production in cyanobacteria.
Besides DXS and IDI catalyzing the critical reactions for terpene synthesis, IspD and IspG were recently identified as bottleneck enzymes of the MEP pathway in cyanobacteria. Systematic overexpression of each enzyme in the MEP pathway identified IspD as the third enzyme for enhanced isoprene production in Synechocystis 6803 (Englund et al. 2018). Kinetic flux profiling showed that the flux through the reaction catalyzed by IspG was very low in Synechococcus 7942 (Gao et al. 2016). Overexpressing the ispG gene from Thermosynechococcus elongatus increased isoprene production 60% (Gao et al. 2016). The final strain in the study, with expression of ispS, dxs, idi, and ispG, produced 1.26 g L−1 isoprene in a photobioreactor. The productivity was 102 mg L−1 day−1 in the first 9 days, which is over two orders of magnitude higher than all the reported literature on cyanobacterial terpenoid production (Table 1).
Another strategy to increase terpene precursors is to express the heterologous MVA pathway, which is widely used for terpene production in E. coli. Introduction of this pathway increases the carbon flux toward the terpene precursors, which further enhances productivity of various terpenoids such as isoprene, lycopene, limonene, and farnesene (Niu et al. 2017). With cyanobacteria, the Melis group successfully introduced the heterologous MVA pathway into Synechocystis 6803 for enhanced production of isoprene (Bentley et al. 2014) and β-phellandrene (Formighieri and Melis 2016). Unlike previous work in E. coli which expressed the MVA pathway from S. cerevisiae, the upper MVA pathway in their studies was derived from Enterococcus faecalis (HMGS, HMGR) and E. coli (AtoB), whereas the lower MVA pathway was obtained from Streptococcus pneumoniae (MK, PMK, PMD, and IDI) (Formighieri and Melis 2016; Bentley et al. 2014). Both pathways were successfully introduced into the genome of Synechocystis 6803, and all enzymes were actively expressed (Bentley et al. 2014). Expressing the MVA pathway increased terpene production over twofold (Formighieri and Melis 2016; Bentley et al. 2014), suggesting that the heterologous pathway indeed directs an extra carbon flux toward terpene synthesis. However, such improvement was less impactful than previous efforts in E. coli. For instance, expression of the S. cerevisiae MVA pathway increased artemisinic acid production 36 fold (Martin et al. 2003). Also, limonene production improved nearly 90 fold by optimizing the MVA pathway from Staphylococcus aureus and S. cerevisiae (Alonso-Gutierrez et al. 2013). The heterologous MVA pathway used in E. coli has been extensively studied and perhaps this optimized pathway can be expressed in cyanobacteria to see if terpene production could be further improved.
Recently, the pentose phosphate (PP) pathway in cyanobacteria was engineered for enhanced production of limonene. Lin et al. (2017) used the Optforce computational algorithm to identify two genes in the PP pathway, ribose 5-phosphate isomerase (rpi) and ribulose 5-phosphate 3-epimerase (rpe) that are important for limonene synthesis. The limonene titer increased 2.3 fold to 6.7 mg L−1 by expressing the rpi and rpe genes from E. coli and the gpps gene from Abies grandis.
Challenges in enhancing terpenoid production in cyanobacteria
Redirection of photosynthetic carbon
One of the major challenges to enhance terpenoid production by cyanobacteria is carbon partitioning in photosynthesis. Most of the fixed carbon is directed toward sugar phosphates for biomass instead of the products. The sugar biosynthetic pathways in Synechocystis 6803 consumes more than 80% of the fixed carbon for biomass accumulation, whereas approximately 5% of the photosynthetic carbon is allocated to terpene biosynthesis, including carotenoids, phytol groups of chlorophyll, and prenyl tails of quinone (Melis 2013). The low carbon flux to the terpene metabolic pathway results in the low yield of heterologous terpenoid production.
A common strategy for redirecting the carbon flux is to remove the competing pathway(s) of desired products. In cyanobacteria, glycogen serves as a major carbon sink for energy storage under nutrient starvation conditions, in which the carbon flux is diverted to glycogen accumulation instead of cellular growth (Hickman et al. 2013; Yoo et al. 2007). Several studies attempted to delete the glycogen biosynthetic pathway for the purpose of redirecting the fixed carbon toward desired products. A recent study reported a two-fold increase in lactic acid production using a glycogen deficient Synechocystis 6803 mutant (van der Woude et al. 2014). Furthermore, overexpressing a heterologous isobutanol pathway served as an alternative carbon sink that rescued the growth phenotype of a Synechococcus 7942 glycogen mutant, and increased the carbon flux to isobutanol synthesis (Li et al. 2014). However, inactivation of glycogen sinks leads to metabolic imbalance under nitrogen-deplete conditions, in which organic acids such as pyruvate and α-ketoglutarate are excreted from the cells (Davies et al. 2014; Hickman et al. 2013; Carrieri et al. 2012). Unfortunately, removing the glycogen synthesis pathway did not lead to improved terpenoid production in cyanobacteria. Davies et al. (2014) constructed a glycogen-deficient mutant for limonene and bisabolene production. However, the production titer was extremely low due to the absence of growth under nitrogen starvation.
Compared to other compounds produced by engineered cyanobacteria, the productivities of terpenes are relatively low, even though many efforts have been done to increase terpene production. Presumably, the MEP pathway in cyanobacteria is highly regulated so that limited amount of the fixed carbon is directed to terpene synthesis. The terpene precursors IPP and DMAPP are distant from the central carbon metabolism. Quantitative analysis reveals that the concentrations of IPP, DMAPP, and other intermediates (MEP and HMB-PP) in the MEP pathway are three orders of magnitude lower than pyruvate in cyanobacteria (Dempo et al. 2014). This indicates that the sparse amount of precursor pools may be a limiting factor for enhancing terpene synthesis. A previous study using cell extracts of Synechocystis 6803 demonstrated that terpene production was increased by compounds in the PP pathway but not by substrates (pyruvate and GAP) of the MEP pathway (Ershov et al. 2002). In addition, expression of the rpi and rpe genes in the PP pathway led to increased production of limonene (Lin et al. 2017). These evidences indicate a connection between the PP pathway and terpene synthesis in cyanobacteria. Moreover, a recent study determined that the E. coli enzymes YajO and RibB (G108S) are able to convert ribulose 5-phosphate to DXP, the first intermediate in the MEP pathway (Kirby et al. 2015). Since cyanobacteria have a relatively high carbon flux in the PP pathway (Abernathy et al. 2017; Young et al. 2011), this novel route can be investigated in order to avoid feedback regulation in the MEP pathway (Banerjee et al. 2013) and direct the carbon flux from the PP pathway to terpene production.
MEP vs. MVA pathways: which is better for cyanobacterial terpenoid production?
Overexpression of the heterologous MVA pathway is a common strategy to improve terpenoid production in heterotrophic microorganisms such as E. coli and S. cerevisiae. Since the native MEP pathway is highly regulated (Banerjee and Sharkey 2014), the non-native MVA pathway is able to avoid transcriptional regulation in the hosts and enable a bypass of the carbon flux to terpene synthesis. Previous studies on cyanobacterial terpenoid production were mainly focused on the MEP pathway. A few studies expressed the heterologous MVA pathway for enhanced terpene synthesis (Table 1). This may indicate the difficulty of introducing the MVA pathway into photosynthetic microorganisms. The MVA pathway contains seven genes, about nine kilobases in length (Bentley et al. 2014). Introducing the entire pathway into cyanobacterial genome may be challenging. Although the heterologous MVA pathway has been successfully expressed in Synechocystis 6803 (Formighieri and Melis 2016; Bentley et al. 2014), the production titer is similar to other studies engineering the MEP pathway (Table 1). The MEP and MVA pathways use different substrates. The former uses GAP and pyruvate, whereas the later requires acetyl-CoA. Isotopically nonstationary 13C flux analyses showed that cyanobacteria have a relatively low flux toward acetyl-CoA compared to the sugar phosphate pathways (Abernathy et al. 2017; Young et al. 2011). Moreover, a recent study compared the precursor pools of the MEP and MVA pathways between Synechococcus 7942 and E. coli BL-21, and found that the concentrations of GAP and pyruvate in Synechococcus 7942 were 5- and 21-fold higher than that in E. coli, whereas the acetyl-CoA pool was 20-fold lower (Gao et al. 2016). These data indicate that expressing the MVA pathway in cyanobacteria may be less effective than in E. coli for enhanced terpene production. Hence, the native MEP pathway is presumably a more attractive target for metabolic engineering, even though further research is needed to elucidate the regulation of the MEP pathway in cyanobacteria. It is known that overexpressing the enzymes in the MEP pathway can reduce terpenoid production in E. coli (Ajikumar et al. 2010; Kim and Keasling 2001). The expression level needs to be optimized in order to maximize the productivity. Despite many efforts to engineer the MEP pathway in cyanobacteria, optimization of the pathway is yet to be attempted. This can be achieved by modifying the ribosome binding site (RBS) (Wang et al. 2018; Oliver et al. 2014) or using inducible promoters (Markley et al. 2015; Oliver et al. 2013; Huang and Lindblad 2013) to modulate gene expression.
Balancing pigment and product synthesis
The photosynthetic pigments such as chlorophyll and carotenoids are essential for cyanobacteria. Production of terpenes will necessarily compete with pigment synthesis because the same precursor pathway is used. Although the expression of prenyltransferases such as GPP and FPP synthases is critical in enhancing terpenoid production (Lin et al. 2017; Choi et al. 2016), the carbon flux for photosynthetic pigments is diverted toward the terpene product. Previous studies reported that photosynthetic pigment levels were lowered in terpene-producing cyanobacteria (Lin et al. 2017; Choi et al. 2016; Englund et al. 2015). Decreased amount of pigments may affect the efficiency of light harvesting and photoprotection and alter the structural stability of photosynthetic complexes (Toth et al. 2015), thus leading to impaired cellular growth and terpene production. To further increase terpenoid production in cyanobacteria, the carbon fluxes between the product and pigment synthesis should be balanced in addition to increasing the terpene precursors. Modulating the expression of prenyltransferases could presumably optimize carbon fluxes and improve terpene synthesis. A recent study reported that the expression level of GPP synthase is critically important for enhancing monoterpene production in E. coli. Optimization of GPP synthase expression by engineering the RBS sequence led to a sixfold improvement in geraniol production (Zhou et al. 2015).
Future perspectives
Cyanobacterial production of terpenoids demonstrates the potential of sustainable bioproduction using light and CO2, although further research is needed to enhance the productivity. Challenges including the regulation of MEP pathway, carbon partitioning, and balancing pigment and limonene synthesis should be addressed collectively in order to further increase the productivity of terpenoids. With many obstacles required to be overcome, recent developments on the genetic tools for cyanobacteria (Sengupta et al. 2018) can facilitate strain construction and accelerate the design-build-test cycle, which will benefit systematic engineering of cyanobacteria. The advantage of using cyanobacteria as a production platform is direct conversion of CO2 to desired compounds without the use of sugar feedstocks. However, the relatively slow rates of growth and CO2 fixation of standard laboratory strains presumably lead to low productivity of chemicals. Particularly, the small fraction of fixed carbon allocated to terpene metabolism makes terpenoid productivity even lower than other compounds. To utilize cyanobacteria for commercial bioproduction, a better chassis is needed to achieve higher production titer and productivity. Synechococcus elongatus UTEX 2973 is a fast-growing cyanobacterium with the fastest growth rate reported to date (Yu et al. 2015). The doubling time of this strain can be as fast as 1.5 h under high light and CO2 conditions (Ungerer et al. 2018), which is comparable to the heterotrophic growth rate of the yeast S. cerevisiae. The fast growth of Synechococcus 2973 reflects the rapid accumulation of biomass, at a rate of over 1 g L−1 day−1 (Ungerer et al. 2018). Moreover, this strain synthesizes significant amount of glycogen as a way to reserve excess fixed carbon, with over 35% of dry cell weight after 3 days of growth (Ungerer et al. 2018). Recently, Synechococcus 2973 was engineered for sugar production. The productivity of sucrose reaches 0.85 g L−1 day−1 (the highest amount from photoautotrophic production), demonstrating the potential of using this strain for biotechnology applications (Song et al. 2016). With recent advances in metabolic engineering of terpene biosynthetic pathway, this fast-growing cyanobacterium offers an opportunity to improve the productivity of terpenoids.
Author contribution statement
PCL compiled the literature, wrote the manuscript, and composed the figure. HBP wrote and edited the manuscript and completed the literature.
Abbreviations
- DMAPP:
-
Dimethylallyl pyrophosphate
- DXS:
-
1-Deoxy-d-xylulose 5-phosphate synthase
- GAP:
-
Glyceraldehyde 3-phosphate
- IDI:
-
Isopentenyl diphosphate isomerase
- IPP:
-
Isopentenyl pyrophosphate
- IspS:
-
Isoprene synthase
- MEP:
-
2-C-Methyl-d-erythritol-4-phosphate
- MVA:
-
Mevalonate
References
Abernathy MH, Yu J, Ma F, Liberton M, Ungerer J, Hollinshead WD, Gopalakrishnan S, He L, Maranas CD, Pakrasi HB, Allen DK, Tang YJ (2017) Deciphering cyanobacterial phenotypes for fast photoautotrophic growth via isotopically nonstationary metabolic flux analysis. Biotechnol Biofuels 10:273. https://doi.org/10.1186/s13068-017-0958-y
Ajikumar PK, Tyo K, Carlsen S, Mucha O, Phon TH, Stephanopoulos G (2008) Terpenoids: opportunities for biosynthesis of natural product drugs using engineered microorganisms. Mol Pharm 5(2):167–190
Ajikumar PK, Xiao WH, Tyo KE, Wang Y, Simeon F, Leonard E, Mucha O, Phon TH, Pfeifer B, Stephanopoulos G (2010) Isoprenoid pathway optimization for Taxol precursor overproduction in Escherichia coli. Science 330(6000):70–74. https://doi.org/10.1126/science.1191652
Alonso-Gutierrez J, Chan R, Batth TS, Adams PD, Keasling JD, Petzold CJ, Lee TS (2013) Metabolic engineering of Escherichia coli for limonene and perillyl alcohol production. Metab Eng 19:33–41. https://doi.org/10.1016/j.ymben.2013.05.004
Banerjee A, Sharkey T (2014) Methylerythritol 4-phosphate (MEP) pathway metabolic regulation. Nat Prod Rep 31(8):1043–1055
Banerjee A, Wu Y, Banerjee R, Li Y, Yan H, Sharkey TD (2013) Feedback inhibition of deoxy-d-xylulose-5-phosphate synthase regulates the methylerythritol 4-phosphate pathway. J Biol Chem 288(23):16926–16936. https://doi.org/10.1074/jbc.M113.464636
Bentley FK, Melis A (2012) Diffusion-based process for carbon dioxide uptake and isoprene emission in gaseous/aqueous two-phase photobioreactors by photosynthetic microorganisms. Biotechnol Bioeng 109(1):100–109
Bentley FK, Zurbriggen A, Melis A (2014) Heterologous expression of the mevalonic acid pathway in cyanobacteria enhances endogenous carbon partitioning to isoprene. Mol Plant 7(1):71–86. https://doi.org/10.1093/mp/sst134
Betterle N, Melis A (2018) Heterologous leader sequences in fusion constructs enhance expression of geranyl diphosphate synthase and yield of β-phellandrene production in cyanobacteria (Synechocystis). ACS Synth Biol 7(3):912–921
Breitmaier E (2006) Terpenes: importance, general structure, and biosynthesis. In: Terpenes, chap 1. Wiley, Weinheim
Brenner MP, Bildsten L, Dyson F, Fortson N, Garwin R, Grober R, Hemley R, Hwa T, Joyce G, Katz J (2006) Engineering microorganisms for energy production. DTIC Document
Carrieri D, Paddock T, Maness P-C, Seibert M, Yu J (2012) Photo-catalytic conversion of carbon dioxide to organic acids by a recombinant cyanobacterium incapable of glycogen storage. Energy Environ Sci 5(11):9457–9461
Chaves JE, Rueda-Romero P, Kirst H, Melis A (2017) Engineering isoprene synthase expression and activity in cyanobacteria. ACS Synth Biol 6(12):2281–2292. https://doi.org/10.1021/acssynbio.7b00214
Choi SY, Lee HJ, Choi J, Kim J, Sim SJ, Um Y, Kim Y, Lee TS, Keasling JD, Woo HM (2016) Photosynthetic conversion of CO 2 to farnesyl diphosphate-derived phytochemicals (amorpha-4, 11-diene and squalene) by engineered cyanobacteria. Biotechnol Biofuels 9(1):202
Chuck CJ, Donnelly J (2014) The compatibility of potential bioderived fuels with Jet A-1 aviation kerosene. Appl Energy 118:83–91
Ciddi V, Srinivasan V, Shuler M (1995) Elicitation of Taxus sp. cell cultures for production of taxol. Biotechnol Lett 17(12):1343–1346
Colby SM, Alonso WR, Katahira EJ, McGarvey DJ, Croteau R (1993) 4S-limonene synthase from the oil glands of spearmint (Mentha spicata). cDNA isolation, characterization, and bacterial expression of the catalytically active monoterpene cyclase. J Biol Chem 268(31):23016–23024
Davies FK, Work VH, Beliaev AS, Posewitz MC (2014) Engineering limonene and bisabolene production in wild type and a glycogen-deficient mutant of Synechococcus sp. PCC 7002. Front Bioeng Biotechnol 2:21. https://doi.org/10.3389/fbioe.2014.00021
Dempo Y, Ohta E, Nakayama Y, Bamba T, Fukusaki E (2014) Molar-based targeted metabolic profiling of cyanobacterial strains with potential for biological production. Metabolites 4(2):499–516. https://doi.org/10.3390/metabo4020499
Dexter J, Fu P (2009) Metabolic engineering of cyanobacteria for ethanol production. Energy Environ Sci 2(8):857–864
Dismukes GC, Carrieri D, Bennette N, Ananyev GM, Posewitz MC (2008) Aquatic phototrophs: efficient alternatives to land-based crops for biofuels. Curr Opin Biotechnol 19(3):235–240. https://doi.org/10.1016/j.copbio.2008.05.007
Englund E, Andersen-Ranberg J, Miao R, Hamberger B, Lindberg P (2015) Metabolic engineering of Synechocystis sp. PCC 6803 for production of the plant diterpenoid manoyl oxide. ACS Synth Biol 4(12):1270–1278. https://doi.org/10.1021/acssynbio.5b00070
Englund E, Shabestary K, Hudson EP, Lindberg P (2018) Systematic overexpression study to find target enzymes enhancing production of terpenes in Synechocystis PCC 6803, using isoprene as a model compound. Metab Eng 49:164–177. https://doi.org/10.1016/j.ymben.2018.07.004
Ershov YV, Gantt RR, Cunningham FX Jr, Gantt E (2002) Isoprenoid biosynthesis in Synechocystis sp. strain PCC6803 is stimulated by compounds of the pentose phosphate cycle but not by pyruvate or deoxyxylulose-5-phosphate. J Bacteriol 184(18):5045–5051
Formighieri C, Melis A (2015) A phycocyanin phellandrene synthase fusion enhances recombinant protein expression and β-phellandrene (monoterpene) hydrocarbons production in Synechocystis (cyanobacteria). Metab Eng 32:116–124
Formighieri C, Melis A (2016) Sustainable heterologous production of terpene hydrocarbons in cyanobacteria. Photosynth Res 130(1–3):123–135. https://doi.org/10.1007/s11120-016-0233-2
Gao X, Gao F, Liu D, Zhang H, Nie X, Yang C (2016) Engineering the methylerythritol phosphate pathway in cyanobacteria for photosynthetic isoprene production from CO2. Energy Environ Sci 9(4):1400–1411
Gordon GC, Korosh TC, Cameron JC, Markley AL, Begemann MB, Pfleger BF (2016) CRISPR interference as a titratable, trans-acting regulatory tool for metabolic engineering in the cyanobacterium Synechococcus sp. strain PCC 7002. Metab Eng 38:170–179
Halfmann C, Gu L, Gibbons W, Zhou R (2014a) Genetically engineering cyanobacteria to convert CO2, water, and light into the long-chain hydrocarbon farnesene. Appl Microbiol Biotechnol 98(23):9869–9877
Halfmann C, Gu L, Zhou R (2014b) Engineering cyanobacteria for the production of a cyclic hydrocarbon fuel from CO2 and H2O. Green Chem 16(6):3175–3185
Hickman JW, Kotovic KM, Miller C, Warrener P, Kaiser B, Jurista T, Budde M, Cross F, Roberts JM, Carleton M (2013) Glycogen synthesis is a required component of the nitrogen stress response in Synechococcus elongatus PCC 7942. Algal Res 2(2):98–106
Huang HH, Lindblad P (2013) Wide-dynamic-range promoters engineered for cyanobacteria. J Biol Eng 7(1):10. https://doi.org/10.1186/1754-1611-7-10
Kanno M, Carroll AL, Atsumi S (2017) Global metabolic rewiring for improved CO2 fixation and chemical production in cyanobacteria. Nat Commun 8:14724. https://doi.org/10.1038/ncomms14724
Kim SW, Keasling JD (2001) Metabolic engineering of the nonmevalonate isopentenyl diphosphate synthesis pathway in Escherichia coli enhances lycopene production. Biotechnol Bioeng 72(4):408–415
Kirby J, Nishimoto M, Chow RW, Baidoo EE, Wang G, Martin J, Schackwitz W, Chan R, Fortman JL, Keasling JD (2015) Enhancing terpene yield from sugars via novel routes to 1-deoxy-d-xylulose 5-phosphate. Appl Environ Microbiol 81(1):130–138. https://doi.org/10.1128/AEM.02920-14
Kiyota H, Okuda Y, Ito M, Hirai MY, Ikeuchi M (2014) Engineering of cyanobacteria for the photosynthetic production of limonene from CO2. J Biotechnol 185:1–7. https://doi.org/10.1016/j.jbiotec.2014.05.025
Knoot CJ, Ungerer J, Wangikar PP, Pakrasi HB (2018) Cyanobacteria: promising biocatalysts for sustainable chemical production. J Biol Chem 293(14):5044–5052
Li X, Shen CR, Liao JC (2014) Isobutanol production as an alternative metabolic sink to rescue the growth deficiency of the glycogen mutant of Synechococcus elongatus PCC 7942. Photosynth Res 120(3):301–310. https://doi.org/10.1007/s11120-014-9987-6
Li H, Shen CR, Huang C-H, Sung L-Y, Wu M-Y, Hu Y-C (2016) CRISPR-Cas9 for the genome engineering of cyanobacteria and succinate production. Metab Eng 38:293–302
Li S, Sun T, Xu C, Chen L, Zhang W (2018) Development and optimization of genetic toolboxes for a fast-growing cyanobacterium Synechococcus elongatus UTEX 2973. Metab Eng 48:163–174
Lin P-C, Saha R, Zhang F, Pakrasi HB (2017) Metabolic engineering of the pentose phosphate pathway for enhanced limonene production in the cyanobacterium Synechocystis sp. PCC 6803. Sci Rep 7(1):17503. https://doi.org/10.1038/s41598-017-17831-y
Liu D, Pakrasi HB (2018) Exploring native genetic elements as plug-in tools for synthetic biology in the cyanobacterium Synechocystis sp PCC 6803. Microb Cell Fact 17(1):48
Liu X, Sheng J, Curtiss R III (2011) Fatty acid production in genetically modified cyanobacteria. Proc Natl Acad Sci 108:6899–6904
Lucker J, El Tamer MK, Schwab W, Verstappen FW, van der Plas LH, Bouwmeester HJ, Verhoeven HA (2002) Monoterpene biosynthesis in lemon (Citrus limon). cDNA isolation and functional analysis of four monoterpene synthases. Eur J Biochem FEBS 269(13):3160–3171
Markley AL, Begemann MB, Clarke RE, Gordon GC, Pfleger BF (2015) Synthetic biology toolbox for controlling gene expression in the cyanobacterium Synechococcus sp. strain PCC 7002. ACS Synth Biol 4(5):595–603. https://doi.org/10.1021/sb500260k
Martin VJ, Pitera DJ, Withers ST, Newman JD, Keasling JD (2003) Engineering a mevalonate pathway in Escherichia coli for production of terpenoids. Nat Biotechnol 21(7):796–802. https://doi.org/10.1038/nbt833
Melis A (2013) Carbon partitioning in photosynthesis. Curr Opin Chem Biol 17(3):453–456. https://doi.org/10.1016/j.cbpa.2013.03.010
Niu FX, Lu Q, Bu YF, Liu JZ (2017) Metabolic engineering for the microbial production of isoprenoids: carotenoids and isoprenoid-based biofuels. Synth Syst Biotechnol 2(3):167–175. https://doi.org/10.1016/j.synbio.2017.08.001
Oliver JW, Machado IM, Yoneda H, Atsumi S (2013) Cyanobacterial conversion of carbon dioxide to 2,3-butanediol. Proc Natl Acad Sci USA 110(4):1249–1254. https://doi.org/10.1073/pnas.1213024110
Oliver JW, Machado IM, Yoneda H, Atsumi S (2014) Combinatorial optimization of cyanobacterial 2,3-butanediol production. Metab Eng 22:76–82. https://doi.org/10.1016/j.ymben.2014.01.001
Paddon CJ, Keasling JD (2014) Semi-synthetic artemisinin: a model for the use of synthetic biology in pharmaceutical development. Nat Rev Microbiol 12(5):355
Sengupta A, Pakrasi HB, Wangikar PP (2018) Recent advances in synthetic biology of cyanobacteria. Appl Microbiol Biotechnol. https://doi.org/10.1007/s00253-018-9046-x
Sharkey TD, Wiberley AE, Donohue AR (2008) Isoprene emission from plants: why and how. Ann Bot 101(1):5–18. https://doi.org/10.1093/aob/mcm240
Siwko ME, Marrink SJ, de Vries AH, Kozubek A, Uiterkamp AJS, Mark AE (2007) Does isoprene protect plant membranes from thermal shock? A molecular dynamics study. Biochim Biophys Acta (BBA) Biomembr 1768(2):198–206
Song K, Tan X, Liang Y, Lu X (2016) The potential of Synechococcus elongatus UTEX 2973 for sugar feedstock production. Appl Microbiol Biotechnol 100(18):7865–7875. https://doi.org/10.1007/s00253-016-7510-z
Toth TN, Chukhutsina V, Domonkos I, Knoppova J, Komenda J, Kis M, Lenart Z, Garab G, Kovacs L, Gombos Z, van Amerongen H (2015) Carotenoids are essential for the assembly of cyanobacterial photosynthetic complexes. Biochem Biophys Acta 1847(10):1153–1165. https://doi.org/10.1016/j.bbabio.2015.05.020
Tracy NI, Chen D, Crunkleton DW, Price GL (2009) Hydrogenated monoterpenes as diesel fuel additives. Fuel 88(11):2238–2240
Ungerer J, Pakrasi HB (2016) Cpf1 is a versatile tool for CRISPR genome editing across diverse species of cyanobacteria. Sci Rep 6:39681
Ungerer J, Tao L, Davis M, Ghirardi M, Maness P-C, Yu J (2012) Sustained photosynthetic conversion of CO2 to ethylene in recombinant cyanobacterium Synechocystis 6803. Energy Environ Sci 5(10):8998–9006
Ungerer J, Lin PC, Chen HY, Pakrasi HB (2018) Adjustments to photosystem stoichiometry and electron transfer proteins are key to the remarkably fast growth of the cyanobacterium Synechococcus elongatus UTEX 2973. mBio 9(1):e02327-17. https://doi.org/10.1128/mbio.02327-17
van der Woude AD, Angermayr SA, Puthan Veetil V, Osnato A, Hellingwerf KJ (2014) Carbon sink removal: increased photosynthetic production of lactic acid by Synechocystis sp. PCC6803 in a glycogen storage mutant. J Biotechnol 184:100–102. https://doi.org/10.1016/j.jbiotec.2014.04.029
Wang X, Liu W, Xin C, Zheng Y, Cheng Y, Sun S, Li R, Zhu XG, Dai SY, Rentzepis PM, Yuan JS (2016) Enhanced limonene production in cyanobacteria reveals photosynthesis limitations. Proc Natl Acad Sci USA 113(50):14225–14230. https://doi.org/10.1073/pnas.1613340113
Wang B, Eckert C, Maness PC, Yu J (2018) A genetic toolbox for modulating the expression of heterologous genes in the cyanobacterium Synechocystis sp. PCC 6803. ACS Synth Biol 7(1):276–286. https://doi.org/10.1021/acssynbio.7b00297
Wendt KE, Ungerer J, Cobb RE, Zhao H, Pakrasi HB (2016) CRISPR/Cas9 mediated targeted mutagenesis of the fast growing cyanobacterium Synechococcus elongatus UTEX 2973. Microb Cell Fact 15(1):115
Yoo SH, Keppel C, Spalding M, Jane JL (2007) Effects of growth condition on the structure of glycogen produced in cyanobacterium Synechocystis sp. PCC6803. Int J Biol Macromol 40(5):498–504. https://doi.org/10.1016/j.ijbiomac.2006.11.009
Young JD, Shastri AA, Stephanopoulos G, Morgan JA (2011) Mapping photoautotrophic metabolism with isotopically nonstationary 13C flux analysis. Metab Eng 13(6):656–665. https://doi.org/10.1016/j.ymben.2011.08.002
Yu J, Liberton M, Cliften PF, Head RD, Jacobs JM, Smith RD, Koppenaal DW, Brand JJ, Pakrasi HB (2015) Synechococcus elongatus UTEX 2973, a fast growing cyanobacterial chassis for biosynthesis using light and CO(2). Sci Rep 5:8132. https://doi.org/10.1038/srep08132
Zhou C, Li Z, Wiberley-Bradford AE, Weise SE, Sharkey TD (2013) Isopentenyl diphosphate and dimethylallyl diphosphate/isopentenyl diphosphate ratio measured with recombinant isopentenyl diphosphate isomerase and isoprene synthase. Anal Biochem 440(2):130–136. https://doi.org/10.1016/j.ab.2013.05.028
Zhou J, Wang C, Yang L, Choi E-S, Kim S-W (2015) Geranyl diphosphate synthase: an important regulation point in balancing a recombinant monoterpene pathway in Escherichia coli. Enzyme Microb Technol 68:50–55
Acknowledgements
Funding to support this work was provided by the Office of Science (BER), U. S. Department of Energy, to HBP. PCL was supported by a fellowship from the McDonnell International Scholars Academy at Washington University.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interest
The authors declare that they have no conflict of interest.
Rights and permissions
About this article

Cite this article
Lin, PC., Pakrasi, H.B. Engineering cyanobacteria for production of terpenoids. Planta 249, 145–154 (2019). https://doi.org/10.1007/s00425-018-3047-y
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00425-018-3047-y