
Abstract
Preharvest sprouting (PHS) and late maturity α-amylase (LMA) are the two major causes of unacceptably high levels of α-amylase in ripe wheat grain. High α-amylase activity in harvested grain results in substantially lower prices for wheat growers and at least in the case of PHS, is associated with adverse effects on the quality of a range of end-products and loss of viability during storage. The high levels of α-amylase are reflected in low falling number, the internationally accepted measure for grain receival and trade. Given the significant losses that can occur, elimination of these defects remains a major focus for wheat breeding programs in many parts of the world. In addition, the genetic, biochemical and molecular mechanisms involved in the control of PHS and LMA as well as the interactions with environmental factors have attracted a sustained research interest. PHS and LMA are independent, genetically controlled traits that are strongly influenced by the environment, where the effects of particular environmental factors vary substantially depending on the stage of grain development and ripening. This review is a summary and an assessment of results of recent research on these important grain quality defects.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Much of the early research on preharvest sprouting (PHS) in wheat has been captured in a publication edited by (Derera 1989a). In addition, there have been regular international symposia on preharvest sprouting in cereals beginning in 1975 (see Supplementary 1 for a list of symposia proceedings). More recent research has tended to concentrate on grain dormancy, the major genetic mechanism involved in providing resistance to preharvest sprouting, informed to some extent by the extensive research on grain dormancy in barley and rice as well as in the model plant species, Arabidopsis thaliana. By comparison, late maturity α-amylase (LMA) is a relatively new area of research and involves the untimely synthesis of α-amylase during the middle stages of grain development. Most of this research has been carried out in Australia and the UK and wheat breeders in some parts of the world are only just coming to the realization that it represents a source of low falling number in their germplasm. LMA was reviewed by (Mares and Mrva 2008b).
This review will be restricted to wheat and focuses mainly on developments in the time since these reviews. The authors express their apologies to those whose work may have been unintentionally overlooked or, given that this is not intended to be a compilation of literature, could not be cited.
Definitions
Preharvest sprouting (PHS) refers to the germination of grain in the ear prior to harvest generally in response to rain.
Late maturity α-amylase (LMA) refers to the synthesis of high pI α-amylase during the middle stages of grain development in the absence of sprouting or rain. The enzyme is retained in the grain through to harvest-ripeness.
Preharvest sprouting
Preharvest sprouting (Fig. 1a) in wheat under field conditions is triggered by rain, but the amount of sprouting that occurs in response to a particular rain event or series of events is not easy to predict.

a Sprouted and non-sprouted spikes of wheat. b black point affected, non-sprouted grains (columns 1–3), sound grains (columns 4–6) and black point affected, sprouted grains (columns 7–9). c LMA-affected grains with sound appearance but low falling Number/high a-amylase. Photographs by Cameron Mares
Apart from rain, which is in most cases an absolute requirement, sprouting is affected by a large number of factors that include the following:
Varietal variation in genetic resistance to preharvest sprouting. This represents the base response to a particular rain event that can be modified by environmental conditions during grain ripening, during the rain event and indeed immediately following the rain event. Basically, grain of varieties with greater resistance needs to stay wet for a longer period of time before sprouting will be initiated. This resistance gradually disappears with after-ripening but can dissipate very quickly if moist grain is subjected to cold temperatures (Mares 1984).
Stage of maturity. Sprouting susceptible varieties acquire the capacity to sprout sometime after the middle stages of grain development whereas in resistant varieties it may not occur until well after physiological maturity and harvest-ripeness. As ripening and/or after-ripening proceeds, the time required for grains to germinate or sprout decreases until eventually a significant proportion of grains will germinate in 1–2 days if wet. As a consequence, the amount of sprouting triggered by a rain event is very dependent on the stage of maturity. Note here that primary tillers ripen earlier than secondary tillers such that by the time all the crop is ready for harvest, some primary tillers may have been ripe for several days. Similarly some areas of the crop may ripen earlier than others due to variation in the microenvironment and will be at greater risk.
Amount of rain, duration of the rain event, and weather conditions after the rain (i.e. do they favor the grain remaining wet or do they favor drying). As a rough guide, it would appear that around 10–15 mm of rain is probably the minimum required to initiate sprouting but obviously if it is spread over several hours more water is likely to be captured by the wheat ear than if it all falls in a short burst. Overall, for sprouting susceptible crops at harvest-ripeness, the grain will need to stay moist for around 2 days although this time will decrease further the longer harvest is delayed.
Temperature and moisture stress. The effects of temperature on sprouting are quite complex and can be quite different depending on the moisture content of the grain. However, provided the crop is not under moisture stress then warmer temperatures during ripening tend to result in greater susceptibility to sprouting at harvest-ripeness. If the crop is under moisture stress combined with high temperatures then resistance to sprouting may be significantly increased compared to expectation (Biddulph et al. 2007). One possible explanation for this is that environmental-associated variation on grain drying rate may not be accompanied by a similar variation in rate of grain after-ripening. The effects of temperature on germinability of wet grain are also quite complex; cool temperatures may reduce or break dormancy whilst higher temperatures can accentuate any grain dormancy that is present (Mares 1984).
Rainfall prior to harvest-ripeness. Rain (more than 15–20 mm) during the 10–20 days prior to harvest-ripeness often results in little or no sprouting but may predispose the crop to be more susceptible to later rainfall (Mares 1993). In a study conducted in northern NSW, Australia, there was a strong positive correlation between the total rainfall in the 10–20 days prior to harvest-ripeness and the amount of sprouting that occurred when samples were subsequently subjected to a standard wetting treatment at harvest-ripeness or at 10 or 20 days after harvest-ripeness.
Crop phenology, crop architecture, and physical/chemical properties of the wheat ear. These can all play a role in determining how much water is retained by the ear and how long the ear remains wet. Some of these factors were considered by King (1989) but there has been little subsequent research. Possible inhibitory effects of the glumes on the germination of the enclosed grains have been proposed but not confirmed. There is a significant lag phase between wetting of intact spikes and both water uptake and germination of the enclosed grain relative to isolated grains, and indeed some variation in the length of this lag phase (Mares 1983).
Black point. Grains exhibiting black point symptoms (Fig. 1b) such as described by Williamson (1997) and (Fernandez et al. 2011) tend to germinate more readily than grains from the same sample that are free of symptoms.
Stages in preharvest sprouting
Sprouting encompasses a sequential series of events from the interception of rain by the vegetative structures of the wheat ear, transfer of water to the enclosed grain, germination and subsequent production of a range of hydrolytic enzymes. Potentially therefore there are a number of steps that could be affected by genetic or environmental factors to confer resistance. The extent of variation in the rate of these events was compared in a small set of genotypes with the results suggesting that genetic variation was greatest for grain dormancy, i.e. failure of grains to germinate when placed in a warm, moist environment normally considered conducive to germination, and therefore the most promising target for variety improvement (Mares 1987). That being said, a resistance mechanism, whether it be germination inhibitors or simply variation in the rate of transfer of water from the glumes of the wheat ear to the enclosed grain, associated with the vegetative structures of the wheat ear would be very advantageous since it would have no carry-over effects on the germination of seed to establish the next crop generation. This would greatly simplify the retention of sprouting resistance in breeding programs that use rapid population development techniques or in long season wheat varieties where the turnaround between harvest and seeding is very short. In both of these situations, there is significant selection pressure for low or no dormancy unless active measures are taken to break dormancy prior to seeding. Once grains achieve the level of moisture required to initiate germination, and if the grains remain wet, the production of hydrolytic enzymes can proceed, and in the case of α-amylase, this follows an exponential rate curve. This is important to understand since only a relatively small increase in total α-amylase of a bulk sample is required to reduce falling number below receival standards and create issues for end-product processing and quality. End-products prepared from sprouted grain are invariably poorer in quality compared with sound grain (Edwards et al. 1989). As a general rule, it is not common for all, or even a high proportion of, grains to sprout simultaneously, rather a small percentage of grains germinate to varying degrees depending on the conditions. However, because of the propensity of grains to produce very high levels of α-amylase, a small proportion of sprouted grains may be sufficient to drop the falling number below receival standards (Canadian Grain Commission fact sheet). Visual inspection provides only a rough indication of the extent of damage and is unreliable for classification of incoming wheat deliveries. Once sprouting is detected, the use of an objective test such as falling number is far more reliable method of determining the extent of sprouting damage and its likely impact on processing.
Selection for preharvest sprouting resistance
Since preharvest sprouting results in significant losses in many wheat growing areas of the world, a high priority target for many wheat breeders is resistance to sprouting. Early work in this area tended to rely on opportunistic screening in field trials in response to rain. In view of the complex interactions between genotype, environment, and stage of maturity this is not seen as a viable method even where rainfall patterns are very reliable; there are simply too many variables. The most appropriate screening method for selecting resistance to preharvest sprouting is effectively a choice between sprouting induced by artificial wetting in a controlled environment with scoring achieved by visual observation, percent germination or falling number; or grain dormancy assessed by a standard germination test on isolated grains. In both cases, it is important to remove maturity effects as far as possible by testing material, sampled either from specifically designed field or glasshouse experiments, at a defined stage of ripening. For reasons that are not entirely clear, the heritability of resistance scored after wetting of ears can often be very low compared with heritability for grain dormancy (Trethowan 1995). By contrast, DePauw and McCaig (1991) reported that rain simulation treatments gave high heritability estimates. Muncvold et al. (2009) and Chao et al. (2010) both compared ear wetting treatments and grain germination tests in genetic studies of resistance to preharvest sprouting and reported a number of genetic loci associated with variation in wetting induced sprouting that did not correspond with loci associated with dormancy. However, it was evident in these studies that the major loci affecting both PHS and dormancy tended to be consistent across environments in contrast to loci associated only with ear sprouting but not grain dormancy. Mares (1993) observed a significant effect of rain prior to maturity on the level of resistance retained at harvest-ripeness and proposed the use of rain shelters to exclude rain during the final 2 weeks of ripening (Mares 1989). Further modification of screening trial management was reported by Mares et al. (2005) with the aim of maximizing genetic effects and reducing effects of environmental factors.
Grain dormancy
For a short period after flowering as the embryo develops, grains or detached embryos are incapable of germination. At a later stage, around the middle of the period between flowering and maturity, detached embryos, and indeed proximal sections of grains, will germinate readily when placed on agar or moist filter paper at 20 °C. Germination characteristics and rates for detached embryos or embryo-containing grain sections are similar for most wheat genotypes irrespective of the germinability of intact grains sampled at the same point in development. Observations of this nature are the basis for the generally held view that dormancy in wheat is primarily coat imposed although the mechanism(s) involved is far from clear. This hypothesis is supported by genetic (Noll et al. 1982; Mares 1999; Flintham et al. 1999) and mutation studies (Warner et al. 2000; Mares 1999) as well as observations that damage to the seed coat reduces grain dormancy (Mares 1989). Results of reciprocal crosses involving dormant and non-dormant parents (Noll et al. 1982; Mares 1993) also provide strong evidence that the seed coat of dormant genotypes is an important factor in the expression of the dormant phenotype. Seed coat-imposed dormancy does not necessarily mean that all the genes involved are expressed in the seed coat, rather it is likely that there are a number of genes expressed in the embryo that determine whether the embryo is sensitive to the effects of the seed coat. It is likely that oxygen availability, limited by the low solubility of oxygen in water and the reported diffusion coefficient of oxygen through seed coats of various seeds (Edwards 1973; Lenoir et al. 1986), could restrict germination. Whether there is genetic variation for sensitivity to oxygen concentration, whether the effects simply enhance existing dormancy mechanisms as appears to be the case in barley (Benech-Arnold et al. 2006) and whether changes during after-ripening affect oxygen availability remain unclear. For isolated grains, water uptake by the embryo appears to be independent of variation in dormancy and restricted to the micropyle (Rathjen et al. 2009). In a wheat spike, the micropyle is located within the zone of attachment of the grain to the rachilla and it is not known whether this impedes moisture flow into the grain.
Approaching physiological maturity, intact grains of some genotypes become capable of germination or in other words are released from dormancy. Grains of these genotypes will germinate or sprout quite readily when harvest-ripe and are consequently characterized as non-dormant and the variety ranked as sprouting susceptible. It is not clear whether such genotypes simply acquire the capacity to germinate as soon as the embryo is mature or whether dormancy is imposed but then released after a very short period. Some observations appear to be consistent with the latter proposal; firstly, under some but certainly not all environmental conditions, wheat grains may show a transient period of germinability at around the time detached embryos acquire the capacity to germinate; and secondly, when detached embryos or embryo-containing grain sections are incubated with abscisic acid (ABA), the germination characteristics typical of the intact grain can be re-imposed. Sensitivity of germination to inhibition by ABA has been reported to change in parallel with dormancy loss during after-ripening (Walker-Simmons 1987). For genotypes that are characterized as being dormant or sprouting resistant, dormancy release is either delayed until after harvest-ripeness or in some cases requires a prolonged period of after-ripening. At least in the latter instance, the changes that trigger dormancy release take place in grain that may have less than 10 % moisture content. These changes have not been characterized and most research has concentrated on changes that occur when the grains are imbibed to initiate germination. In using terms such as dormancy and dormancy release, it is important to remember that these are not absolute traits but rather they are changing continuously with time, are dependent on the environmental conditions during grain development and also on the temperature used in the germination test. Indeed, even grains of reputed non-dormant cultivars continue to change during after-ripening with germination rate increasing and time to 50 % germination decreasing for some weeks. Taking a snapshot at a particular stage in development/after-ripening, e.g. at physiological maturity, at a particular moisture content, at a set time after flowering, or a point when the grain is ready to harvest provides useful information that is used in genetic and molecular studies. Whilst not practical for large numbers of genotypes, a more complete picture can be obtained by sampling for germination testing at intervals during grain development and during after-ripening (Gerjets et al. 2009). Gerjets et al. (2009) used a small number of wheat varieties that varied in dormancy. In reports presented at the 12th International Symposium on Preharvest Sprouting in Cereals in 2012, Mares et al. and Torada et al. used the same technique to compare changes in closely related genotypes that not only differed in dormancy but also for closely related genotypes that contained different individual dormancy quantitative trait loci (QTL) or combinations of QTL. This provided strong evidence for the additive effects of QTL on the time to dormancy release and on their relative effects on dormancy measured at a particular stage of ripeness. This information will be useful to breeders since it allows QTL to be ranked according to the magnitude of the effect on dormancy and its relationship to the relative risk of sprouting in the breeder’s target environment. The grain development/germination profile (Fig. 2) can be characterized by a number of parameters that vary in their ease of estimation. These include: time to dormancy release, time to 50 % germination in a test conducted for a fixed number of days, the slope of the germination response following the release of dormancy, or the germination index at a specific point in ripening, often the point corresponding to harvest-ripeness in a particular country. It should be noted that the final phase of grain ripening involves the loss of moisture, whose rate is very much dependent on the environmental conditions (temperature, relative humidity, wind) and to some extent independent of the rate of processes leading to grain harvest-ripeness.

Alternate methods for describing changes in dormancy/germination during grain ripening for genotypes (1–5) with increasing numbers of PHS QTL and increasing dormancy. Germination index at harvest-ripeness (12 % moisture, 55 Dpa) (black dashed line), time to achieve a GI of 0.5 (Dpa) (blue dashed line), slope of the GI × Dpa curve for genotype 2 following release from dormancy (red dashed line), with the time of release from dormancy estimated by the intercept on the Dpa axis
Grain of bread wheat, T. aestivum, can be red or white in color. Red color is controlled by dominant wild-type alleles, R-A1b, R-B1b, and R-D1b, at independent R gene loci located on the long arms of the homoeologous group 3 chromosomes. The presence of any one, two or all three dominant alleles confers red grain coat color with the darkness of the color generally increasing with the number of wild-type alleles. In contrast, white grain coat color is recessive and requires naturally occurring mutant alleles, R-A1a, R-B1a, and R-D1a, to be present at all three R loci. Historically, sprouting resistance and grain dormancy has been linked to red grain coat color, however, it is now clear that there is substantial variation for resistance in both classes. Despite the independent development of sprouting tolerant white-grained wheat varieties in several countries over the past 30 years (Derera 1989b; Anderson et al. 1993; Mares et al. 2005; Muncvold et al. 2009; Liu and Bai 2010), it is still common unfortunately to find sweeping statements in recent literature to the effect that white-grained wheat varieties are susceptible and red-grained wheat varieties resistant to preharvest sprouting. Whilst there are numerous examples of red wheat varieties that are susceptible to sprouting it is quite evident that the most resistant red types are more resistant than the best white-grained genotypes. Mares (1999) reported the isolation of dormant and non-dormant red-grained progeny from crosses between a single gene red, dormant variety and a white, non-dormant variety clearly demonstrating the red color does not guarantee resistance. Later work indicated that the variation within the red-grained sub-population was associated with a QTL on chromosome 4A and that this QTL conferred intermediate dormancy in some of the white-grained progeny (Mares et al. 2005). Similarly, Flintham (2000) compared the resistance in progeny from a cross between two 3-gene red varieties, one resistant, and one susceptible and observed a 1:1 (resistant: susceptible) segregation ratio. The difference between the two groups was reported to be a major gene referred to as PHS that was not associated with red grain coat color. This gene was subsequently shown to be located on chromosome 4A (Flintham et al. 2002) consistent with earlier reports of the association between a QTL on 4A and grain dormancy (Anderson et al. 1993; Mares and Mrva 2001; Kato et al. 2001). These observations appear to suggest that dormancy in red-grained wheat varieties involves an interaction between the R-gene(s) and other genes that are not associated with grain color but which on their own confer some level of dormancy. These other genes can therefore be utilized to develop sprouting resistant white-grained wheat genotypes. Populations derived from crosses between red, dormant, and white, non-dormant parents contain some white-grained progeny with dormancy intermediate to, but never equivalent to, the red dormant parent. The intermediate dormancy of a white-grained mutant recovered from mutagenesis of dormant red wheat, AUS1490, (R-A1b, R-B1a, R-D1a) is consistent with this hypothesis (Mares 1999; Mares et al. 2005). Himi et al. (2011) reported that the R genes are in fact myb transcription factors, Tamyb10, and demonstrated that the AUS1490 mutant contained a point mutation in the R-A1 gene, Tamyb10-A1. Earlier, Warner et al. (2000) reported that two white grained mutants derived from Chinese Spring, R-D1b, had reduced dormancy compared with the parent genotype. To this point, it has not been unequivocally demonstrated whether red grain coat color per se has an effect on dormancy phenotype. Flintham (2000) compared sprouting resistance in sets of near-isogenic lines representing individual R genes in several white-grained backgrounds and concluded that the different dominant R alleles had similar effects on resistance. In contrast, Fofana et al. (2009) mapped germination index, sprouting index and falling number in AC Domain × White RL4137, both derived from the 3-gene red wheat RL4137, and reported that the proportion of variation associated with QTL linked to the individual R genes were quite different and ranked in order 3B > 3D > 3A. The R genes appear to be additive in terms of their effect on grain coat color and there is anecdotal evidence to suggest that the effects of the individual R genes on sprouting resistance are also additive.
Quantitative trait loci (QTL) associated with variation in grain dormancy
Clearly, given the complexity of preharvest sprouting and the difficulties associated with phenotypic selection of resistant genotypes, marker-assisted selection is an extremely attractive option for breeders. This aspiration must be tempered by the substantial number of QTL reported to be associated with sprouting resistance (Flintham et al. 2002, Supplementary 2) although many have not been validated in subsequent studies. In the case of red-grained wheat, breeders have the advantage of simple visual selection for presence of R genes, which are additive in effect with QTL not associated with grain color. Depending on the level of resistance required in their target environment it may well be that red coat color in combination with one of the major dormancy QTL provides sufficient protection, particularly given the advances in machine technology that allow rapid harvest of large areas. By comparison, there are no visual markers for dormancy in white-grained wheat genotypes and several QTL may have to be assembled to provide adequate resistance in higher risk environments. Not a simple task in view of the numbers of individuals that would need to be tested, the difficulties in phenotyping, and the need to retain a host of other traits required in a commercial wheat variety. Major QTL that have been validated by several researchers in different germplasm are located on chromosomes 4A (Kato et al. 2001; Mares and Mrva 2001; Mares et al. 2005; Flintham et al. 2002), 3AS (Osa et al. 2003; Mori et al. 2005) and 2B (Muncvold et al. 2009; Chao et al. 2010; Somyong et al. 2011). Tightly linked or diagnostic markers have been developed for these QTL however, a substantial proportion of observed genetic variation remains unexplained (Mares et al. 2009) and a combination of phenotyping and genotyping is required to recover the full dormancy at least from some sources. Dormancy QTL located on chromosome 4A has been fine mapped but as yet a candidate gene has not been identified. By contrast, the QTL on the distal end of 3AS is reported to be co-localized with wheat homologue of Mother of FT and TFL1 (MFT) which appeared to be upregulated in dormant seeds grown at lower temperature (Nakamura et al. 2011; Liu et al. 2013a). The QTL on 2B has been fine mapped and a potential candidate gene, a putative ABA receptor, identified (Somyong et al. 2011). Zhang et al. (2014) cloned a wheat ortholog of the rice dormancy gene, OsSdr4, and whilst it was reported to reside in a similar chromosome interval as the 2B QTL described by (Muncvold et al. 2009) it explained only a small proportion of the observed variation in dormancy phenotype. A single nucleotide polymorphism in the promotor region of TaSdr-B1 gave rise to two alleles that were associated with differences in germination index. Other QTL of potential interest are the wheat homologues of Vp-1, a mutant of which was shown to be associated with vivipary in maize (McCarty et al.1989). McKibbin et al. (2002) reported that the majority of transcripts of Vp-1 homologues in modern wheat and ancestral species are mis-spliced, possibly contributing to susceptibility to preharvest sprouting. Whilst the Vp-1 locus does not appear to have figured strongly in PHS and dormancy QTL mapping studies, Yang et al. (2007, 2014) and Chang et al. (2011) have presented evidence that allelic variants of Vp-1B and Vp-1A are associated with resistance to sprouting. These studies were conducted using collections of cultivars and should be validated in specifically constructed populations. Similarly, considerable research in Arabidopsis seed dormancy has focused on DOG1 (delay of germination) genes and insertion of a DOG-1-like wheat homologue into Arabidopsis was reported to increase dormancy, prompting Ashikawa et al. (2010) to suggest that overexpression of these genes might reduce sprouting in wheat.
Role of plant hormones
Mechanisms involved in grain dormancy in wheat remain elusive (Jacobsen et al. 2013). Abscisic acid and gibberellin have long been implicated in development of, and release from, dormancy in a wide range of plant species. Whilst abscisic acid appears to be critical for development of grain dormancy in wheat it seems likely that it is the sensitivity of the embryo to this hormone rather than the actual concentration that is critical. Walker-Simmons (1987) reported that embryo sensitivity to ABA was retained longer in dormant wheat varieties and a number of subsequent studies have shown that changes in embryo sensitivity parallel changes in dormancy as measured by germination of the intact grain. A common method of breaking dormancy in wheat involves imbibition at cool, usually 4 °C, for a few days. Loss of dormancy in this system was rapid and associated with an equally rapid loss in embryo sensitivity to ABA (Noda et al. 1993). Noda et al. (2002) reported that ABA sensitivity was conditioned by the QTL located on chromosome 4A. More recent work has failed to confirm this hypothesis and rather suggests that embryo sensitivity to ABA is a characteristic of grain that shows reduced germination rather than a property related to a particular QTL (Mares unpublished data). In contrast to the establishment of dormancy, the role of hormones in the release from dormancy during dry after-ripening remains unclear. (Liu et al. 2013b) investigated hormone synthesis, catabolism and signaling in dormant and after-ripened grain, finding little or no significant change except following imbibition. Unfortunately these authors only examined whole grains and changes in the embryo may have been masked by the much larger endosperm and seed coat tissues. In contrast, Barrero et al. (2009) concentrated on the embryo of barley and reported evidence of changes in ABA metabolism and sensitivity related to after-ripening. Wilson et al. (2005) concluded that most of the transcripts required for germination in wheat grains accumulated in the embryo prior to imbibition and germination. A role for gibberellin has not been demonstrated and its function rather appears to facilitate germination once dormancy has been released (Bewley 1997). In addition to ABA, Jacobsen et al. (2013) have reported that blue light, jasmonate, and nitric oxide play important roles in regulation of dormancy in imbibed wheat grain.
Late maturity alpha-amylase (LMA)
LMA refers to a genetic defect that appears to be widely distributed in bread, T. aestivum L., and durum, T. durum, wheat as well as in synthetic hexaploid wheat (Mares and Mrva 2008a; Mrva et al. 2009). Depending on the genotype and the environment, presence of the defect can result in a short period of new synthesis of high pI α-amylase by the grain aleurone beginning around 20–30 days after anthesis. The enzyme is retained through to harvest causing a reduction in falling number and potentially a failure to meet receival or market specifications. LMA-affected grain retains a bright, sound appearance (Fig. 1c) and both incidence and magnitude are very difficult to predict.
Expression of LMA
Tall LMA genotypes (wild-type height genes, Rht-B1a and Rht-D1a) and semi-dwarf LMA genotypes where height reduction is not due to gibberellin insensitivity (for example Rht8 as opposed to GA-insensitivity mutant alleles Rht1 (Rht-B1b) or Rht2 (Rht-D1b)) may show a constitutive pattern of expression in a wide range of environmental conditions (Mrva et al. 2008). By comparison, semi-dwarf LMA genotypes, where height reduction is conditioned by the mutant alleles Rht1 or Rht2, only show constitutive expression when grown under a very narrow range of conditions (Mrva and Mares unpublished data). Whilst these conditions are still to be fully characterized, constitutive expression in these semi-dwarfs has been observed in controlled environments where temperature ranged from 15 to 25 °C (night/day), in glasshouse experiments where maximum daily temperature was maintained below 25 °C, and in some field experiments that ripened under mild conditions in the absence of moisture stress. LMA can be consistently triggered by subjecting these semi-dwarf LMA genotypes to a cool temperature shock (transfer from warm day temperatures >25 °C to cool, 18/12 °C, day/night environment) of several days duration applied during a window of sensitivity that occurs in mid-grain development (Mrva and Mares 2001). The difference in pattern of LMA expression between tall and semi-dwarf genotypes appears to be solely attributable to the effects of the GA-insensitivity alleles, Rht1 and Rht2, and there is no evidence that there is a fundamental difference in LMA per se. Transfer of LMA from tall to semi-dwarf backgrounds, or vice versa, results in a shift in expression pattern to that typical of the recipient height type. An exception to this pattern appears to be Rht1 or Rht2 LMA semi-dwarfs that also contain the 1B/1R wheat rye translocation. Some of these genotypes may exhibit constitutive expression (Mrva et al. 2008) suggesting that the presence of the translocation may under some conditions overcome the effects of the GA-insensitive semi-dwarfing genes. In many field situations, semi-dwarf LMA genotypes do not exhibit an LMA phenotype either because the maximum daily temperature during ripening exceeds 25 °C, there is no cool temperature shock or the shock does not coincide with the window of sensitivity, or there is some level of moisture stress which acts to repress LMA expression. Semi-dwarf wheat varieties now dominate global wheat production and whilst we know from targeted phenotyping that LMA is widespread, the strong effects of environment and maturity make it extremely difficult to predict the occurrence and severity of LMA expression in the field.
Quantitative assessment of LMA phenotype
Determination of the LMA status of a particular wheat genotype currently depends on facilitating effective LMA expression, either constitutive or cool temperature shock induced, followed by the measurement of α-amylase protein or activity.
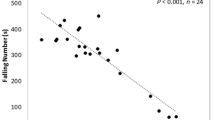
During grain development the low pI α-amylase, controlled by Amy-2 genes located on the group 7 chromosomes, which is synthesized in the pericarp shortly after anthesis declines to residual levels of activity by physiological maturity. This decline may be variable between genotypes and may be affected by environmental conditions, notably frost which may disrupt the normal degradation. The activity of low pI α-amylase can therefore confound the determination of LMA if assays for total α-amylase activity are used. To overcome this problem, a high pI-specific ELISA based on the method described by Verrity et al. (1999) has been developed (Mares and Mrva 2008b; Barrero et al. 2013). Unfortunately the antibodies required for the ELISA are not available commercially or outside Australia and as a result, effort has been focused on alternative methods. Cheng et al. (2014) cloned the family members of high pI genes expressed in LMA and suggested that it would be possible to develop a PCR-based assay. The obvious limitation of this proposal is the apparent short life of the high pI α-amylase mRNA (Barrero et al. 2013).
Physiology and biochemistry of LMA
Wheat grains normally do not synthesize high pI α-amylase until after maturity when they may sprout in response to rain or germinate following sowing the next season’s crop. In LMA-prone wheat genotypes, synthesis may be triggered before the grain has reached physiological maturity and whilst the grain moisture content is still between 50 and 60 % fresh weight. It involves the coordinated transcription of the Amy-1 genes located on the long arms of the group 6 chromosomes and takes place in isolated cells or cell islands distributed throughout the grain aleurone system (Mrva et al. 2006). There is no evidence for involvement of the embryo in contrast to the situation in germinating grains where synthesis is initiated in the scutellum, and later the aleurone, and gradually extends towards the distal end of the grain. Appearance of high pI α-amylase is preceded by a transient period of mRNA synthesis and whilst the mRNA appears to be short-lived, the enzyme protein is quite stable and is retained through to maturity (Mares and Mrva 2008b; Barrero et al. 2013). The mechanism(s) involved in the inhibition of further mRNA synthesis have not been identified. Unlike germination where α-amylase synthesis can follow an exponential rate curve, in LMA, synthesis reaches a plateau at a relatively low level of activity which is nevertheless sufficient to reduce falling number below the acceptable limits for receival into milling grades. The different patterns of enzyme synthesis mean that LMA-affected and sprouted grains can be distinguished by cutting grains in half and comparing α-amylase activities in each half. Also unlike germination, there is no evidence of concomitant synthesis of low pI α-amylase, endo-protease or other hydrolytic enzymes by the aleurone. Either LMA is limited to specifically high pI α-amylase, or the period of aleurone activation is too short for synthesis of enzymes that normally appear sometime after α-amylase in germinating grains. The period in grain development immediately preceding and during high pI α-amylase synthesis is characterized by quite dramatic changes in gene expression and hormone composition (Barrero et al. 2013). Microarray experiments identified a limited numbers of genes whose expression was either up-regulated or down-regulated in LMA versus non-LMA genotypes. Barrero et al. (2013) reported dramatically elevated levels of gibberellins such as GA19 and much lower levels of auxin in the de-embryonated fraction of grains sampled shortly after the initiation of α-amylase synthesis. Follow-up studies by the authors of this review have demonstrated that the increase in GA levels precedes the synthesis of both mRNA and α-amylase protein (Mrva and Mares unpublished data). Interestingly, the predominant GA species identified, GA19, is reported to be an inactive precursor of biologically active GA1 which was not detected making it difficult to connect the hormone profile with the stimulation of α-amylase synthesis. By contrast, Kondhare et al. (2014) reported both GA1 and GA3 in extracts from whole grains of LMA genotypes, however, it is possible their results may have been confounded by the use of whole grain rather than the specific grain fraction, aleurone, involved in LMA expression. Nevertheless, several lines of evidence are consistent with the involvement of GA in LMA expression: the well-established signal transduction pathway in grain aleurone involving GA, DELLA (Rht in wheat) and a GAmyb that triggers α-amylase synthesis; the effect of the GA-insensitive mutants, Rht1, Rht2 and Rht3, in reducing the expression of LMA; and stimulation of α-amylase synthesis in LMA-prone cultivars following application of exogenous GA during grain development (Kondhare et al. 2012). The latter report is open to some question given that only total α-amylase was measured and it is well established that application of exogenous GA may stimulate expression of both low and high pI α-amylase. It is possible that GA19 is only converted to biologically active GA in the small fraction of aleurone cells that were reported by Mrva et al. (2006) as undergoing programmed cell death similar to germination or GA-challenged aleurone. If this is the case, the amounts of active GA might well be below the level of detection due to dilution by the bulk of aleurone cells that do not appear to express LMA.
Alternatively, the failure to detect GA1 may be a consequence of efficient 1β-hydroxylation of GAs that occurs in developing grains of wheat. 1β-hydroxylated derivatives of GA20 and GA1 (GA60 and GA55 respectively) have been detected in developing wheat grains, as well as 1β-hydroxylated derivatives of GA9 and GA4 (GA61 and GA54 respectively) (Gaskin et al. 1980). Very efficient 1β-hydroxylation is suggested by the estimate of a 5000-fold excess of GA54 compared to GA1 (Lenton and Gale 1987). Although GA54 was much less (30- to 100-fold) active than GA1 in the induction of α-amylase, it would still contribute the majority of bioactive GA activity (Lenton and Gale 1987). Unfortunately, the lack of deuterated standards for the 1β-hydroxylated GAs means that quantitation by standard GC–MS procedures is lacking. Mares and Mrva (2008b) suggested that the effect of a cool shock in stimulating expression of LMA in LMA-prone semi-dwarfs might be due to a reduction in the GA insensitivity based on earlier reports summarized in Mares and Mrva (2008b), a report by Tomkinson et al. (1997) and the absence of the cool temperature shock requirement in semi-dwarfs where height reduction was not dependent on GA insensitivity (Mrva et al. 2008). This suggestion appears to be supported by recent work reported by Kondhare et al. (2013).
Genetic control
LMA is inherited as a recessive trait (Mrva and Mares 1999). Early research using a population involving the LMA-prone Australian variety, Cranbrook, indicated that the trait appeared to be multigenic with a major locus on the long arm of chromosome 7B and a minor locus on 3B (Mrva and Mares 2001). Subsequent research has identified other QTL on 3A, 2D (Mrva and Mares unpublished data) and 6B (Emebri 2010) whilst the major QTL on 7B has been confirmed in over eight sources of LMA used in Australian breeding programs, germplasm from China and Mexico and in synthetic hexaploid wheat (Mrva et al. 2009), as well as in cultivars derived from the UK variety, Huntsman (Mohler et al. 2014). The QTL on 3A and 3B are located close to the centromere and appear to be homoeologous loci. Fine mapping of the LMA QTL on 7B and 3B is already well advanced to the point of identification of a very small number of candidate genes residing in a chromosome interval of a fraction of a cM around the 7B QTL. Interestingly, with the exception of the minor LMA 2D QTL, the LMA QTL are restricted to the A and B genomes. LMA has been identified in a significant number of durum cultivars (AABB) whilst LMA is present at extremely high frequency in synthetic hexaploid wheat (SHW) (Mrva et al. 2009) that are produced by interspecies hybridization of durum wheat and the D-genome donor of bread wheat, Aegilops tauschii. This is very disappointing given that the generation of SHWs was strategy devised to access the previously untapped genetic variation present in these wild diploids. Whilst there has been only limited study of the genetic control of LMA in SHWs there has been no indication of novel QTL on the D genome, rather the LMA QTL appear to derive from the durum parent.
Concluding remarks
PHS
Major sprouting and/or dormancy QTL have been identified that explain part of the genetic variability and the identification of the underlying genes, or candidate genes, for major QTL is an exciting development. Marker-assisted-selection is very effective for these QTL but to capture the entire resistance from some sources it is still necessary to supplement genotyping with phenotyping. The need for phenotyping raises the issue of how to retain resistant or dormant individuals within breeding programs where there is often little opportunity for natural after-ripening. Imbibition at cold temperatures or physical damage to the seed coat are quite effective at eliminating dormancy but are not necessarily very practical in a large scale breeding operation. Finally, some of the major QTL that have been identified are much less effective when grain ripens under warmer temperatures and may therefore become less useful in a warming global environment. Preliminary results indicate that QTL that are clearly present in some sources, but not as yet identified, could be very useful in buffering resistance or dormancy against the effects of fluctuating environmental conditions. Mutation studies have identified both ABA-insensitive (reduced dormancy) and ABA-hypersensitive (increased dormancy) mutants in wheat (Schramm et al. 2013 and references cited) with the latter offering some prospect for improving PHS resistance. Vp-1 genes in wheat are associated with a high degree of mis-splicing and much less effective in reducing premature germination. However, insertion of Vp-1 genes from wild oat, Avena fatua, (McKibbin et al. 2002) or maize, Zea mays, (Huang et al. 2012) into wheat has been reported to increase resistance to preharvest sprouting.
LMA
Recent surveys of breeders germplasm have demonstrated that this genetic defect is widespread and at high frequency. Because the research is relatively new it is not possible to decide whether this is a recent development or whether it has long been the case but simply not detected. The scale of the problem presents a serious impediment to progress in breeding new cultivars, particularly in countries where grain is exported into markets with stringent quality specifications, and a significant economic concern for breeding companies. Fortunately screening procedures are improving rapidly whilst advances on the molecular genetic front provide a real hope that genotyping screening will be an effective strategy in the near future. Whilst LMA is associated with dramatic changes in both hormone content and transcript profiles during grain development there are no obvious effects on grain appearance, development, or morphology. The high degree of retention of LMA in breeding programs may reflect some as yet unknown positive effects on grain production or alternately simply reflect the absence of significant selection pressure against this trait.
Abbreviations
- PHS:
-
Preharvest sprouting
- LMA:
-
Late maturity α-amylase
- QTL:
-
Quantitative trait locus
- ABA:
-
Abscisic acid
- GA:
-
Gibberellic acid
- PCR:
-
Polymerase chain reaction
- SHW:
-
Synthetic hexaploid wheat
References
Anderson JA, Sorrells ME, Tanksley SD (1993) RFLP analysis of genomic regions associated with resistance to preharvest sprouting in wheat. Crop Sci 33:453–459
Ashikawa I, Abe F, Nakamura S (2010) Ectopic expression of wheat and barley DOG1-like genes promotes seed dormancy in Arabidopsis. Plant Sci 179:536–542
Barrero JM, Talbot MJ, White RG, Jacobsen JV, Gubler F (2009) Anatomical and transcriptomic studies of the coleorhiza reveal the importance of this tissue in regulating dormancy in barley. Plant Physiol 150:1006–1021
Barrero JM, Mrva K, Talbot MJ, White JT, Gubler F, Mares DJ (2013) Genetic, hormonal and physiological analysis of late maturity α-amylase (LMA) in wheat. Plant Physiol 161:1265–1277
Benech-Arnold RL, Gualano N, Leymarie J, Come D, Corbineau F (2006) Hypoxia interferes with ABA metabolism and increases ABA sensitivity of dormant barley grains. J Exp Bot 57:1423–1430
Bewley D (1997) Seed germination and dormancy. Plant Cell 9:1055–1066
Biddulph TB, Plummer JA, Setter TL, Mares DJ (2007) Influence of high temperature and terminal moisture stress on dormancy in wheat (Triticum aestivum L.). Field Crops Res 103:139–153
Canadian Grain Commission Fact Sheet “Sprouted and severely sprouted kernels” https://www.grainscanada.gc.ca/fact-fait/sd-gg-eng.htm
Chang C, Zhang H-P, Zhao Q-X, Feng J-M, Si H-Q, Lu J, Ma C-X (2011) Rich allelic variations of Viviparous-1A and their associations with seed dormancy/pre-harvest sprouting of common wheat. Euphytica 179:343–353
Chao S, Xu SS, Elias EM, Faris JD, Sorrells ME (2010) Identification of chromosome locations of genes affecting preharvest sprouting and seed dormancy using chromosome substitution lines in tetraploid wheat (Triticum turgidum L.). Crop Sci 50:1180–1187
Cheng C-R, Oldack K, Mrva K, Mares D (2014) Analysis of high pI α-Amy-1 gene family members expressed in late maturity a-amylase in wheat (Triticum aestivum L.). Mol Breed 33:519–529
DePauw RM, McCaig TN (1991) Components of variation, heritabilities and correlations for indices of sprouting tolerance and seed dormancy in Triticum spp. Euphytica 52:221–229
Derera NF (1989a) Preharvest Sprouting in Cereals. CRC Press Inc., Boca Raton
Derera NF (1989b) Breeding for preharvest sprouting tolerance. In: Derera NF (ed) Preharvest sprouting in cereals. CRC Press Inc., Boca Raton, pp 111–128
Edwards MM (1973) Seed dormancy and environment: internal oxygen relationships. In: Heydecker W (ed) Seed ecology. Butterworths, London, UK, pp 169–188
Edwards RA, Ross AS, Mares DJ, Ellison FW, Tomlinson JD (1989) Enzymes from rain-damaged and laboratory germinated wheat I. Effects on product quality. J Cereal Sci 10:157–167
Emebri LC, Oliver JR, Mrva K, Mares DJ (2010) Association mapping of late maturity a-amylase (LMA) activity in a collection of synthetic hexaploid wheat. Mol Breed 26:39–49
Fernandez MR, Sissons M, Conner RL, Wang H, Clarke JM (2011) Influence of biotic and abiotic factors on dark discoloration of durum wheat kernels. Crop Sci 51:1205–1214
Flintham JE (2000) Different genetic components control coat-imposed and embryo-imposed dormancy in wheat. Seed Sci Res 10:43–50
Flintham J, Adlam R, Gale M (1999) Eighth International Symposium on Pre-Harvest Sprouting in Cereals 1998. Weipert D (ed). Association of Cereal Research. Federal Centre for Cereal, Potato and Lipid Research, Detmold, Germany, pp 67–76
Flintham JE, Adlam R, Bassoi M, Holdsworth M, Gale M (2002) Mapping genes for resistance to sprouting damage in wheat. Euphytica 126:39–45
Fofana B, Humphreys DG, Cloutier S, Brule-Babel A, Woods S, Luckow OM, Somers DJ (2009) Mapping quantitative trait loci controlling pre-harvest sprouting resistance in a red 3 white seeded spring wheat cross. Euphytica 165:509–521
Gaskin P, Kirkwood PS, Lenton JR, MacMIllan J, Radley ME (1980) Identification of gibberellins in developing wheat grain. Agric Biol Chem 44:1589–1593
Gerjets T, Scholefield D, Foulkes MJ, Lenton JR, Holdsworth MJ (2009) An analysis of dormancy, ABA responsiveness, after-ripening and pre-harvest sprouting in hexaploid wheat (Triticum aestivum L.) caryopses. J Exp Bot 61:597–607
Himi E, Maekawa M, Miura H, Noda K (2011) Development of PCR markers for Tamyb10 related to R-1, red grain color gene in wheat. Theor Appl Genet 122:1561–1576
Huang T, Qu B, Li H-P, Zuo Z-X, Liao Y-C (2012) A maize viviparous 1 gene increases seed dormancy and preharvest sprouting tolerance in transgenic wheat. J Cereal Sci 55:166–173
Jacobsen JV, Barrero JM, Hughes T, Julkowska M, Taylor JM, Xu Q, Gubler F (2013) Roles for blue light, jasmonate and nitric oxide in the regulation of dormancy and germination in wheat grain (Triticum aestivum L.). Planta 238:121–138
Kato K, Nakamura W, Tabiki T, Miura H, Sawada S (2001) Detection of loci controlling seed dormancy on group 4 chromosomes of wheat and comparative mapping with rice and barley genomes. Theor Appl Genet 102:980–985
King RW (1989) Physiology of sprouting resistance. In: Derera NF (ed) Preharvest field sprouting in cereals. CRC Press Inc, Boca Ratan, pp 27–60
Kondhare KR, Kettlewell PS, Farrell AD, Hedden P, Monaghan JM (2012) Effects of exogenous abscisic acid and gibberellic acid on pre-maturity α-amylase formation in wheat grains. Euphytica 181:51–60
Kondhare KR, Kettlewell PS, Farrell AD, Hedden P, Monaghan JM (2013) The role of sensitivity to abscisic acid and gibberellin in pre-maturity α-amylase formation in wheat grains. J Cereal Sci 58:472–478
Kondhare KR, Hedden P, Kettlewell PS, Farrell AD, Monaghan JM (2014) Use of the hormone-biosynthesis inhibitors fluridone and paclobutrazol to determine the effects of altered abscisic acid and gibberellin levels on pre-maturity a-amylase formation in wheat grains. J Cereal Sci 60:210–216
Lenoir C, Corbineau F, Come D (1986) Barley (Hordeum vulgare) seed dormancy as related to glumella characteristics. Physiol Plant 68:301–307
Lenton JR, Gale MD (1987) Hormonal changes during cereal grain development. In: Mares DJ (ed) 4th international symposium on pre-harvest sprouting in cereals. Westview Press, Boulder, pp 253–264
Liu S, Bai G (2010) Dissection and fine mapping of a major QTL for preharvest sprouting in white wheat Rio Blanco. Theor Appl Genet 121:1395–1404
Liu S, Sehgal SK, Li J, Lin M, Trick HN, Yu J, Gill BS, Bai G (2013a) Cloning and characterization of a critical regulator for preharvest sprouting in wheat. Genetics 195:263–273
Liu A, Gao F, Kanno Y, Jordan MC, Kamiya Y, Seo M, Ayele B (2013b) regulation of wheat seed dormancy by after-ripening is mediated by specific transcriptional switches that induce changes in seed hormone metabolism and signalling. PLoS One 8:e56570
Mares DJ (1983) Investigation of the preharvest sprouting resistance mechanisms in some Australian wheats. In: Kruger JE, LaBerge DE (eds) Third international symposium on preharvest sprouting in cereals. Westview Press Inc., Boulder Co, pp 59–65
Mares DJ (1984) Temperature dependence of germinability of wheat (Triticum aestivum L.) grain in relation to pre-harvest sprouting. Aust J Agric Res 35:115–128
Mares DJ (1987) Preharvest sprouting in white grained wheats. In: Mares DJ (ed) Fourth international symposium on preharvest sprouting in cereals. Westview Press Inc., Boulder Co, pp 64–74
Mares DJ (1989) Preharvest sprouting damage and sprouting tolerance: Assay methods and instrumentation. In: Derera NF (ed) Preharvest sprouting in cereals. CRC Press Inc., Boca Raton, pp 129–170
Mares DJ (1993) Genetic studies of sprouting tolerance in red and white wheats. In: Walker-Simmons MK, Ried JL (eds) Pre-Harvest Sprouting in Cereals 1992. American association of cereal chemists, St Paul, pp 21–29
Mares DJ (1999) The seed coat and dormancy in wheat grains. In: Weipert D (ed) Eighth international symposium on pre-harvest sprouting in cereals 1998. Association of Cereal Research, Federal Centre for Cereal, Potato and Lipid Research, Detmold, pp 77–81
Mares DJ, Mrva K (2001) Mapping quantitative trait loci associated with variation in grain dormancy in Australian wheat. Aust J Agric Res 52:1257–1265
Mares DJ, Mrva K (2008a) Genetic variation for quality traits in synthetic wheat germplasm. Aust J Agric Res 59:406–412
Mares D, Mrva K (2008b) Late-maturity a-amylase: low falling number in wheat in the absence of preharvest sprouting. J Cereal Sci 47:6–17
Mares DJ, Mrva K, Cheong J, Williams K, Watson B, Storlie E, Sutherland M, Zou Y (2005) A QTL located on chromosome 4A associated with dormancy in white- and red-grained wheats of diverse origin. Theor Appl Genet 111:1357–1364
Mares DJ, Rathjen J, Mrva K, and Cheong J (2009) Genetic and environmental control of dormancy in white-grained wheat (Triticum aestivum L.) Euphytica 168:311-318.
Mares DJ, Rathjen J, Mrva K, Cheong J, Oldach K (2012). Germinability during grain development and after-ripening in wheat lines carrying different alleles at dormancy QTL. J Nyachiro (ed). In: 12th International Symposium on Preharvest Sprouting in Cereals, Canada 2012 Abstract
McCarty DR, Carson CB, Stinard PS, Robertson DS (1989) Molecular analysis of vívíparous-I: an abscisic acid insensitive mutant of Maize. Plant Cell 1:523–532
McKibbin RS, Wilkinson MD, Bailey PC, Flintham JE, Andrew LM, Lazzeri PA, Gale MD, Lenton JR, Holdsworth MJ (2002) Transcripts of Vp-1 homeologues are misspliced in modern wheat and ancestral species. PNAS 99:10203–10208
Mohler V, Albrecht T, Mrva K, Schweizer G, Hartl L (2014) Genetic analysis of falling number in three bi-parental common winter wheat populations. Plant Breed 133:448–453
Mori M, Uchino N, Chono M, Kato K, Miura H (2005) Mapping QTLs for grain dormancy on wheat chromosome 3A and the group 4 chromosomes, and their combined effect. Theor Appl Genet 110:1315–1323
Mrva K, Mares DJ (1996a) Inheritance of late maturity α-amylase in wheat. Euphytica 88:61–67
Mrva K, Mares DJ (1996b) Expression of late maturity α-amylase in wheat containing gibberellic acid insensitivity genes. Euphytica 88:68–76
Mrva K, Mares DJ (1999) Regulation of high pI α-amylase synthesis in wheat aleurone by a gene(s) located on chromosome 6B. Euphytica 109:17–23
Mrva K, Mares DJ (2001) Quantitative trait locus analysis of late maturity α-amylase in wheat using the doubled haploid population Cranbrook x Halberd. Aust J. Agric Res 52:1267–1273
Mrva K, Wallwork M, Mares DJ (2006) a-Amylase and programmed cell death in aleurone of ripening wheat grains. J Expt Botany 57:877–885
Mrva K, Mares DJ, Cheong J (2008) Genetic mechanisms involved in late maturity α-amylase in wheat. In: Appels R, Eastwood R, Lagudah E, Langridge P, MacKay M, McIntyre L, Sharp P (eds). Proceedings of the 11th International Wheat Genetics Symposium, Brisbane 2008. Sydney University Press pp 940–942
Mrva K, Cheong J, Yu B, Law HY, Mares DJ (2009) Late maturity α-amylase in synthetic hexaploid wheat. Euphytica 168:403–411
MrvaK, Mares DJ(1996) Control of late maturity α-amylase synthesis compared to enzyme synthesis during germination. In: Noda K, Mares DJ (eds) Preharvest Sprouting in Cereals 1995. Centre for Academic Societies Osaka, pp 419–426
Muncvold JD, Tanaka J, Benscher D, Sorrells ME (2009) Mapping quantitative trait loci for preharvest sprouting resistance in white wheat. Theor Appl Genet 119:1223–1235
Nakamura S, Abe F, Kawahigashi H, Nakazono K, Tagiri A, Matsumoto T, Utsugi S, Ogawa T, Handa H, Ishida H, Mori M, Kawaura K, Ogihara Y, Miura H (2011) A Wheat homolog of MOTHER OF FT AND TFL1 acts in the regulation of germination. Plant Cell 23:3215–3229
Noda K, Kawabata C, Kawakami N (1993) Wheat grain imbibition at low temperatures and embryo responsiveness to ABA. In: Walker-Simmons MK, Ried JL (eds) Preharvest Sprouting in Cereals 1992. American Association of Cereal Chemists, St Paul, pp 367–372
Noda K, Matsuura T, Maekawa M, Taketa S (2002) Chromosomes responsible for sensitivity of embryo to abscisic acid and dormancy in wheat. Euphytica 123:203–209
Noll JS, Dyck DL, Czarnecki E (1982) Expression of RL4137 type of dormancy in F1 seeds of reciprocal crosses in common wheat. Can J Plant Sci 62:345–349
Osa M, Kato K, Mori M, Shindo C, Torada A, Miura H (2003) Mapping QTLs for seed dormancy and the Vp1 homologue on chromosome 3A in wheat. Theor Appl Genet 106:1491–1496
Rathjen JR, Strounina EV, Mares DJ (2009) Pathway for Water Movement into Dormant and Non-Dormant Wheat (Triticum aestivum L.) Grains. J Exp Bot 60:1619–1631
Schramm EC, Nelson SK, Kidwell KK, Steber CM (2013) Increased ABA sensitivity results in higher seed dormancy in soft white spring wheat cultivar ‘Zak’. Theor Appl Genet 126:791–803
Somyong S, Munckvold JD, Tanaja J, Benscher D, Sorrells ME (2011) Comparative genetic analysis of a wheat seed dormancy QTL with rice and Brachypodium identifies candidate genes for ABA perception and calcium signalling. Funct Integr Genomics 11:479–490
Tomkinson CL, Lyndon RF, Arnold GM, Lenton JR (1997) The effects of temperature and the Rht3 dwarfing gene on growth, cell extension, and gibberellin content and responsiveness in the wheat leaf. J Exp Bot 48:963–970
Torada A, Koike M (2012) Marker assisted development and evaluation of near isogenic lines for seed dormancy genes on chromosome 3A and 4A in bread wheat. J Nyachiro (ed). In: 12th International Symposium on Preharvest Sprouting in Cereals, Canada 2012. Abstract
Trethowan RM (1995) Evaluation and selection of bread wheat (Triticum aestivum L.) for preharvest sprouting tolerance. Aust J Agric Res 46:463–474
Verrity JCK, Hac L, Skerritt JH (1999) Development of a field enzyme-linked immunosorbent assay (ELISA) for detection of a-amylase in preharvest-sprouted wheat. Cereal Chem 76:673–681
Walker-Simmons MK (1987) ABA levels and sensitivity in developing wheat embryos of sprouting resistant and susceptible cultivars. Plant Physiol 84:61–66
Warner RL, Kudrna DA, Spaeth SC, Jones SS (2000) Dormancy in white-grain mutants of Chinese Spring wheat (Triticum aestivum L.). Seed Sci Res 10:51–60
Williamson PM (1997) Black point of wheat: in vitro production of symptoms, enzymes involved, and association with Alternaria alternate. Aust J Agric Res 48:13–19
Wilson ID, Barker GLA, Lu C, Coghill JA, Beswick RW, Lenton JR, Edwards KJ (2005) Alteration of the embryo transcriptome of hexaploid winter wheat (Triticum aestivum cv. Mercia) during maturation and germination. Funct Integr Genomics 5:144–154
Yang Y, Zhao XL, Xia LQ, Chen XM, Xia XC, Yu Z, He H, Roder M (2007) Development and validation of a Viviparous-1 STS marker for pre-harvest sprouting tolerance in Chinese wheats. Theor Appl Genet 115:971–980
Yang Y, Zhang CL, Liu SX, Sun YQ, Meng JY, Xia LQ (2014) Characterization of the rich haplotypes of Viviparous-1A in Chinese wheats and development of a novel sequence tagged site marker for pre-harvest sprouting resistance. Mol Breed 33:75–88
Zhang Y, Miao X, Xia X, He Z (2014) Cloning of seed dormancy genes (TaSdr) associated with toleranceto pre-harvest sprouting in common wheat and development of a functional marker. Theor Appl Genet 127:855–866
Acknowledgments
The authors gratefully acknowledge funding support from the Australian Grains Research and Development Corporation and thank Dr Peter Chandler, CSIRO, Australia for helpful discussion of the possible role of gibberellins in LMA.
Conflict of interest
We declare that we have no conflict of interest.
Author information
Authors and Affiliations
Corresponding author
Electronic supplementary material
Below is the link to the electronic supplementary material.
Rights and permissions
About this article

Cite this article
Mares, D.J., Mrva, K. Wheat grain preharvest sprouting and late maturity alpha-amylase. Planta 240, 1167–1178 (2014). https://doi.org/10.1007/s00425-014-2172-5
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00425-014-2172-5