
Abstract
Immunomodulation is a means to modulate an organism’s function by antibody production to capture either endogenous or exogenous antigens. We have recently succeeded in obtaining gibberellin (GA)-deficient phenotypes in Arabidopsis thaliana by using anti-bioactive GA antibodies. In this study, a single-chain antibody (scFv) against GA24, a precursor GA, was utilized to repress the biosynthesis of bioactive gibberellins. Stable accumulation of the scFv in endoplasmic reticulum (ER) was achieved by being produced as a fusion with GFP as well as KDEL ER-retention signal. The transgenic plants showed GFP fluorescence in the reticulate cortical ER network in epidermal cells. The GFP-scFv fusion produced in plants maintained its binding activity. The transgenic plants showed GA-deficient phenotypes, including reduced rosette leaf development, delayed flower induction and reduced stem elongation of the main culm, especially in the early stage of inflorescence growth. Contrarily, stem elongation of the main culm at a later stage, or that of lateral shoots was much less affected by scFv production. These phenotypes were different from anti-bioactive GA scFv-producing lines, whose stem elongation was continuously repressed throughout the inflorescence development. The GA-deficient phenotypes were recovered by treatment with GA24 and bioactive GA4, the latter being more effective. The transgenic lines contained conspicuously higher endogenous GA24 and clearly less GA4 than wild-type plants. The expression of GA 20-oxidase and GA 3-oxidase genes, which are feedback-regulated by GA signaling, were up-regulated in those plants. These results demonstrate that the scFv trapped GA24 in ER and inhibited metabolism of GA24 to bioactive GA4.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
The methodology to modulate plant functions by antibody production in planta is called ‘immunomodulation’. Since Owen et al. (1992) showed that plants producing anti-phytochrome antibody have decreased responsiveness to light, immunomodulational studies in plants have seen a steady increase in the number of antibodies produced, including those against phytohormones to repress phenomena regulated by phytohormones such as abscisic acid (Artsaenko et al. 1995; Phillips et al. 1997; Strauss et al. 2001; Wigger et al. 2002) and jasmonic acid (ten Hoopen et al. 2007), metabolic enzymes to modulate pigment metabolism (De Jaeger et al. 1999) and starch synthesis (Jobling et al. 2003), plant viral coat proteins to confer virus resistance (Tavladoraki et al. 1993; Voss et al. 1995; Fecker et al. 1997; Zimmermann et al. 1998), herbicides to confer herbicide resistance (Eto et al. 2003; Horsman et al. 2007), and against small heat shock proteins that prevent the normal heat stress response (Miroshnichenko et al. 2005).
Recently, we succeeded in obtaining GA-deficient phenotypes in Arabidopsis such as reduced leaf size, delayed bolting, reduced inflorescence length and decreased germination, by producing a single-chain variable fragment (scFv), a recombinant form of antibody, recognizing bioactive GAs (Suzuki et al. 2008). When the scFv was localized in endoplasmic reticulum (ER), it was most stably accumulated and gave clear GA-deficient phenotypes. The transgenic plants responded to exogenous GA4 and contained 15–40 times higher endogenous GA4 than wild-type plants. They also showed changes in expression levels of enzyme genes involved in the regulation of GA4 level similarly to the GA-deficient mutants. These results clearly demonstrated that the level of free functional GA4 was decreased by being trapped with scFv in ER to the extent that GA-deficient phenotypes were created. A dramatic increase in the total sum of GA4 (free plus scFv-bound) was attributable to the enhanced GA biosynthesis caused by repressed GA action, and a reduction in GA4 catabolism due to protection by scFv binding in ER.
While the above study was a good example of immunomodulation where GA function was inhibited by the antibody to prevent bioactive GAs from interacting with the GA receptors, there is another possible mode of immunomodulation for GA function, which is inhibition of GA biosynthesis by using antibodies against biosynthetic precursors of bioactive GAs. We previously reported the preparation of dwarf tobacco plants producing an antibody, which was raised against BSA-conjugated GA24 (Shimada et al. 1999). The antibody was highly reactive with GA24 and GA19, which are aldehyde-type GAs located two steps upstream of the bioactive GA4 and GA1, respectively. While the antibody showed no cross-reactivity with GA20, GA1 and GA4, all having γ-lactone on the A ring of the GA skeleton, GA17 and GA13, having a C-20 carboxyl group, nor with GA53 having a C-20 methyl group, it did show reactivity to GA15, which has a δ-lactone on the A ring. The antibody was also not reactive with GA24 methyl ester. Based on these results, it was argued that the antibody critically recognized either C-19 → C-20 lactol or δ-lactone on the A ring, and the dissociated form of C-7 carboxyl group on the B ring (Shimada et al. 1999). The observed dwarf phenotype for transgenic tobacco was considered to be caused by capture of biosynthetic precursor GAs by the scFv, because GA1, the major bioactive GA in tobacco, could not be detected from the transgenic tobacco plants whereas wild-type plants contained a detectable level of GA1. However, the results were preliminary in the sense that statistical significance was not evaluated for the dwarf phenotype. Afterwards, we tried to produce the same scFv in Arabidopsis, so as to demonstrate the significant effect of the antibody to modulate plant growth, and also to show that the effect is universal among different plant species. However, the antibody was not stably accumulated in Arabidopsis and conferred no phenotypes to the transgenic plants. In this study, we again tried to produce the same scFv as a fusion with green fluorescence protein (GFP) in ER along the lines of the previous study where GFP-fused anti-bioactive GA scFv was stably accumulated in Arabidopsis ER. Previous immunomodulational studies targeting small molecules supported the idea that the effective inhibition of the target function can be obtained by producing antibodies in the ER (Artsaenko et al. 1995; Eto et al. 2003; Suzuki et al. 2008). This can be interpreted as antibody being generally more stable in ER than in any other compartment tested, and that targets of antibodies with low molecular weight and relatively high hydrophobicity can pass through membrane systems and reach antibodies in ER. Because antigens are sequestered in ER by antibodies and cannot bind their receptors in other compartments, this mode of immunomodulation is called ‘mis-targeting’ of antigens (Conrad and Manteuffel 2001). Since the anti-GA24 scFv was stably accumulated in ER but not in cytosol in tobacco (Shimada et al. 1999), we followed the mis-targeting system in this study. Consequently, the anti-precursor GA scFv was stably accumulated and gave clear phenotypes in Arabidopisis. Interestingly, the transgenic plants producing anti-GA4 scFv and anti-GA24 scFv showed different severity depending on the growth stages, although both of them showed GA-deficient phenotypes as a whole.
Materials and methods
Construction of the plant expression vector
The anti-GA24 scFv gene was PCR-amplified with a template plasmid pHlL24B (Shimada et al. 1999), with a primer set for integrating Bsp1407I and NotI sites (fw, 5′-ggtgtaca[Bsp1407I]tggaggtgaagctggtggagtc-3′; rv, 5′-tgcggccgc[NotI]tacagttggtgcagcatcagccc-3′). The scFv fragment was replaced with a scFv gene encoding anti-GA4 scFv using Bsp1407I and NotI sites in pBI-ER-D, a binary vector constructed previously (Suzuki et al. 2008).
Plant transformation
The binary vector was transferred into Agrobacterium tumefaciens strain LBA4404 (pAL4404), which was used to transform Arabidopsis plants (Arabidopsis thaliana L. Heynh., Brassicaceae, accession Columbia, from Arabidopsis Biological Resource Center, Ohio State University, Columbus, OH, USA) as previously described (Bechtold et al. 1993). Transgenic plants (T1) were selected on Murashige and Skoog (MS) agar medium containing 100 μg/ml of kanamycin. Homozygous lines were established from the independent T1 lines whose next generation showed a 3 to 1 segregation for kanamycin resistance and sensitivity, and which were used for all subsequent analyses.
Plant growth conditions
Surface-sterilized seeds were sown on sterilized MS media containing 3% (w/v) sucrose and 0.8% (w/v) agar (Wako, Osaka, Japan). The seeds were moved to a growth chamber at 23°C with continuous fluorescent light (35 μmol m−2 s−2) after chilling at 4°C for 3 days.
Microscopic analysis of GFP fluorescence from GFP-scFv proteins
The subcellular localization of GFP-scFv in transgenic plants was analyzed by observing GFP fluorescence under a confocal laser scanning microscope (IX81/FV500, Olympus, Tokyo, Japan). Excitation at 488 nm was provided by Ar ion laser, and fluorescence emission was collected at 500–530 nm. Images were acquired with Olympus FluoView software. A fluorescence microscope (BX60, Olympus) was also used for GFP observation with either a U-MWIB/GFP or a U-MNIBA filter (Olympus).
Radioimmunoassay and Western blotting
Rosette leaves (200 mg) was frozen in liquid nitrogen and ground to a fine powder, and protein was extracted from the powder with 1 ml of 50 mM Tris-HCl (pH 8.0) containing 200 mM NaCl, 5 mM EDTA and 0.1% Tween 20 and one tablet of proteinase inhibitor mixture (Complete Mini, Roche, Mannheim, Germany) for 10 mL of the buffer. The binding activity of scFv from the transgenic plants to GA24 was detected by radioimmunoassay. The extract (100 μL) was mixed with 3H-labeled GA24 (10,000 dpm) in PBS containing 180 μL rabbit normal serum, and incubated at 4°C overnight. For competitive radioimmunoassay, an excess of non-labeled GA24 was added to the mixture to give a 0.17 μM final concentration. To measure the unspecific binding, the protein extract was eliminated from the incubation reaction.
The presence of scFv protein was detected by Western blotting. Twenty micro liters of the protein extract was applied to sodium dodecyl sulfate-polyacrylamide gel electrophoresis. Proteins were electrotransferred onto a nitrocellulose membrane, and a c-myc tag fused to scFv was detected with the anti-c-myc antibody (9E10 mouse ascites 1,000 times diluted with PBS containing 2% skimmed milk) (Evan et al. 1985) and HRP-conjugated anti-mouse antibody (5,000 times diluted with PBS; GE Healthcare, Uppsala, Sweden). The HRP activity was detected by an ECL Plus Western blotting detection system (GE Healthcare).
Quantification of endogenous gibberellins and real-time RT-PCR analysis
The aerial part of wild-type plants (4.0 g), A24-2 (3.9 g) and A24-3 (3.5 g) was spiked with 50 ng of deuterium-labeled GA4 and GA24 (purchased from Dr. Mander, Australian National University), and homogenized in methanol. The extract was filtered and concentrated in vacuo. The concentrate was acidified with hydrogen chloride to pH 3 and extracted with ethyl acetate, which was then extracted with 0.5 M potassium phosphate buffer (pH 8.5). The aqueous layer was again extracted with ethyl acetate after being adjusted to pH 3. The obtained ethyl acetate acidic fraction was dried over sodium sulfate and concentrated to dryness. The concentrate was dissolved in a small volume of 50 mM potassium phosphate buffer (pH 8.0) and loaded onto a column packed with insoluble polyvinylpyrrolidone (1 g, Tokyo Kasei, Tokyo, Japan), prepared and equilibrated with the same phosphate buffer, and the GA fraction was obtained as the first 10-mL elution with the same buffer. The resulting GA fraction was acidified to pH 3.0 with hydrogen chloride and loaded onto an OASIS HLB cartridge (500 mg, Waters, Milford, MA, USA). After washing with 6 mL of 20% methanol containing 1% acetic acid, the GAs were recovered by elution with 12 mL of 60% methanol containing 1% acetic acid. The eluate was concentrated to dryness, loaded onto Sep-Pak Silica (500 mg, Waters) equilibrated with chloroform–ethyl acetate (1:1, v/v) containing 1% acetic acid, and eluted with 10 mL of the same solvent. The purified GA fraction was analyzed with an API3000 LC-MS/MS device (Applied Biosystems, Foster City, CA, USA). The sample was separated in Senshu Pak Pegasil ODS (2.0 mm i.d. × 150 mm, Senshu Scientific, Tokyo, Japan). Elution was performed as follows, with solvents A [0.1% (v/v) acetic acid in water] and B [0.1% (v/v) acetic acid in acetonitrile]: 0–2 min, isocratic with 10% B; 2–20 min, linear gradient from 10% B to 100% B; 20–24 min, isocratic elution with 100% B. The flow rate was 0.2 mL min−1. MS-MS analysis was carried out in the negative-ion mode with a turbo-ionspray inlet system. GA4, GA24 and their internal standards, d2-GA4 and d2-GA24, were detected by the combination of the primary and the secondary ions at m/z 331/257 for GA4, m/z 345/257 for GA24, m/z 333/259 for d2-GA4, and m/z 347/259 for d2-GA24, respectively, in the multiple-reaction monitoring mode.
Real-time RT-PCR analysis exactly followed the method in our previous report (Suzuki et al. 2008).
Results
Preparation of transgenic Arabidopsis producing GFP-scFv fusion proteins
ScFv is composed of variable domains of an H chain (VH) and an L chain (VL) of an immunoglobulin molecule connected with a flexible linker peptide. The scFv gene used in this study had been prepared from a hybridoma cell line producing a monoclonal antibody against aldehyde-type GAs, GA24 and GA19 (Shimada et al. 1999). Here we designate the scFv anti-GA24 scFv because the major GA biosynthetic pathway in Arabidopsis is the early non-3, 13-hydroxylation pathway (Talon et al. 1990; Magome et al. 2004), on which GA4 and GA24 are a bioactive GA and its biosynthetic precursor, respectively, i.e., GA24 is converted to GA9, the immediate precursor of bioactive GA4. Although the antibody is also reactive with GA15, which is the precursor of GA24, it is the minor target of the antibody in Arabidopis (Magome et al. 2004). GFP was fused to the N-terminus of the anti-GA24 scFv, and a c-myc epitope tag and KDEL ER-retention signal to the C-terminus. A cauliflower mosaic virus 35S RNA promoter was used for constitutive expression. More than ten independent transgenic plants were obtained as kanamycin-resistant plants.
Confocal imaging of GFP fluorescence, Western blotting and radioimmunoassay of scFvs produced in plants
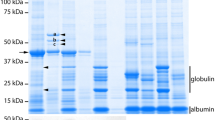
In the initial screening of transgenic plants, some lines here designated as A24-1, 2 and 3 showed clear dwarf phenotypes presumably due to GFP-scFv production (details will be shown later), and showed strong GFP fluorescence. Using these lines, sub-cellular localization of GFP-scFv was examined by confocal laser scanning microscopy. The reticulate shape of fluorescence was observed in the epidermal cells of rosette leaves (Fig. 1a). This reticulate structure is a well-known cortical ER network (Staehelin 1997; Suzuki et al. 2008), indicating that GFP-scFv was successfully targeted to the ER compartment. GFP-scFvs in the extracts from A24-1, 2 and 3 were clearly detected by western blotting at the expected size on SDS-PAGE (Fig. 1b), which were quite different from our previous experiment where scFv could hardly be detected by the same detection method when we tried to produce scFv without fusing with GFP (data not shown). The protein extract from the transgenic line A24-2, which showed a higher GFP-scFv level than the other two lines, was subjected to radioimmunoassay to confirm that the GFP-scFv maintains its binding activity with GA24. After incubation of the protein extract with 3H-labeled GA24, radioactivity was detected in the ammonium sulfate-precipitated protein fraction containing GFP-scFv, and the recovery of 3H-GA24 in the protein fraction was decreased by an excess non-labeled GA24 as a competitor of 3H-GA24 to the level of the unspecific binding (Fig. 1c). As the extract from wild-type plants showed no significant binding, the binding activity observed in the A24-2 plants could be attributed to scFv produced in the plants. These results indicate that functional scFv was successfully produced in Arabidopsis as fusion with GFP in the ER compartment.

Production of GFP-scFv protein in transgenic plants. a The green fluorescence from GFP-scFv protein was observed with a confocal laser scanning microscope, showing subcellular localization of the protein in the reticulate cortical ER network in epidermal cells. Bar 20 μm. b The production of GFP-scFv was detected by western blotting. The protein extract from rosette leaves (two individual plants from the same transgenic line) corresponding to approximately 4 mg fr. wt. was loaded into each lane and detected with anti-c myc-tag antibody. WT wild-type plant. c The binding activity of scFv in plant extract of A24-2 line toward GA24 was detected by radioimmunoassay using 3H-labeled GA24 as a tracer. The level of unspecific binding shown as a dashed line was determined by eliminating the plant extract from the reaction mixture. The binding above the level of unspecific binding was shown to be competitively replaced by excess GA24. The assay was also performed with wild-type plant extract to show that the binding activity in the transgenic plant was derived from the scFv
Phenotypes of transgenic plants
The effects of scFv production on conferring GA-deficient phenotypes to plants were examined in detail using the transgenic lines with a high accumulation of GFP-scFv (A24-1, 2 and 3). Figure 2a shows the typical appearance of the A24-2 line and a wild-type plant at 58 days after germination, the point in time when both lines had already stopped elongation. The plant height of A24-2 was less than one-half of the wild-type plant. The dwarf phenotype was statistically significant for all three lines, A24-1, 2 and 3, and the severity of the dwarf phenotype was approximately correlated with the accumulation level of GFP-scFv protein (Figs. 1b, 2b). Analysis of the temporal fluctuation of the A24-2 plant height showed that stem elongation was extremely slow during the first several days after bolting, whereas the elongation rate at a later stage was almost comparable to that of wild-type plants (Fig. 2c).

GA-deficient phenotypes of the transgenic plants producing anti-GA24 scFv. a Typical appearance of plants producing anti-GA24 scFv after transition to reproductive growth. A transgenic plant line (A24-2) and a wild-type plant (WT) 58 days after germination are shown. Bar, 3 cm. b Comparison of the inflorescence length of each transgenic line at 45 days after germination. Each data represents the mean ± SD of 10 plants. ** indicates significant difference compared to wild-type plants at P < 0.01 by Welch’s t-test. c Temporal fluctuation of plant height of wild-type plants and A24-2 plants. Each point represents the mean ± SD of 10 plants. d The A24-2 plants which started flowering are shown beside a WT plant at the same age (42 days after germination). Bar 3 cm. e Typical appearance of a A24-2 plant and a WT plant during vegetative growth (28 days after germination). Bar 3 cm. f Comparison of the longest rosette leaf length between A24-2 and WT plants. Each data represents the mean ± SD from 30 to 40 plants at 28 days after germination. ** indicates significant difference compared with WT at P < 0.01 by Welch’s t-test. g Temporal fluctuation of the longest rosette leaf length of A24-2 and WT. Each point represents the mean ± SD of 10 plants
Another GA-deficient phenotype was observed as a delay in bolting which was accompanied with increased rosette leaf number, a typical late flowering phenotype. Bolting of A24-2 was 6.3 days later than in wild-type plants and had about four more leaves (Table 1). Table 1 also shows the day at first flower opening and the inflorescence length at that time. The delay of the first flower opening in A24-2 (3.8 days) was not as big as that of bolting (6.3 days), because flowers in the A24-2 line started to open soon after bolting on a shorter inflorescence than wild-type plants (Table 1; Fig. 2d). It is apparent that the antibody is more effective in repressing flower induction than flower development.
Extension of rosette leaves of the transgenic line was also repressed by antibody production. The leaves had shorter petioles and their overall size was clearly smaller than wild-type plants (Fig. 2e, f). Their leaves also had a darker green color (Fig. 2e) and were less flexible, which was reminiscent of a GA-deficient mutant such as ga1-3 (Sun and Kamiya 1994). The time course of rosette leaf expansion indicated that the leaves of A24-2 continued to extend for a longer period than wild-type plants (Fig. 2g).
Verification of capture of GA24 by GFP-scFv causing repressed GA4 production
To verify that the GA-deficient phenotypes can be attributed to the repressed biosynthesis of GA4 caused by capture of the precursor GA24 by the produced scFv, first the ability of the plants to respond to GA4 was confirmed. When an excess amount of GA4 was applied to the plants by spraying GA4 solution, the GA-deficient phenotypes, including reduced rosette size, delay of bolting and reduced stem elongation were clearly rescued (Fig. 3a, b), indicating that the A24-2 plants retained their responsiveness to bioactive GA. In this recovery assay using an excess amount of GA, the effects of treating with GA4 and GA24 were not significantly different (data not shown). However, when a small amount of GA solution was applied to the tip of a rosette leaf, the effect of GA4 on the recovery of stem elongation was much bigger than that of GA24 (Fig. 3c). There were two possible explanations for this difference. One was that GA24 is less active than GA4 regardless of the scFv production because the conversion ratio of GA24 to bioactive GA4 is not 100%, and/or transport efficiency of GA24 is lower than that of GA4. The other possibility was that the exogenous GA24 was captured by scFv in the treated rosette leaf similarly to the endogenous GA24, and only part of the applied GA24 was converted to GA4. To examine the former possibility, ga1-3, a GA biosynthetic mutant, was treated with GAs in an identical manner. When applied to the tip of a rosette leaf, GA24 was even more effective than GA4 in recovering stem elongation of ga1-3 (Fig. 3d). This result is consistent with the observation that bioactive GA1 was less effective than its precursors on shoot elongation of morning glory when they were treated onto the cotyledons (Yang et al. 1995). The precursor GA20 has also been shown to be more effectively transported from lower leaves to upper apices than bioactive GA1 in pea (Proebsting et al. 1992). The present result is highly suggestive that the lower effectiveness of GA24 to recover stem elongation of A24-2 plants was due to the accumulated scFv, which captured the applied GA24 and inhibited conversion to GA4.

Verification for capture of GA24 by GFP-scFv causing repressed GA4 production. a Typical appearance of A24-2 plants treated with GA4. The A24-2 plants at 35 days after germination were sprayed with 100 μM GA4 or mock-treated. The plants were recorded 5 days after the treatment. b Inflorescence lengths of A24-2 lines sprayed with GA4 or mock solution. Inflorescence length was measured 4 days after the treatment as in a. ** indicates significant difference compared with mock-treated plants at P < 0.01 by Welch’s t-test. c Inflorescence lengths of A24-2 lines whose rosette leaves were treated with GA4, GA24 or mock solution. Ten microliters of 100 μM GA4 or GA24 solution was applied to the tip of the longest rosette leaf of each A24-2 plant at 40 days after germination which had not started bolting. The inflorescence lengths were measured 5 days after the treatment. ** indicates significant difference compared with both mock-treated and GA24-treated plants at P < 0.01 by Welch’s t-test. The GA24-treatment showed no significant difference with mock-treatment. d Comparison of GA24 and GA4 treatment on recovery of the GA-deficient phenotypes of ga1-3. Half a microliter of 100 μM GA4 or GA24 solution was applied to ga1-3 as in c. Note that mock-treated plants produced no elongated inflorescence. ** indicates significant difference compared with GA4-treated plants at P < 0.01 by Welch’s t-test. e Endogenous levels of GA24 and GA4 in wild-type plants (WT), A24-1 and A24-2 lines. Aerial parts of each plant line were extracted, purified and analyzed by LC-MS/MS with deuterium-labeled GA24 and GA4 as internal standards. The numerals are GA levels (ng/FW)
If capture of GA24 by scFvs in the ER is effective enough to repress its conversion to bioactive GA4 and cause GA-deficient phenotypes, the transgenic plants are expected to have a higher level of GA24 and a lower level of GA4 than wild-type plants. The LC-MS/MS analysis showed that the endogenous level of GA24 in A24-2 and A24-3 was conspicuously higher than in wild-type plants, while the level of GA4 was less than 20% of that in wild-type plants (Fig. 3e). These results strongly suggest that the GA-deficient phenotypes were caused by decreased biosynthesis of bioactive GA4 due to repression of GA24 metabolism as a result of its capture by the antibody.
To obtain a supportive data to show that the transgenic plants are in the GA deficient state, transcript levels of GA biosynthetic genes, AtGA20ox1 and AtGA3ox1, which are feedback-regulated by activated GA signaling, were analyzed. Quantitative real-time RT-PCR analysis showed that expression of AtGA3ox1 and AtGA20ox1 are significantly up-regulated in the A24-2 line (Fig. 4). This result also indicated that the conspicuous accumulation of GA24 in A24-2 plants was due not only to repressed metabolism of GA24 by binding of scFv but also to its enhanced biosynthesis.

The effect of GFP-scFv production on gene expression of GA biosynthetic enzyme genes. The transcript level of AtGA3ox1 and AtGA20ox1 was obtained by real-time RT-PCR, and the level in A24-2 was shown as a relative value against that in wild-type plants (WT = 100%). AtActin2 was used as the internal control in the analysis. Each data column represents the mean + SD of 3 quantification results using cDNAs independently prepared from 3 plants. ** and * indicate significant differences compared with WT at P < 0.01 and P < 0.05, respectively, by Student’s t-test
Differences of phenotypes between plants producing anti-GA4 scFv and anti-GA24 scFv
Although, the overall phenotypes caused by anti-GA4 scFv production and anti-GA24 scFv were similar in the sense that both antibodies gave GA-deficient phenotypes for rosette leaf size, flower induction and stem elongation, there were apparent differences between them in the extent of effects depending on the growth stages. During the early stage of vegetative growth, rosette leaf development was more strongly repressed by anti-GA24 scFv than by anti-GA4 scFv. Figure 5a shows the typical appearance of A24-2 and E-ER-4 plants, one of the transgenic Arabidopsis lines producing anti-GA4 scFv (Suzuki et al. 2008) 30 days after germination when no line, including wild-type plants, had started to bolt. At this early growth stage, A24-2 plants have smaller rosette leaves than E-ER-4 (Fig. 5b) and their green color was slightly darker (Fig. 2e). The effect of anti-GA24 scFv was also bigger than anti-GA4 scFv on delaying flower indution; the bolting time of A24-2 was more than 6 days later than wild-type plants (Table 1) whereas that of E-ER-4 was only about two days later (Suzuki et al. 2008). Figure 5c shows the time when A24-2 plant started to bolt. At this time, a wild-type plant and E-ER-4 plants had already started their elongation of the inflorescence stem. Note that the rosette leaf size of A24-2 is even bigger than E-ER-4 (Fig. 5c, d), because rosette leaves of A24-2 continued to grow for longer period than wild-type plants and E-ER-4 (cf. Fig. 2g), although it is not clear if the longer expansion growth of rosette leaves is associated with the delayed bolting.

Comparison of phenotypes between transgenic lines producing anti-GA4 scFv (E-ER-4) and those producing anti-GA24 scFv (A24-2). a Typical appearance of A24-2, E-ER-4 and wild-type plants (WT) 20 days after germination. b Longest rosette leaf lengths of A24-2, E-ER-4 and WT 20 days after germination. ** indicates significant difference compared with WT at P < 0.01 by Welch’s t-test. The longest rosette size for A24-2 was significantly smaller than that for E-ER-4 at P < 0.01. c Typical appearance of A24-2, E-ER-4 and WT 42 days after germination. d Longest rosette leaf lengths of A24-2, E-ER-4 and WT 42 days after germination. ** and * indicate significant differences compared with WT at P < 0.01 and P < 0.05, respectively by Welch’s t-test. The longest rosette size for E-ER-4 was significantly smaller than that for A24-2 at P < 0.01. e Typical appearance of A24-2 and E-ER-4 58 days after germination. f The value of lateral branch length divided by main culm length as an index for the differential effect of antibody production on repressed growth of main culm and that of lateral branch. The length of main culm and that of longest lateral branch was measured 58 days after germination, and their ratio is shown. * indicates significant difference compared with WT at P < 0.05 by Welch’s t-test. The value for E-ER-4 was not significantly different from that of WT
As described before, the inflorescence stem elongation of A24-2 line was extremely slow during the first several days after bolting, whereas the elongation rate at a later stage was recovered almost to the level of wild-type plants (Fig. 2c). In contrast, the elongation rate of E-ER-4 line was constantly about half of that of wild-type plants (Suzuki et al. 2008). Consequently, the internode length between fruits, which is an index of stem elongation at a later stage of inflorescence growth after flowering, of E-ER-4 was shorter than that of A24-2 (Fig. 5e). The effect of anti-GA24 scFv on repressing elongation of lateral branches was also smaller than anti-GA4 scFv. Therefore, the value of lateral branch length divided by main culm length for A24-2 was larger than the value for wild-type plants, whereas that of E-ER-4 was not significantly different from that of wild-type plants (Fig. 5f).
As a whole, anti-GA24 scFv was more repressive than anti-GA4 scFv on growth during the early stages including rosette leaf development, flower induction and inflorescence stem elongation just after bolting, and less repressive on later inflorescence growth, whereas anti-GA4 scFv showed a constant mild effect on repressing plant growth. These tendencies for the extent of antibody effects were reproducibly observed for other transgenic lines (data not shown).
Discussion
In our previous study, we showed that immunomodulation could be applied to modulate GA-regulated plant phenomena by using anti-bioactive GA4 antibody. The antibody in that study was supposed to prevent bioactive GAs from binding with the GA receptor. In this study, we evaluated the possibility to immunomodulate biosynthesis of GA using anti-precursor GA antibody. To this end, we prepared Arabidopsis plants producing anti-GA24 scFv fused to GFP along the lines of the previous study where GFP-fused anti-GA4 scFv was stably accumulated in Arabidopsis ER. Consequently, the GFP-fused anti-GA24 scFv was also successfully accumulated to a level high enough to obtain GA-deficient phenotypes. This result further supported the usefulness of ‘mis-targeting’ of antigens to block their functions, when antibodies are more stable in a specific compartment such as ER and antigens can reach antibodies. It is likely that GA24 synthesized in cytosol was diffusible across ER membrane system to the lumen of ER, and was captured by the scFv, resulting in shortage of biosynthetic precursor of bioactive GA4. Since the same scFv was not stably accumulated when it was produced without being fused with GFP, it seems that GFP worked to stabilize the fusion protein in ER. Scheller et al. (2006) showed that elastin-like polypeptides fused to the C-terminus of scFv increased the accumulation level of scFv in ER to a large extent. Although KDEL ER-retention signal was shown to stabilize scFvs in the cytosol where scFvs harbouring intramolecular disulphide bonds are generally unstable because of its reducing condition (Schouten et al. 1997), there is only limited information how to stabilize scFvs in other compartments. Because fusing scFvs with other proteins or peptides is simpler than modifying scFv molecules by mutagenesis, it is quite advantageous to establish the fusing component which can be generally applied to stabilize scFvs, not only for the immunomodulational study but also for obtaining antibodies for pharmaceutical use.
The rescue of phenotypes of A24-lines by GA4 showed that the transgenic plants could retain their responsiveness to GA. The fact that GA4 was more effective than GA24 in rescuing the phenotypes suggests that the scFvs inhibit the conversion of GA24 to GA4. The conspicuous elevation of endogenous GA24 level and the clear reduction of GA4 level in the anti-GA24 scFv-producing plants further supported the hypothesis that the GA-deficient phenotypes were caused by scFvs preventing conversion of GA24 to bioactive GA4. GA15, another possible target of the scFv located in the major early non-3, 13-hydroxylation pathway, was not detected by LC-MS/MS analysis even from the transgenic lines (data not shown). The level of GA15 might be too low to be captured by the scFv. The GA-deficient state was further supported by the expression levels of GA 20-oxidase and GA 3-oxidase genes. Their higher levels in A24-2 than in wild-type plants suggest that the feedback repression of those genes was relieved by the reduction of GA signaling. Therefore, the increased level of GA24 is attributable both to the enhanced biosynthesis of GA24, and to the protection of GA24 by the scFv from metabolizing enzymes. It seems that the protecting effect of scFv is enhanced by the subcellular compartment of the scFv accumulation, i.e., the captured GA24 is separated in ER not only from GA receptors but also from the metabolizing enzymes such as GA 20-oxidases or possibly GA 2-oxidase, which are located in the cytosol. This conspicuous accumulation of target antigen was also observed for GA4 (Suzuki et al. 2008), abscisic acid (Phillips et al. 1997; Strauss et al. 2001) and jasmonic acid (ten Hoopen et al. 2007) in transgenic plants producing scFvs against respective antigens, suggesting that the content of rare endogenous compounds can be elevated by producing antibodies recognizing those compounds.
Although both the present immunomodulational study using anti-precursor GA antibody and the previous study using anti-bioactive GA antibody gave GA-deficient phenotypes by repressed GA biosynthesis and repressed action of bioactive GA, respectively, their phenotypes were apparently different in terms of the extent of phenotypes depending on the growth stages. In summary, the early stage of development including rosette leaf extension, flower induction and early inflorescence growth was more effectively repressed by anti-GA24 scFv than by anti-GA4 scFv, whereas the later inflorescence growth was less effectively repressed by anti-GA24 scFv than by anti-GA4 scFv. Although the extent of phenotype was dependent on the production level of scFv, the tendency of immunomodulated growth was consistent among different transgenic lines, both those producing anti-GA24 scFv and those producing anti-GA4 scFv. The fact that the shoot apical meristem (SAM) before bolting is adjacent to rosette leaves, and the results that indicate that the phenotypes on the SAM, i.e., flower induction and early stem elongation, and on rosette leaves were more severe in A24-lines suggest a possibility that flower induction and early stem elongation depend on GAs biosynthesized in rosette leaves. This is supported by the result that GAs applied to rosette leaves can rescue the GA-deficient phenotypes, late flower induction and reduced stem elongation, of ga1-3 mutant (Fig. 3d). It is well known that GAs are involved in flower induction of long-day (LD) plants. In some plant species, the LD condition was shown to promote GA biosynthesis (Talon and Zeevaart 1990; Talon et al. 1991), but this response is not restricted to any specified tissue. In spinach, GA levels increase in the petiole, blade and also in the shoot tip in response to LD. Spinach GA 20-oxidase, which catalizes the conversion of GA24 or GA19 to GA9 or GA20, the respective immediate precursors of bioactive GA4 or GA1, was shown to be involved in this regulation as a key enzyme; LD increases not only the transcript level but also its protein level (Wu et al. 1996; Lee and Zeevaart 2002). In Arabidopsis, although the GA levels in shoot tips have not been measured, GA contents in rosette leaves were shown to increase in response to LD (Xu et al. 1997). Since the transcript level of GA 20-oxidase increases in the shoot tips of Arabidopsis in response to LD (Xu et al. 1997), it is likely that the biosynthesis of GA in the elongation zone is at least partly responsible for the early stem elongation. It is probable that a precursor GA such as GA24 is translocated from rosette leaves to the SAM and is used for the biosynthesis of GA4 for flower induction and early stem elongation, in which the LD-induced GA 20-oxidase is involved. On the other hand, the mode of GA action for flowering in Arabidopsis grown under short-day condition has been studied in more detail (Eriksson et al. 2006). The authors first showed that GA4 is the active gibberellin in the regulation of flowering based on the observation that GA4 level in shoot apices increased dramatically just before flower initiation, and that GA4 showed a promoting effect on flowering when applied to shoot apices while other biologically active GAs, GA1 and GA5, were inactive. Thereafter they suggested that the GA4 involved in flowering is transported from outside sources based on the fact that the expression levels of GA 20-oxidase and GA 3-oxidase genes remained unchanged during the period when GA4 levels in shoot apices increased. They further confirmed the transportation of exogenously applied GA4 on rosette leaves to shoot apices. All these studies as well as our results suggest the above hypothesis that the GAs synthesized in rosette leaves may be translocated to the SAM and participate in both flower induction and promotion of early stem elongation.
It is difficult to explain why anti-GA24 scFv and anti-GA4 scFv gave different extents of severity depending on the growth stages. It is apparent that merely the level of scFv production or their affinity to antigen cannot account for this difference. Many factors must be taken into account. First, the endogenous GA24 and GA4 are regulated to be at different levels and this regulation is also temporally and spatially specific. This means that the ease with which GA24 and GA4 are captured is different and this difference depends on the kind of tissue and its age. On the other hand, the accumulation level of scFv is also inconsistent despite the fact that gene expression was driven by a CaMV 35S RNA promoter, which is supposed to be constitutively active. Judging from the fluorescence strength, the level of scFv is higher in older tissues such as old rosette leaves and lower in young tissues such as young leaves or shoot tips. Moreover, the competitor of anti-GA24 scFv at capturing targets is different from that of anti-GA4 scFv, i.e., GA 20-oxidase for anti-GA24 scFv and GA receptor for anti-GA4 scFv. Thus, both the relative affinities and the relative amounts of the antibodies and their competitors must be important factors to determine the phenotype severity. The relative locations of the proteins, ER versus cytosol in A24 lines, and ER versus nucleus in A4 lines, will also influence the outcome. In addition to the combination of these factors, translocation of GAs is another factor that makes it difficult to anticipate the effects of scFv production. As was shown in Fig. 3d, GAs in rosette leaves could migrate to the SAM, promote flower induction, and stem elongation. Given that the GAs in rosette leaves are involved in the GA-regulated events in the SAM, the strong phenotypes in A24-lines mean that anti-GA24 scFv captured GA24 and inhibited translocation of GA24 to the SAM, or inhibited the conversion of GA24 to GA9, and then to GA4 in rosette leaves and cut off the supply of GA9 or GA4 to the SAM. Since A4-lines showed only mild phenotypes on flower induction and early stem elongation compared to A24-lines, GA24 or GA9 in rosette leaves of A4-lines were freely migrated to the SAM and converted to GA4 in the SAM, which could not be captured by anti-GA4 scFv efficiently enough to give strong phenotypes because the scFv level is low in the SAM. On the other hand, the stronger effect of anti-GA4 scFv than anti-GA24 scFv on late inflorescence growth might be attributable to a lower endogenous level of GA4 than GA24 in the inflorescence stem. The above-mentioned factors relating to the competitors of scFvs should also be integrated into this discussion. These speculations must be validated by quantitative estimate of scFvs, GA receptors, GA 20-oxidases, and GAs in specific parts of plants related to phenotypes of interest and also by a demonstration of the translocation of endogenous GAs. In spite of these ambiguities, the differential phenotypes by anti-bioactive GAs scFv and anti-precursor GAs scFv expand the capability of immunomodulation.
Abbreviations
- GA:
-
Gibberellin
- scFv:
-
Single chain variable fragment
- ER:
-
Endoplasmic reticulum
- GFP:
-
Green fluorescent protein
- LD:
-
Long day
References
Artsaenko O, Peisker M, Zurnieden U, Fiedler U, Weiler EW, Muntz K, Conrad U (1995) Expression of a single-chain Fv antibody against abscisic-acid creates a wilty phenotype in transgenic tobacco. Plant J 8:745–750
Bechtold N, Ellis J, Pelletier G (1993) In-planta Agrobacterium-mediated gene-transfer by infiltration of adult Arabidopsis thaliana plants. C R Acad Sci Paris Life Sci 316:1194–1199
Conrad U, Manteuffel R (2001) Immunomodulation of phytohormones and functional proteins in plant cells. Trends Plant Sci 6:399–402
De Jaeger G, Buys E, Eeckhout D, De Wilde C, Jacobs A, Kapila J, Angenon G, Van Montagu M, Gerats T, Depicker A (1999) High level accumulation of single-chain variable fragments in the cytosol of transgenic Petunia hybrida. Eur J Biochem 259:426–434
Eriksson S, Böhlenius H, Moritz T, Nilsson O (2006) GA4 is the active gibberellin in the regulation of LEAFY transcription and Arabidopsis floral initiation. Plant Cell 18:2172–2181
Eto J, Suzuki Y, Ohkawa H, Yamaguchi I (2003) Anti-herbicide single-chain antibody expression confers herbicide tolerance in transgenic plants. FEBS Lett 550:179–184
Evan GI, Lewis GK, Ramsay G, Bishop JM (1985) Isolation of monoclonal-antibodies specific for human c-myc proto-oncogene product. Mol Cell Biol 5:3610–3616
Fecker LF, Koenig R, Obermeier C (1997) Nicotiana benthamiana plants expressing beet necrotic yellow vein virus (BNYVV) coat protein-specific scFv are partially protected against the establishment of the virus in the early stages of infection and its pathogenic effects in the late stages of infection. Arch Virol 142:1857–1863
Horsman J, McLean MD, Olea-Popelka FC, Hall JC (2007) Picloram resistance in transgenic tobacco expressing an anti-picloram scFv antibody is due to reduced translocation. J Agric Food Chem 55:106–112
Jobling SA, Jarman C, Teh MM, Holmberg N, Blake C, Verhoeyen ME (2003) Immunomodulation of enzyme function in plants by single-domain antibody fragments. Nature Biotechnol 21:77–80
Lee DJ, Zeevaart JAD (2002) Differential regulation of RNA levels of gibberellin dioxygenases by photoperiod in spinach. Plant Physiol 130:2085–2094
Magome H, Yamaguchi S, Hanada A, Kamiya Y, Oda K (2004) dwarf and delayed-flowering 1, a novel Arabidopsis mutant deficient in gibberellin biosynthesis because of overexpression of a putative AP2 transcription factor. Plant J 37:720–729
Miroshnichenko S, Tripp J, zur Nieden U, Neumann D, Conrad U, Manteuffel R (2005) Immunomodulation of function of small heat shock proteins prevents their assembly into heat stress granules and results in cell death at sublethal temperatures. Plant J 41:269–281
Owen M, Gandecha A, Cockburn B, Whitelam G (1992) Synthesis of a functional antiphytochrome single-chain-Fv protein in transgenic tobacco. Biotechnology 10:790–794
Phillips J, Artsaenko O, Fiedler U, Horstmann C, Mock HP, Muntz K, Conrad U (1997) Seed-specific immunomodulation of abscisic acid activity induces a developmental switch. EMBO J 16:4489–4496
Proebsting WM, Hedden P, Lewis MJ, Croker SJ, Proebsting LN (1992) Gibberellin concentration and transport in genetic lines of pea. Plant Physiol 100:1354–1360
Scheller J, Leps M, Conrad U (2006) Forcing single-chain variable fragment production in tobacco seeds by fusion to elastin-like polypeptides. Plant Biotechnol J 4:243–249
Schouten A, Roosien J, de Boer JM, Wilmink A, Rosso MN, Bosch D, Stiekema WJ, Gommers FJ, Bakker J, Schots A (1997) Improving scFv antibody expression levels in the plant cytosol. FEBS Lett 415:235–241
Shimada N, Suzuki Y, Nakajima M, Conrad U, Murofushi N, Yamaguchi I (1999) Expression of a functional single-chain antibody against GA24/19 in transgenic tobacco. Biosci Biotechnol Biochem 63:779–783
Staehelin LA (1997) The plant ER: a dynamic organelle composed of a large number of discrete functional domains. Plant J 11:1151–1165
Strauss M, Kauder F, Peisker M, Sonnewald U, Conrad U, Heineke D (2001) Expression of an abscisic acid-binding single-chain antibody influences the subcellular distribution of abscisic acid and leads to developmental changes in transgenic potato plants. Planta 213:361–369
Suzuki Y, Mizuno T, Urakami E, Yamaguchi I, Asami T (2008) Immunomodulation of bioactive gibberellin confers gibberellin-deficient phenotypes in plants. Plant Biotechnol J 6:355–367
Sun T-p, Kamiya Y (1994) The Arabidopsis GA1 locus encodes the cyclase ent-kaurene synthetase A of gibberellin biosynthesis. Plant Cell 6:1509–1518
Talon M, Zeevaart JAD (1990) Gibberellins and stem growth as related to photoperiod in Silene armeria L. Plant Physiol 92:1094–1100
Talon M, Koornneef M, Zeevaart JAD (1990) Endogenous gibberellins in Arabidopsis thaliana and possible steps blocked in the biosynthetic pathways of the semidwarf ga4 and ga5 mutants. Proc Natl Acad Sci USA 87:7983–7987
Talon M, Zeevaart JAD, Gage DA (1991) Identification of gibberellins in spinach and effects of light and darkness on their levels. Plant Physiol 97:1521–1526
Tavladoraki P, Benvenuto E, Trinca S, Demartinis D, Cattaneo A, Galeffi P (1993) Transgenic plants expressing a functional single-chain Fv-antibody are specifically protected from virus attack. Nature 366:469–472
ten Hoopen P, Hunger A, Müller A, Hause B, Kramell R, Wasternack C, Rosahl S, Conrad U (2007) Immunomodulation of jasmonate to manipulate the wound response. J Exp Bot 58:2525–2535
Voss A, Niersbach M, Hain R, Hirsch HJ, Liao YC, Kreuzaler F, Fischer R (1995) Reduced virus infectivity in Nicotiana tabacum secreting a TMV-specific full-size antibody. Mol Breed 1:39–50
Wigger J, Phillips J, Peisker M, Hartung W, zur Nieden U, Artsaenko O, Fiedler U, Conrad U (2002) Prevention of stomatal closure by immunomodulation of endogenous abscisic acid and its reversion by abscisic acid treatment: physiological behaviour and morphological features of tobacco stomata. Planta 215:413–423
Wu K, Li L, Gage DA, Zeevaart JAD (1996) Molecular cloning and photoperiod-regulated expression of gibberellin 20-oxidase from the long-day plant spinach. Plant Physiol 110:547–554
Xu Y-L, Gage DA, Zeevaart JAD (1997) Gibberellins and stem growth in Arabidopsis thaliana. Plant Physiol 114:1471–1476
Yang Y-Y, Yamaguchi I, Takeno-Wada K, Suzuki Y, Murofushi N (1995) Metabolism and translocation of gibberellins in seedlings of Pharbitis nil (I). Effect of photoperiod on stem elongation and endogenous gibberellins in cotyledons and their phloem exudates. Plant Cell Physiol 36:221–227
Zimmermann S, Schillberg S, Liao YC, Fisher R (1998) Intracellular expression of TMV-specific single-chain Fv fragments leads to improved virus resistance in Nicotiana tabacum. Mol Breed 4:369–379
Acknowledgments
The authors like to thank Professor Yasuo Niwa, University of Shizuoka, for providing a CaMV35S-sGFP(S65T)-NOS3′ plasmid. This research was partly supported by the Program for Promotion of Basic Research Activities for Innovative Biosciences (PROBRAIN).
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Urakami, E., Yamaguchi, I., Asami, T. et al. Immunomodulation of gibberellin biosynthesis using an anti-precursor gibberellin antibody confers gibberellin-deficient phenotypes. Planta 228, 863–873 (2008). https://doi.org/10.1007/s00425-008-0788-z
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00425-008-0788-z