
Abstract
Arabidopsis thaliana possesses three RpoT genes which encode three different phage-type RNA polymerases with yet unknown function in organelle transcription: RpoTm and RpoTp, imported into mitochondria and plastids, respectively, and RpoTmp, co-targeted into both organelles. Expression of the RpoT genes was analyzed by quantitative RT-PCR, histochemical β-glucuronidase (GUS) assays and in situ hybridization. Transcripts of all three RpoT genes accumulated to very low amounts in all organs. Surprisingly, RT-PCR revealed their highest levels in flower tissues. RpoTm transcripts were the most abundant in all organs, except mature leaves, in which RpoTp transcripts showed the highest accumulation. In the developing seedling, RpoTm::GUS and RpoTmp::GUS expression precedes that of RpoTp::GUS, the latter showing up only 7 days after germination. The RpoTm and RpoTmp promoters expressed GUS mainly in meristematic and mitochondria-rich cells such as the distal part of the root and companion cells flanking the phloem, whereas RpoTp::GUS activity was found in green tissues as the parenchyme cells of young leaves, the primary cortex of the stem, and sepals of buds and young flowers. Sites of GUS expression coincided spatially with those of in situ hybridization. Our data demonstrate an overlapping expression pattern of RpoTm and RpoTmp, and a completely differing pattern of RpoTp expression. The results suggest that RpoTm and RpoTmp recognize different types of mitochondrial promoters. The plastid polymerase RpoTp might play a major role in green tissue, i.e. in chloroplast transcription, whilst the dual-targeted RpoTmp in plastids should function mainly in the transcription of genes in non-green types.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Mitochondria and plastids, acquired through endosymbiosis from ancestral free-living cyanobacteria and α-proteobacteria, respectively, possess their own genomes and transcription systems. The RNA polymerases (RNAPs) of the two extranuclear organelles fall into two classes: one eubacterial-type multi-subunit enzyme encoded by and active in plastids, and a single-subunit core RNAP, related to RNAPs of bacteriophages T3 and T7, found in both plant organelles (Weihe 2004). Thus, plastids harbor at least two different types of RNAPs: the plastid-encoded eubacterial-type plastid polymerase (PEP), composed of four subunits (α, β, β′, β′′) and one of several nuclear-encoded sigma factors. A second plastid RNAP which is nuclear-encoded (NEP) is represented by one (monocots) or two (dicots) phage-type RNAPs (for reviews see Hess and Börner 1999; Weihe 2004). Isolation of the RpoT genes from Chenopodium album (Weihe et al. 1997), Arabidopsis thaliana (Hedtke et al. 1997, 1999, 2000), maize (Young et al. 1998; Chang et al. 1999), wheat (Ikeda and Gray 1999), Nicotiana tabacum (Hedtke et al. 2002), Nicotiana sylvestris (Kobayashi et al. 2001b, 2002; Hedtke et al. 2002), barley (Emanuel et al. 2004), and rice (Kusumi et al. 2004) and demonstration of subcellular localization of their products has led to the identification of the plant phage-type mitochondrial and plastid RNA polymerases (RpoTm and RpoTp, respectively). One RpoT gene (RpoTmp) found in Arabidopsis and Nicotiana, but not in monocots, has been proposed to encode a protein most probably targeted into both mitochondria and plastids (Hedtke et al. 2000, 2002; Kobayashi et al. 2001a;). Experimental evidence that RpoTp is a catalytic subunit of plastid NEP comes from the analysis of plastid transcription in tobacco and Arabidopsis plants overexpressing N. sylvestris and A. thaliana RpoTp (Liere et al. 2004). Thus far, it is not understood why transcription of dicot organelle genomes, both in mitochondria and plastids, requires two phage-type polymerases, while in monocots only one RpoT polymerase is engaged in the transcription of the mitochondrial and plastid genome, respectively. It is also unknown whether the two phage-type RNAPs in each organelle (RpoTm and RpoTmp in mitochondria, RpoTp and RpoTmp in plastids) fulfill distinct roles in organelle transcription, e.g. have different preferences for certain promoters or sequence motifs. Only recently, analysis of an Arabidopsis activation-tagged T-DNA insertion line has provided evidence for a specific role of RpoTmp at least in plastid gene expression. The mutation of the RpoTmp gene affected the light-induced accumulation of several plastid mRNAs and proteins and resulted in a lower photosynthetic efficiency. In contrast to these alterations in plastid gene expression, no major effect of the RpoTmp mutation on the accumulation of examined mitochondrial gene transcripts and proteins was observed (Baba et al. 2004). Overexpression of RpoTp in tobacco has provided evidence for distinct promoter specifity of RpoTp (Liere et al. 2004). No data have been reported yet supporting distinct roles of the two phage-type RNAPs, RpoTm and RpoTmp, in plastids and mitochondria.
While systematic data on the expression of the RpoT genes during plant development and the specific role the individual polymerases play in developmentally regulated and tissue-specific transcription of the organelles is lacking, a study of the expression of the two RpoT polymerases in a barley leaf gradient has shown that the developmental stage of the plastid in the leaves has a strong impact on the expression and activity of the organelle phage-type RNAPs and their target genes (Emanuel et al. 2004). In a recent study, it was shown that the NEP (RpoTp) enzyme level in maize leaves decreases as plastids mature, while production of mRNAs by NEP increases (Cahoon et al. 2004). Interorganellar cross-talk in the regulated expression of the RpoT genes might play an important role in the concerted action of genes located within plastids, mitochondria and the nucleus of the plant cell. For dicotyledonous plants, as Arabidopsis, which harbor a second phage-type RNAP (RpoTmp) in both mitochondria and plastids, no such data are available. To address these questions, we have initiated an investigation of the cell- and tissue-specific expression of the RpoT genes in Arabidopsis development. The expression of the AtRpoTm, AtRpoTp and AtRpoTmp genes was studied by quantitative real-time RT-PCR, histochemical β-glucuronidase analysis and in situ hybridization. Here we report remarkable changes in the expression and transcript accumulation of all three RpoT genes indicating both distinct roles of the encoded RNA polymerases in organelle transcription and developmental control of their activity.
Materials and methods
Plant material and growth conditions
Arabidopsis thaliana (ecotype Columbia) seeds were grown, after a 2-day 4°C vernalization treatment, on MS medium at 20°C under a 8h/16h light/dark cycle. After 10 days the seedlings were transferred to soil and after 14 days the seedlings were exposed under a light regime of 16h/8h light/dark. The intensity of light in all experiments was 210 μmol photons s−1 m−2.
Isolation of RNA and quantitative real-time RT-PCR
Total cellular RNA was extracted from 100 mg plant tissue using TRIZOL (Invitrogen). After a wash step with 70% EtOH, the RNA pellet was resuspended in 50 μl H2O, column-purified and DNase-treated using the NucleoSpin RNA plant kit (Macherey-Nagel, Düren, Germany). The integrity of the eluted and quantified RNA was verified by agarose gel electrophoresis. About 2 μg RNA were reverse transcribed using Omniscript reverse transcriptase (QIAGEN). RNA was heated to 65°C for 5 min, followed by addition of reaction buffer, 1 mM of each dNTP, 50 pmol of random hexamer primers, 20 U RNase inhibitor (Fermentas, St. Leon-Rot, Germany), 4 U reverse transcriptase and water to a final volume of 20 μl. Reactions were incubated at 37°C for 1 h. The enzyme was inactivated at 93°C for 5 min. Quantitative PCR was carried out in an Applied Biosystems GeneAmp 5700 sequence detection system using SYBR Green PCR core reagents (Applied Biosystems). Each 25 μl reaction contained 1× buffer, 2 mM of each dNTP, 3 mM MgCl2, 0.25 U AmpEraseUng, 0.625 U Amplitaq Gold DNA-polymerase, 45 pmol of each primer and different amounts of cDNA as template, corresponding to 100 ng (RpoTm, RpoTmp, RpoTp, rpoB, GAPDH) and 10 pg (18S-rRNA), of reverse transcribed total RNA.
Primer pairs for quantitative real-time RT-PCR were chosen to yield amplification products between 90 and 110 bp. The reactions were incubated for 2 min at 50°C, then for 10 min at 95°C, followed by 40 cycles of 15 s at 95°C, 30 s at 60°C and 45 s at 72°C. After the last cycle, the PCR products were subjected to heat denaturation over a temperature gradient from 60 to 95°C at 0.03°C/s to check for the absence of non-specific amplification products. To account for variability in the initial RNA concentrations, all quantitations were normalized to the amount of cytoplasmic 18S-rRNA or GAPDH as internal standards by the ΔC T method ( \(2^{{ - \Delta C_{{\text{T}}} }} \;{\text{[}}\Delta C_{{\text{T}}} = C_{{\text{T}}} \) target – C T endogenous reference]). Two different preparations of cDNA were analyzed for each gene to control the quality of total RNA and the conversion efficiency of the reverse transcription reaction. All quantitative real-time RT-PCRs were performed in duplicates using two different dilutions from two independent sets of cDNA syntheses. This was repeated on two sets of independent RNA preparations each, yielding 16 individual values for each RNA sample altogether. Non-template control reactions were performed for each pair of primers. Negative controls (omitting reverse transcriptase) were included on all samples to verify the complete removal of DNA. The data were analyzed using PE GeneAmp 5700 SDS software version 1.3 (Applied Biosystems) and plotted as normalized reporter signal (Rn). Rn represents the level of fluorescence detected during the PCR process after subtraction of background noise versus cycle number. A threshold was set manually to 0.8 in the upper third of the linear phase of the amplification curve. The C T value (threshold cycle) indicated the cycle number when the amount of amplified targets, i.e. the reporter fluorescence, reached the threshold.
Construction of the RpoT-promoter-β-glucuronidase gene fusions
Genomic fragments containing 930 or 1,983 bp (RpoTm), 756 or 2,238 bp (RpoTmp) and 889, 2,025 or 3,420 bp (RpoTp) of the sequence upstream of the ATG start codon were amplified by polymerase chain reaction. DNA fragments were verified by sequencing and introduced into the promoterless Agrobacterium binary plasmid pGPTV-bar (Becker et al. 1992) via HindIII/XbaI restriction sites. The RpoT-pGPTV-bar constructs were transferred into Agrobacterium strain GV3101::pMP90 (C58).
Plant transformation
Arabidopsis wild type plants (ecotype Columbia, Col-O) were transformed by means of the floral dipping method (Clough and Bent 1998). Transformants were selected on Murashige and Skoog (MS) medium containing 30 mg/l phosphinotricin, 5% sucrose and 0.8% agar. The genomic integration of introduced RpoT-promotor-gus fusions in T1 transgenic plants was verified by genomic PCR and Southern blot hybridization. For each construct at least 12 independent transgenic plants were assayed for β-glucuronidase (GUS) activity at different developmental stages and in various tissue types.
Histochemical GUS assay
β-glucuronidase analyses were done on T1 plants (obtained by self-fertilization of the primary transformants) and verified in the T2 and T3 generations. Plant tissue samples were stained under vacuum with X-Gluc solution containing 1 mg/ml X-Gluc (5-bromo-4-chloro-3-indolyl-β-glucuronide, Fermentas), 50 mM sodium phosphate pH 7.0, 10 mM EDTA, 0.1% Triton X-100, 2 mM potassium ferrocyanide and 100 mg/ml chloramphenicol (Jefferson et al. 1987a, b). All staining reactions were incubated at 37°C for 12 h. GUS activity was observed in transversal stem sections, prepared from embedded or fresh material. Stained stems were cut into approximately 1 cm long segments and fixed overnight at 4°C in 50 mM sodium phosphate (pH 7.0) containing 2% glutaraldehyde. After being rinsed in 50 mM sodium phosphate (pH 7.0, 4°C) for 15 min, the tissue was dehydrated in a graded ethanol series of 10, 20, 40, 60, 80, 90, 95, and 100% (v/v, 4°C) each for 10 min and 100% ethanol twice for 30 min. The tissue was infiltrated with pure London Resin White (London Resin Co. Ltd., London, UK) for 1 h, followed by one change of LR White for 1 h and one change overnight. The stem segments were placed into Gelantine Capsules (Plano) and polymerized at 60°C for 24 h. Transverse sections (14 μm) were cut with a microtome (Microm HM 355) using 45° glass knives, mounted on slides, and covered with Roti®-Histokit (Roth GmbH, Karlsruhe, Germany). Sections were examined in a Zeiss microscope (Axioskop 2 plus) by bright-field microscopy and photographed using the ColorView Soft Imaging System. For hand-cut sections from fresh material, stem segments were first dehydrated as described above and then rehydrated. The segments were fixed in 3% agarose and cut with ethanol-washed razor blades. The cross sections were observed by dark-field microscopy.
In situ hybridization
In situ hybridization of RpoT transcripts was performed with Arabidopsis flowers and stem sections. Digoxigenin-labeled sense and antisense RNA probes were synthesized from RpoT cDNA using T7 polymerases according to the instructions of the supplier (DIG RNA labeling Kit, Roche, Mannheim, Germany).
Plant tissues were fixed, dehydrated, and embedded in paraffin essentially as described by Drews et al. (1991), (http://www.its.caltech.edu/~plantlab/protocols/insitu.html). Sections of Arabidopsis flowers, 6–8 μm thick, were cut and mounted on poly-l-lysine coated slides (Poly-PrepTM Slides, Sigma) and processed essentially as described by Drews et al. (1991).
Tissue sections were covered with 100 ng of denatured RNA probe in 60 μl hybridization solution (50% formamide, 0.3 M NaCl, 10 mM Tris/HCl pH 7.5, 1 mM EDTA, 1× Denhardt´s solution, 10% dextran sulphate, 1 M DDT, 1.5 mg/ml yeast tRNA). Hybridization was done in a humid chamber for 16 h at 53°C. Washing steps were performed in 2× SSC, for 30 min, in 0.2× SSC for 30 min at 56°C and for 10 min in 0.2× SSC at room temperature. Immunological detection using antibodies against digoxigenin coupled with alkaline phosphatase and final staining procedure with NBT/BCIP was done according to the supplier’s manual (Roche).
Results
Transcript accumulation using real-time RT-PCR
Previous analyses of mRNA accumulation by RNA gel blot hybridization showed an extremely low abundance of RpoT transcripts in mature leaves of Chenopodium album and A. thaliana (Hedtke et al. 1997; Weihe et al. 1997). Therefore, to study the accumulation of mRNA in different tissues and developmental stages, we exploited the more sensitive and accurate method of quantitative real-time RT-PCR. Tissue samples were derived from Arabidopsis plants grown in soil over a period of 9 weeks. RNA was isolated from flowers, stems, rosette leaves, and roots. Total adult flowers were used, considering developed sepals and differentiated anthers as criteria for adult organs.
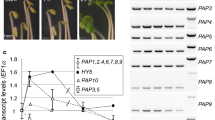
Comparison of different parts of the adult plant revealed a remarkable high transcript accumulation in total flower tissue relative to leaves, stems and roots (Fig. 1a). The steady state level of all three RpoT transcripts was 2.8–9 fold higher in total flower tissue compared to leaves, stems and roots. The ratio between the three RpoT mRNAs was found to be similar in all tissues investigated except leaf tissue. RpoTm showed the highest abundance in flowers, stems and roots, whereas the RpoTp mRNA was the most abundant in leaves, accumulating to levels more than twice of RpoTm and more than three times of RpoTmp.

a, b Analysis of RpoT transcript accumulation by quantitative real-time RT-PCR. a RpoT genes were assayed for transcript abundance in flower, stem, leaf and root tissue. Data were normalized to the amount of cytosolic 18S-rRNA and expressed as 2-dCT (for details, see “Material and Methods”). b Comparison of cotyledons (7-days-old) and leaf tissue harvested at 42 days of plant growth. The bars represent relative transcript abundance and were calculated with GAPDH as endogenous reference. 1w one week, 6w six weeks
In order to investigate developmental changes of the RpoT transcript accumulation, mRNAs from leaves of different ages were quantified. We compared cotyledons of 7-days-old seedlings and leaves from 42-days-old plants showing fully developed leaf rosettes. Quantification of the steady state levels of RpoT transcripts revealed a changing pattern of transcript accumulation of the RpoT genes during the early phase of leaf development (Fig. 1b): In cotyledons of 7-days-old plants, RpoTm mRNA was the most abundant of the three polymerases genes, accumulating 1.7 fold relative to RpoTp and 1.8 fold relative to RpoTmp. During further development, the total level of RpoTm transcripts in the leaves declined to values of approximately 30% after 42 days of plant growth (Fig. 1b). Similarly, RpoTmp transcript accumulation was also higher in cotyledons and decreased to 37% in leaves of 42-days-old plants. In contrast to RpoTm and RpoTmp transcripts, RpoTp mRNA in the mature leaves accumulated to approximately the same level as in the cotyledons. Thus, RpoTp is the most abundant of the three RpoT mRNAs in mature leaves.
Tissue-specific RpoT expression during development as revealed by GUS-staining
In order to study the expression pattern of the RpoT genes during development of different tissues, 12 individual RpoT::GUS plants of each transgenic line were analyzed at 2, 4, 7 and 14 days after germination. In 2-days-old seedlings, GUS staining was first observed for RpoTm in the distal part of the root, i.e. the elongation and differentiation zone (Fig. 2, a1). No activity at this stage was seen for RpoTmp and RpoTp (Fig. 2, a2, a3). After 4 days, when the seedlings began to form primary leaves, the strongest GUS staining was observed with the RpoTm::GUS lines, showing up in apical meristem, in the root tips and in the vascular tissue of the cotyledons (Fig. 2, b1). RpoTmp::GUS plants were stained in the root tips (Fig. 2, b2), while GUS activity in RpoTp::GUS plants was still lacking (Fig. 2, b3). After 7 days, the delayed RpoTmp activity showed up, in an identical pattern to RpoTm::GUS plants, in the vascular tissue of the cotyledons, in the root and, at the highest level, in the primary leaves (Fig. 2, c1, c2). In the division zone of the root, RpoTm::GUS and RpoTmp::GUS expression was observed throughout further development of the plants. The decline of cell division activity was accompanied by a fading of the GUS staining towards the proximal parts of the root tips (see Fig. 2, d1, d2). Budding lateral roots were heavily stained in RpoTm::GUS and RpoTmp::GUS plants (Fig. 2, e1, e2). At day 7, the first, very weak staining of RpoTp::GUS appeared in the primary leaves (Fig. 2, c3). This RpoTp activity, confined to the leaves, became prominent only after 14 days (Fig. 2, f3). In contrast to RpoTm and RpoTmp (Fig. 2, f1, f2), the RpoTp::GUS activity occurred mainly in the non-vascular, photosynthetically active parenchyme tissue of the primary leaf (Fig. 2, f3). The cotyledons of the RpoTp::GUS plants showed no detectable staining during the whole development. The intensity of staining in RpoTm::GUS and RpoTmp::GUS plants in the primary leaf tissue decreased during further development and shifted towards the vascular system. Young primary and young secondary leaves at 14 days were intensely stained, while in the older primary leaves GUS activity was restricted to the vascular system (Fig. 2, f1, f2). Microscopic examination revealed that the GUS activity occurred in the phloem (i.e. the companion cell layer), rather than in the xylem (Fig. 3e).

a–f β-Glucuronidase staining of seedlings of A. thaliana transformed with RpoT-promoter::GUS fusion constructs. Histochemical localization of RpoTm-(a1–f1), RpoTmp-(a2–f2), RpoTp-(a3–f3) promoter::GUS activity was analyzed at different developmental stages. Seedlings were analyzed at ages of: 2 days (a1–3), 4 days (b1–3), 7 days, cotyledons (c1–3), 7 days (root tips, d1–3), 7 days (developing lateral roots, e1–3), and 14 days after germination (f1–3)

a–f β-Glucuronidase staining of vegetative organs of A. thaliana transformed with RpoT-promoter::GUS fusion constructs: RpoTm::GUS (a1–c1), RpoTmp::GUS (a2–c2, d–f), RpoTp::GUS (a3–c3). a1–3 developing stipules on a primary sprout, arrows depict meristematic tissue of stipules, the insert a2b shows a magnification of a stipule. b1–3 cross sections of stems (paraffin embedded, 10 μm-thick slices), c1-3 cross sections of primary sprouts (agarose embedded, ca. 100 μm-thick slices), d developing trichomes of a primary leaf pair, e vascular bundle tissue of a 7 days old cotyledon, f vascular bundle tissue of an adult leaf. c Cotyledon, p phloem, x xylem, arrows stipule
In all older plants (>4 weeks), containing the RpoTm::GUS and RpoTmp::GUS constructs, GUS activity decreased further in mesophyll cells. GUS was preferentially expressed, beside the vascular bundle tissue, in newly formed organs or tissues (Fig. 3, a1, a2, d, e, f). In Arabidopsis, epidermal cells of the fully differentiated stem are able to develop into new organs like stipules. These re-embryonalized meristematic cells, developing into stipules, showed a high GUS activity in RpoTm and RpoTmp plants during the initial phase of development, while adult stipules were only faintly stained (Fig. 3, a1, a2). Microscopic examination revealed a clear difference between pronounced staining of the meristematic cells of the stipule and the unstained cells of the stem (Fig. 3, a2b). In contrast, RpoTp::GUS activity was not observed in meristematic tissues of the stem, but showed up in young stipules in a pattern similar to that observed in young leaves of the seedling and the rosette of older plants (Fig. 3, a3).
A change of RpoTm- and RpoTmp-driven GUS activity was also detected in developing leaf trichomes consisting of single cells. As an example, Fig. 3d shows a remarkably high GUS expression during the early stage of development of trichomes. In fully expanded trichomes, no GUS activity was detectable (Fig. 3d).
The stems of the RpoT::GUS plants showed varying staining intensities during different phases of development (Figs. 3, b1-3, c1-3, 4 a1-3). The high GUS expression observed in RpoTm::GUS and RpoTmp::GUS plants in the middle part of stems of young florescences was fading during later developmental stages. Thus, GUS staining was only faint in older stems of adult flowers (Fig. 4, a1-2). Microscopic examination of transversal sections in young stems revealed that staining occurred in the phloem, predominantly in the companion cells, in the epidermal cell layer and the primary cortex (shown for an RpoTmp::GUS plant, Fig. 3, b2). In stem sections of older florescences and siliques, which externally showed no GUS staining, RpoTm and RpoTmp::GUS activity was detected in microscopic sections, exclusively in the vascular bundle tissue (shown for an RpoTm::GUS plant, Fig. 3, b1). For analysis of later stages of stem development, transversal sections of the primary sprout were examined. These sections showed RpoTm and RpoTmp::GUS activity in the phloem (Fig. 3, c1, c2). In plants which contained at least four copies of RpoTm::GUS and RpoTmp::GUS, stomata in the epidermis of stems were stained as well (shown for RpoTm::GUS plants in Fig. 3, b1).

a–h β-Glucuronidase staining of floral organs of A. thaliana transformed with RpoT-promoter::GUS fusion constructs. Comparison of inflorescenses of RpoTm::GUS (a1), RpoTmp::GUS (a2) and RpoTp::GUS plants (a3). RpoTm::GUS (b, c) and RpoTmp::GUS (d–g) plants showing similar expression patterns in flower organs: b stigmatic papillae of a bud and a whole stained carpel of an adult flower, c pollen tube (siphonogamy), d tapetum of an anther of a bud, e carpel of an adult flower, f carpel of an adult flower (paraffin embedded, 15 μm-thick slice), g abscission layer of a withered flower. RpoTp::GUS plants exhibiting high GUS activity in stems (a3), sepals of a bud (h), sepals of a bud, paraffin embedded, 15 μm-thick slice (i)
A completely different pattern of GUS expression was observed in stems of RpoTp::GUS plants. Young stems, underneath the primordia and buds, showed a much weaker expression than older stems from adult flowers and siliques (Fig. 4, a3). In transversal sections, GUS staining could be attributed to the primary cortex (Fig. 3, b3, c3).
GUS staining was as well detected in flowers (Fig. 4). RpoT::GUS expression was found to be high in flower tissues, with clear differences between RpoTm and RpoTmp on the one hand, and RpoTp on the other. RpoTm and RpoTmp promoter-dependent GUS activity exhibited a similar spatio-temporal pattern, with the highest activity showing up in the primordia of buds (Fig. 4, a1, a2). With the formation of the flower organs, the intensity of GUS staining in buds was fading, and promoter activity occurred preferentially in the vascular system of the stamen, petals and sepals. Anthers, in the case of RpoTm and RpoTmp, showed GUS staining in the tapetum exclusively in developing flower buds (Fig. 4d). A strongly developmentally dependent expression of GUS in RpoTm::GUS and RpoTmp::GUS plants was observed in the carpels. In buds, high GUS activity was confined to the stigmatic papillae (Fig. 4b). After pollination (siphonogamy), the pollen tubes became intensely stained (Fig. 4c), and finally, during the later phase of flower development, GUS activity was spread over the whole carpel (see Fig. 4b, e, f). The carpel of adult flowers and siliques exhibited RpoTm::GUS and RpoTmp::GUS activity in the vascular tissue feeding the embryos (Fig. 4e, f). Furthermore, a strong GUS activity in RpoTm and RpoTmp plants was observed in the abscission zone of aging florescences (Fig. 4g). The pattern of RpoTp::GUS expression in florescences differed substantially from that of RpoTm and RpoTmp. RpoTp::GUS staining was observed exclusively in green tissue, i.e. sepals, stipules and stems (Fig. 4, a3, h, i). Sepals of buds were stained most intensely, and in fully expanded flowers staining decreased (Fig. 4, a3).
Table 1 summarizes the spatio-temporal distribution of RpoT promoter-driven GUS activity during the development of transgenic Arabidopsis plants. Semiquantitative indication of staining intensities reveals the principal similar expression pattern between RpoTm and RpoTmp on the one hand, and the completely different staing in the case of RpoTp::GUS plants on the other.
In situ hybridization studies
In situ hybridization was employed to assay tissue-specific accumulation of endogenous RpoT mRNA. Tissue sections were hybridized with antisense and sense strand digoxigenin-labeled RNA probes derived from the 5′ end of the coding region (exon 1) of the RpoT genes. Exon 1 exhibits no significant DNA sequence similarity between the three RpoT genes. Figure 5 shows representative results of these experiments. In general, the signals obtained were relatively weak, essentially detectable only in flower tissue and transversal sections of the stem.

a–g In-situ hybridization of RpoT genes in developing flowers of A. thaliana. DIG-labeled antisense and sense RNA probes of RpoTm (a1–d1), RpoTmp (a2–d2, e, f, g) and RpoTp (a3–d3) were hybridized to longitudinal sections of flowers at different developmental stages, and to stems. a1–3 Umbel, antisense RNA probe, b1–3 umbel, sense probe, c1–3 cross section of flowers, antisense probe, d1–3 Transversal slices of stems, antisense probe, e anthere with tapetum, RpoTmp antisense probe, f detail of a cross section of a stem showing vascular bundle tissue, RpoTmp antisense probe, g abscission layer of a withered flower, RpoTmp antisense probe. X Xylem, p phloem
Very early stages of flower development showed high in situ hybridization signals of all three RpoT genes in the primordia of buds (Fig. 5, a1-3). In the first visible flower structures, the sepal primordia, relatively high hybridization signals of all three RpoT probes were obtained (not shown). In later phases of development, hybridization in the green tissue, i.e. the sepals, showed up only in the case of RpoTp, whereas RpoTm and RpoTmp transcripts were localized to the vascular bundle tissue, confirming the GUS staining data (Fig. 5, a1-3, c1-3). During early phases of development a relatively high transcript accumulation of the RpoT genes was also observed in the primordia of the carpel and the anthers (Fig. 5, a1-3). RpoTp hybridization decreased as buds developed into adult flowers and was no longer detectable in flowering florescences, while RpoTm and RpoTmp transcripts showed up during the whole flower development (Fig. 5, c1-3). RpoTm and RpoTmp transcript accumulation in anthers was clearly dependent upon the developmental stage: whereas the primordia of anthers showed strong hybridization signals spread over the whole tissue (Fig. 5, a1, a2), transcript accumulation in buds was confined to the tapetum as shown for RpoTmp in Fig. 5e. Thereafter, transcript levels declined in the degenerating tapetum and occurred exclusively in the vascular tissue of stems of anthers (Fig. 5, c1, c2). RpoTm and RpoTmp transcripts were detected in the vascular system of sepals and petals, as well as the stems of anthers (Fig. 5, c1, c2). In contrast, the RpoTp probe showed a diffuse distribution over the sepal tissue, with no preference for the vascular tissues (Fig. 5, c3). In petals, no hybridization signals were detected with RpoTp as probe. In later stages of development, when the flower starts to wither, expression of RpoTm and RpoTmp occurred in the abscission zone (Fig. 5g), corresponding with the data obtained by the GUS analyses.
Striking differences in the differential transcript accumulation of the three RpoT genes, which once more confirmed the promoter GUS activity studies, were observed in stem tissue sections. In transversal sections, RpoTm and RpoTmp transcripts were detected exclusively in the vascular tissue. The signals could be localized to the phloem-surrounding companion cells (Fig. 5, d1, d2, f), whereas RpoTp hybridization signals were found exclusively in the primary cortex (Fig. 5, d3).
Discussion
The tissue- and organ-specific expression of the RpoT genes encoding mitochondrial and plastid RNA polymerases during the development of Arabidopsis plants was examined through detection of mRNA steady-state levels by in situ hybridization, by their quantification using real-time RT-PCR, and by introducing promoter::GUS constructs into A. thaliana and analyzing the GUS activity in several independent transformants. In general, we observed striking differences between the spatio-temporal expression of RpoTm and RpoTmp on the one hand, and RpoTp on the other. Our results indicate that the expression of all RpoT genes is under developmental control. During the first days of plant development, a pronounced RpoTm- and RpoTm-mediated GUS activity in the extension and division zone of the root tips was observed. Active mitochondrial biogenesis has been shown to occur in root apical meristem of Arabidopsis and other species (Kuroiwa et al. 1992; Fujie et al. 1993). The analysis of expression of mitochondrial transcripts (Li et al. 1996) and nuclear transcripts encoding mitochondrial proteins (Ribichich et al. 2001; Elorza et al. 2004) showed that transcript levels correlated with cell division activity in roots, which is in agreement with high GUS activity driven by the promoters of the mitochondrial RNA polymerases, RpoTm and RpoTmp, found in the meristematic zone of Arabidopsis root tips. The developing seedlings showed a high RpoTm and RpoTmp promoter-mediated GUS activity also in the shoot apical system, vascular bundle tissue, developing trichomes and lateral root tips. The only difference was that GUS staining was lower for RpoTmp, compared to RpoTm, during the first days of plant development. This is in agreement with the data of RpoT mRNA steady state levels, obtained by quantitative real-time RT-PCR. With the completion of the organs, transcript accumulation of RpoTm and RpoTmp generally declined. A developmental regulation of mitochondrial gene expression was also reported for primary leaves of wheat, in which an about fivefold to tenfold decrease of the copy number of mitochondrial genes per cell from the meristematic basal section upwards to the mature part of the leaf was observed. The abundance of the corresponding transcripts was found to decrease gene-specifically between 2.5- and 20-fold (Topping and Leaver 1990; Emanuel et al. 2004).
Adult transgenic RpoTm::GUS and RpoTmp::GUS plants showed a strong GUS staining in the vascular tissue, specifically in the companion cells flanking the phloem. Companion cells contain a high number of mitochondria, reflecting the high energetic requirement in these cells. RpoTm encodes a mitochondrial RNAP, whereas RpoTmp specifies a polymerase dually targeted to both mitochondria and plastids. Our results are in favour of an essential role of mitochondria during anther development, as revealed by studies of cytoplasmatic male sterility (for review, see Touzet and Budar 2004). Active biogenesis of mitochondria has been shown to occur in meiotic and tapetal cells during early anther development in maize, where numbers of mitochondria increase 20- to 40-fold (Lee and Warmke 1979). The observed high expression of RpoTm- and RpoTmp-mediated GUS activity in inflorescences, as compared to mature leaves, is consistent with the high number of mitochondria in flower tissues (Huang et al. 1994). In later stages of development, GUS activity was confined to the vascular bundle tissue and the abcission zone.
Taken together, our data indicate nearly identical patterns of expression for RpoTm and RpoTmp. Transcript accumulation as well as GUS activity (see Table 1 for a summary) were prominent mostly in meristemactic and young tissue with high mitochondrial acitivity. The only difference observed was that RpoTm-driven GUS activity appeared earlier than that of RpoTmp, what might be due to the limits of detection of the applied methods, since transcript accumulation detected by real-time RT-PCR was higher for RpoTm than for RpoTmp in all tissues.
In striking contrast, GUS expression under the control of the RpoTp promoter was only detected in green, photosynthetically active tissues of leaves, stems and flowers. Examination of the putative RpoT promoter sequences (2,000 nucleotides upstream the start codon) in silico by scanning against a database of plant cis-acting regulatory DNA elements (http://www.dna.affrc.gojp/PLACE/) did not reveal any general similarity between the three genes. In particular, RpoTm and RpoTmp sequences were not more closely related to each other than to RpoTp (results not shown). To relate the striking similarity of RpoTm and RpoTmp::GUS and the completely different RpoTp::GUS expression to individual DNA sequences or motifs, detailed experimental studies of the promoter sequences will be necessary.
Results almost identical to those of the GUS assays were obtained by in situ hybridization. From comparison with the RT-PCR data it is evident that only those tissues could be stained for GUS activity and by in situ hybridization, that exhibited the highest expression levels. Roots, for example, clearly accumulated RpoTp transcripts (as detected by RT-PCR), but neither GUS staining nor in situ hybridization gave positive results. In general, similar expression patterns are revealed when comparing the GUS expression as described in the present study, and comprehensive microarray data resumed at the Arabidopsis microarray database (http://www.genevestigator.ethz.ch). In a recent study, mRNA levels in different root tissues were analyzed using expression profiling on Arabidopsis microarrays (Birnbaum and Benfey 2004), where transcript accumulation of the RpoT genes showed a similar pattern as obtained in our study by real-time RT-PCR. On average (over different cell layers of the root tips) RpoTm exhibited the highest accumulation, and RpoTmp the lowest. As in our RT-PCR studies, RpoTp expression was well detected in the root cells. The discrepancy between data obtained by different methods might be explained by the fact that RpoTp transcripts accumulate to moderate levels in all or most root cells, summing up to an amount easily detectable by RT-PCR and expression profiling using microarrays, but not seen by detecting GUS activity. Different transcript stability as a result of posttranscriptional regulation as well as translational control might contribute to certain inconsistencies in the results, since GUS expression reflects transcriptional activity of the RpoT promoters as well as the stability of the GUS mRNA and protein, whilst the other methods used in this study measure RpoT transcript accumulation. Since we have obtained identical results independent of promoter length (for constructs used, see Materials and Methods), the shortest version of the respective promoter should contain all the cis elements needed for tissue- and developmental state-specific expression as observed during this study. The expression of GUS activity in the transgenic plants correlated well with the steady-state level of the specific mRNAs as revealed by real-time RT-PCR and in situ hybridization. Therefore, we are confident that the GUS activity observed reflects the expression pattern of the RpoT genes. To rule out an influence of GUS-mRNA stability and GUS enzyme activity on these data, immunodetection of the RpoT polymerases using specific antibodies would be necessary.
According to data with constructs consisting of the RpoTmp transit sequence fused to GFP, RpoTmp is dually targeted to both mitochondria and plastids in Arabidopsis and Nicotiana (Hedtke et al. 2000, 2002; Kobayashi et al. 2001a). Thus, in mitochondria two phage-type polymerases are supposed to be involved in transcription, RpoTm and RpoTmp. This has recently been questioned by the observation that constructs containing the RpoT-5′UTR in addition to the putative transit sequence seem to target RpoTmp only to mitochondria, at least in leaves (Kabeya and Sato 2005). The completely overlapping expression patterns of the RpoTm and RpoTmp would suggest (though in the absence of data on the respective protein levels this remains speculation) that the two polymerases have similar levels of activity at the same developmental stages and in all tissues. Our data, supporting a function of RpoTmp in mitochondria rather than in chloroplasts, are in conflict with a recent study of an RpoTmp null mutant, in which severe effects on plastid, but not on mitochondrial transcription were demonstrated (Baba et al. 2004). If RpoTmp indeed has an impact onto plastid gene expression, then our data would suggest that the two nuclear-encoded plastid polymerases, RpoTmp and RpoTp [representing together the NEP activity, (Liere et al. 2004)] play their specific roles in the plastids of different tissues: RpoTmp preferentially in non-green, and RpoTp in photosynthetically active green tissues. Pronounced expression of RpoTp occurred in green tissue of young leaves and cell types rich in chloroplasts (parenchyme cells). It should be emphasized in this context that RpoTp transcripts are also present in non-green roots, and RpoTmp transcripts were found in green leaves. Yet, the RpoTp gene showed a delayed expression, coinciding with the development of photosynthetic competence, suggesting that RpoTp, like the plastid-encoded PEP, has to play its major role in photosynthetically active cells.
Abbreviations
- GUS:
-
β-Glucuronidase
- NEP:
-
Nuclear-encoded plastid RNA polymerase
- PEP:
-
Plastid-encoded plastid RNA polymerase
- RpoT:
-
Phage-type RNA polymerase
- RNAP:
-
RNA polymerase
References
Baba K, Schmidt J, Espinosa-Ruiz A, Villarejo A, Shiina T, Gardestrom P, Sane AP, Bhalerao RP (2004) Organellar gene transcription and early seedling development are affected in the rpoT;2 mutant of Arabidopsis. Plant J 38:38–48
Becker D, Kemper E, Schell J, Masterson R (1992) New plant binary vectors with selectable markers located proximal to the left T-DNA border. Plant Mol Biol 20:1195–1197
Birnbaum K, Benfey PN (2004) Network building: transcriptional circuits in the root. Curr Opin Plant Biol 7:582–588
Cahoon AB, Harris FM, Stern DB (2004) Analysis of developing maize plastid reveals two mRNA stability classes correlating with RNA polymerase type. EMBO Reports 5:801–806
Chang C-C, Sheen J, Bligny M, Niwa Y, Lerbs-Mache S, Stern DB (1999) Functional analysis of two maize cDNAs encoding T7-like RNA polymerases. Plant Cell 11:911–926
Clough SJ, Bent AF (1998) Floral dip: a simplified method for Agrobacterium-mediated transformation of Arabidopsis thaliana. Plant J 16:735–743
Drews GN, Bowman JL, Meyerowitz EM (1991) Negative regulation of the Arabidopsis homeotic gene AGAMOUS by the APETALA2 product. Cell 65:991–1002
Elorza A, Leon G, Gomez I, Mouras A, Holuigue L, Araya A, Jordana X (2004) Nuclear SDH2-1 and SDH2-2 genes, encoding the iron-sulfur subunit of mitochondrial complex II in Arabidopsis, have distinct cell-specific expression patterns and promoter activities. Plant Physiol 136:4072–4087
Emanuel C, Weihe A, Graner A, Hess WR, Börner T (2004) Chloroplast development affects expression of phage-type RNA polymerases in barley leaves. Plant J 38:460–472
Fujie M, Kuroiwa H, Kawano S, Kuroiwa T (1993) Studies on the behaviour of organelles and their nucleoids in the root apical meristem of Arabidopsis thaliana Col. Planta 189:443–452
Hedtke B, Börner T, Weihe A (1997) Mitochondrial and chloroplast phage-type RNA polymerases in Arabidopsis. Science 277:809–811
Hedtke B, Börner T, Weihe A (2000) One RNA polymerase serving two genomes. EMBO Reports 1:435–440
Hedtke B, Legen J, Weihe A, Herrmann RG, Börner T (2002) Six active phage-type RNA polymerase genes in Nicotiana tabacum. Plant J 30:625–637
Hedtke B, Meixner M, Gillandt S, Richter E., Börner T, Weihe A (1999) Green fluorescent protein as a marker to investigate targeting of organellar RNA polymerases of higher plants in vivo. Plant J 17:557–561
Hess WR, Börner T (1999) Organellar RNA polymerases of higher plants. Int Rev Cytol 190:1–59
Huang J, Struck F, Matzinger DF, Levings CS III (1994) Flower-enhanced expression of a nuclear-encoded mitochondrial respiratory protein is associated with changes in mitochondrion number. Plant Cell 6:439–448
Ikeda TM, Gray MW (1999) Identification and characterization of T7/T3 bacteriophage-like RNA polymerase sequences in wheat. Plant Mol Biol 40:567–578
Jefferson RA, Kavanagh TA, Bevan MW (1987a) GUS fusions: beta-glucuronidase as a sensitive and versatile gene fusion marker in higher plants. EMBO J 6:3901–3907
Jefferson RA, Bevan M, Kavanagh T (1987b) The use of the Escherichia coli beta-glucuronidase as a gene fusion marker for studies of gene expression in higher plants. Biochem Soc Trans 15:17–18
Kabeya Y, Sato N (2005) Unique translation initiation at the second AUG codon determines mitochondrial localization of the phage-type RNA polymerases in the moss Physcomitrella patens. Plant Physiol 138:369–382
Kobayashi Y, Dokiya Y, Sugita M (2001a) Dual targeting of phage-type RNA polymerase to both mitochondria and plastids is due to alternative translation initiation in single transcripts. Biochem Biophys Res Commun 289:1106–1113
Kobayashi Y, Dokiya Y, Kumazawa Y, Sugita M (2002) Non-AUG translation initiation of mRNA encoding plastid-targeted phage-type RNA polymerase in Nicotiana sylvestris. Biochem Biophys Res Commun 299:57–61
Kobayashi Y, Dokiya Y, Sugiura M, Niwa Y, Sugita M (2001b) Genomic organization and organ-specific expression of a nuclear gene encoding phage-type RNA polymerase in Nicotiana sylvestris. Gene 279:33–40
Kuroiwa T, Fujie M, Kuroiwa H (1992) Studies on behavior of mitochondrial DNA: synthesis of mitochondrial DNA occurs actively in a specific region just above the quiescent centre in the root meristem of Pelargonium zonale. J Cell Sci 101:483–493
Kusumi K, Yara A, Mitsui N, Tozawa Y, Iba K (2004) Characterization of a rice nuclear-encoded plastid RNA polymerase gene OsRpoTp. Plant Cell Physiol 45:1194–1201
Lee S, Warmke HE (1979) Organelle size and number in fertile and T-cytoplasmatic male-sterile corn. Am J Bot 60:141–148
Li XQ, Zhang M, Brown GG (1996) Cell-specific expression of mitochondrial transcripts in maize seedlings. Plant Cell 8:1961–1975
Liere K, Kaden D, Maliga P, Börner T (2004) Overexpression of phage-type RNA polymerase RpoTp in tobacco demonstrates its role in chloroplast transcription by recognizing a distinct promoter type. Nucleic Acids Res 32:1159–1165
Ribichich KF, Tioni MF, Chan RL, Gonzalez DH (2001) Cell-type-specific expression of plant cytochrome c mRNA in developing flowers and roots. Plant Physiol 125:1603–1610
Topping JF, Leaver CJ (1990) Mitochondrial gene expression during wheat leaf development. Planta 182:399–407
Touzet P, Budar F (2004) Unveiling the molecular arms race between two conflicting genomes in cytoplasmic male sterility? Trends Plant Sci 9:568–570
Weihe A (2004) The transcription of plant organelle genomes. In: Daniell H, Chase CD (eds) Molecular biology and biotechnology of plant organelles. Springer, Berlin Heidelberg New York, pp 213–237
Weihe A, Hedtke B, Börner T (1997) Cloning and characterization of a cDNA encoding a bacteriophage-type RNA polymerase from the higher plant Chenopodium album. Nucleic Acids Res 25:2319–2325
Young DA, Allen RL, Harvey AJ, Lonsdale DM (1998) Characterization of a gene encoding a single-subunit bacteriophage-type RNA polymerase from maize which is alternatively spliced. Mol Gen Genet 260:30–37
Acknowledgements
We gratefully acknowledge the help of Bettina Linke with in situ hybridization techniques, and Alexandra Viola Bohne and Cornelia Stock with plant propagation and DNA preparations. This work was supported by a grant from the Deutsche Forschungsgemeinschaft, Bonn (SFB 429, TP A5).
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Emanuel, C., von Groll, U., Müller, M. et al. Development- and tissue-specific expression of the RpoT gene family of Arabidopsis encoding mitochondrial and plastid RNA polymerases. Planta 223, 998–1009 (2006). https://doi.org/10.1007/s00425-005-0159-y
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00425-005-0159-y