
Abstract
Antibodies against Brassica napus cytosolic pyruvate kinase (PKc) (EC 2.7.1.40) were employed to examine PKc subunit composition and developmental profiles in castor and soybean seeds. A 56-kDa immunoreactive polypeptide was uniformly detected on immunoblots of clarified extracts from developing castor endosperm or soybean embryos. Maximal PKc activities occurred early in castor oil seed (COS) and soybean development (7.1 and 5.5 (μmol of pyruvate produced/min) g−1 FW, respectively) and were up to 25-fold greater than those of fully mature seeds. Time-course studies revealed a close correlation between extractable PKc activity and the relative amount of the immunoreactive 56-kDa PKc polypeptide. PKc from developing COS was purified 1,874-fold to homogeneity and a final specific activity of 73.1 (μmol of pyruvate produced/min) mg−1 protein. Gel filtration and SDS-PAGE indicated that this PKc exists as a 230-kDa homotetramer composed of 56-kDa subunits. The mass fingerprint of tryptic peptides of the 56-kDa COS PKc subunit best matched three putative PKcs from Arabidopsis thaliana. The purified enzyme was relatively heat-stable and displayed a broad pH optimum of 6.4. However, more efficient substrate utilization (in terms of V max /K m for phosphoenolpyruvate or ADP) was observed at pH 7.4. Glutamate was the most effective inhibitor, whereas aspartate functioned as an activator by partially relieving glutamate inhibition. Together with our previous studies, the results: (1) allow a model to be formulated regarding the coordinate allosteric control of PKc and phosphoenolpyruvate carboxylase by aspartate and glutamate in developing COS, and (2) provide further biochemical evidence that castor plant PKc exists as tissue-specific isozymes that exhibit substantial differences in their respective physical and regulatory properties.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
In developing seeds, the partitioning of imported photosynthate between starch, storage lipid, and storage protein biosynthesis is of major agronomic concern. The metabolism of the glycolytic intermediate phospho pyruvate (PEP) appears to play a prominent role in directing seed carbohydrates towards plastidic fatty acid biosynthesis versus the mitochondrial production of ATP and organic acids required for amino acid interconversion in support of storage protein biosynthesis (Ruuska et al. 2002; Schwender et al. 2004). An important enzyme in glycolytic PEP metabolism is pyruvate kinase (PK; EC 2.7.1.40) which catalyzes the irreversible transfer of Pi from PEP to ADP, yielding pyruvate and ATP.
In contrast to non-plant PKs, the enzyme from plants exists as both cytosolic (PKc) and plastid (PKp) isozymes that differ markedly in their respective physical, immunological and kinetic/regulatory characteristics (Plaxton 1989, 1996; Plaxton et al. 1990, 2002). In addition, plant PKc appears to be expressed as tissue-specific isozymes that demonstrate substantial differences in their respective physical and/or kinetic and regulatory properties. Feedback allosteric control of plant PKc isozymes by various metabolite effectors has been hypothesized to modulate its in vivo activity in accordance with the cell’s momentary demands for tricarboxylic acid cycle and respiratory endproducts; namely ATP and/or C-skeletons that serve as biosynthetic precursors. For example, in aerobic germinating castor oil seed (COS) endosperm, PKc is inhibited by several metabolite effectors to facilitate the massive gluconeogenic conversion of stored triacylglycerides to hexose-phosphates, and activated during anaerobiosis in order to help offset reduced ATP levels that accompany anoxia stress (Kobr and Beevers 1971; Podestá and Plaxton 1991). By contrast, purified PKc from germinated COS cotyledons and expanding castor leaves (Hu and Plaxton 1996; Podestá and Plaxton 1994), as well as ripening banana fruit and Brassica napus suspension cells (Smith et al. 2000; Turner and Plaxton 2000), demonstrated potent inhibition by glutamate which was reversed by the activator aspartate. Reciprocal control by aspartate and glutamate was specific for these amino acids and provided a rationale for the in vivo activation of PKcthat occurs during periods of enhanced NH +4 -assimilation. The abnormal growth, carbon partitioning and dark respiration of transgenic tobacco lacking leaf PKc (Grodzinski et al. 1999; Knowles et al. 1998) highlighted the importance of this enzyme in the control and integration of plant carbon and energy metabolism. The surprising biochemical complexity of vascular plant PK has been corroborated by recent genome sequencing initiatives (currently the NCBI-CDD (Marchler-Bauer et al. 2002) identifies 15 Arabidopsis and 8 rice genes that putatively encode different PK polypeptides).
Primary PEP metabolism in the plant cytosol is also mediated by PEP carboxylase (PEPC; EC 4.1.1.31) in a Mg2+-dependent ß-carboxylation yielding oxaloacetate and Pi. PEPC is particularly abundant in C4- and CAM-leaves where it catalyses the initial fixation of atmospheric CO2. A pivotal role for non-photosynthetic and C3-leaf PEPCs is the anaplerotic replenishment of tricarboxylic acid cycle intermediates that are withdrawn for biosynthesis and N-assimilation (Chollet et al. 1996; Plaxton 1996). PEPC has been purified from several non-photosynthetic tissues and shown to be controlled by a variety of allosteric effectors (particularly inhibition by malate, glutamate and aspartate, and activation by hexose-phosphates) and reversible protein kinase-mediated phosphorylation (Blonde and Plaxton 2003; Law and Plaxton 1995, 1997; Moraes and Plaxton 2000; Schuller et al. 1990; Tripodi and Plaxton 2005; Zhang et al. 1995).
Consistent with the view that PKc and PEPC are principal control sites of plant glycolysis (Plaxton 1996), was the application of metabolic control analysis to assess the distribution of respiratory flux control in tubers of transgenic potato plants containing modified levels of ATP-dependent phosphofructokinase (Thomas et al. 1997). It was determined that phosphofructokinase exerts a low flux control coefficient over plant glycolysis and respiration, whereas far more flux control was exerted in the oxidative metabolism of PEP. The relatively low flux control coefficient of plant phosphofructokinase was explained as a consequence of its potent feedback allosteric inhibition by PEP. In this way PK and PEPC play a central role in the overall regulation of plant glycolysis and respiration, since the control of their activities in vivo will ultimately dictate the rate of mobilization of starch or sucrose for respiration, while controlling the provision of: (1) pyruvate for ATP production via mitochondrial oxidative phosphorylation, and (2) tricarboxylic acid cycle C-skeletons needed for N-assimilation or as biosynthetic precursors.
Despite the obvious importance of PKc and PEPC in controlling plant respiration and cytosolic PEP partitioning, the two enzymes have thus far been subjected to concurrent biochemical characterization in only four systems—Brassica napus suspension cell cultures (Moraes and Plaxton 2000; Smith et al. 2000), ripening banana fruit (Law and Plaxton 1995, 1997; Turner and Plaxton 2000), germinating COS cotyledons (Podestá and Plaxton 1994), and soybean root nodules (McCloud et al. 2001; Schuller et al. 1990; Zhang et al. 1995). Novel high and low M r PEPC isoforms from developing COS were recently purified and characterized (Blonde and Plaxton 2003). In the present study, we describe the purification to homogeneity of developing COS PKc and document the physical, immunological, and kinetic properties of the purified enzyme. The combined results of the present and earlier (Blonde and Plaxton 2003) studies indicate an important role for cytosolic pH, and the allosteric effectors glutamate and aspartate in the synchronous control of PKc and PEPC in developing COS. In addition, we examined PKc subunit structure and activity profiles during the development of COS endosperm and soybean embryos.
Materials and methods
Chemicals and plant material
ADP, ADP-agarose, alkaline phosphatase-conjugated goat anti-(rabbit IgG) IgG, 2,2′-dipyridyl disulfide, NADH, PEP, and rabbit muscle lactate dehydrogenase (EC 1.1.1.27) were purchased from Sigma Chemical Co. Butyl-Sepharose fast flow resin, a Superose-6 HR10/30 gel filtration column, and M r standards were from Amersham Biosciences. Microcystin-LR was from Alexis Biochemicals. Poly(vinylidene fluoride) (PVDF) membranes (0.45 μM) were from Millipore. Fractogel EMD DEAE-650 (S) anion-exchange resin and all other chemicals were from Merck/BDH. Affinity-purified rabbit anti-(Brassica napus suspension cell culture or germinating COS PKc or developing COS PKp) IgGs were obtained as described previously (Plaxton 1989; Plaxton et al. 1990; Smith et al. 2000).
Castor plants (Ricinus communis L., cv Baker 296) were cultivated in a greenhouse at 24°C and 60% relative humidity under natural light supplemented with 16 h of artificial light (600 μmol m−2 s−1). Soybean plants (Glycine max L. cv Maple Glen) were grown in growth chambers at 30°C, 70% relative humidity and 16 h artificial light (600 μmol m−2 s−1). COS and soybean seeds were harvested at various stages of development as previously described (Greenwood and Bewley 1982; Fehr et al. 1971). Developing COS endosperm and soybean embryos were quickly dissected, frozen under liquid N2 and stored at −80°C until used.
Enzyme assays
The PK reaction was coupled to the lactate dehydrogenase reaction and assayed at 25°C by monitoring the oxidation of NADH at 340 nm using a SpectraMax 250 Microplate Spectrophotometer (Molecular Devices). Unless otherwise specified, standard PKc assay conditions were 50 mM HEPES-KOH (pH 6.4), 25 mM KCl, 12 mM MgCl2, 2 mM PEP, 1 mM ADP, 1 mM DTT, 5% (w/v) poly(ethylene)glycol (PEG) 8000, 0.15 mM NADH and 2 units/ml desalted lactate dehydrogenase in a final volume of 250 μl. PEG-8000 was routinely added to the reaction mixture because this organic solute significantly enhances the activity of homogeneous germinating COS endosperm PKc by stabilizing the native tetrameric enzyme in dilute solutions (Podestá and Plaxton 1993). All assays were corrected for contaminating PEP phosphatase by omitting ADP from the reaction mixture, and linear with respect to time and concentration of enzyme assayed. One unit of PKc activity is defined as the amount of enzyme resulting in the production of 1 μmol pyruvate/min at 25°C.
Kinetic studies and protein concentration determination
Apparent V max (V max,app) and K m values were calculated from the Michaelis-Menten equation fitted using a non-linear least-squares regression computer kinetics program (Brooks 1994). The I 50 values represent the concentration of inhibitor required to achieve 50% inhibition of enzymatic activity and were determined using the aforementioned computer kinetics program. All kinetic parameters are the means of three or more independent determinations and reproducible to within ±10% (SE) of the mean value. Free and metabolite-bound Mg2+ concentrations were calculated using a computer program that corrects for temperature, pH and ionic strength (Brooks and Storey 1992). PKc activity was independent of free Mg2+ concentration in the range 4–20 mM. For kinetic studies, a minimal free Mg2+ concentration of 8 mM was maintained by making organic anion stock solutions equimolar with MgCl2. Substrate concentrations stated in the text refer to their total concentration in the assay medium.
Protein concentrations were determined using a Coomassie Blue G-250 dye-binding method with bovine gamma-globulin as the protein standard (Bollag et al. 1996).
Preparation of clarified extracts from developing COS and soybean seeds, and purification of PKc from developing COS
All procedures were conducted at 0–4°C unless otherwise indicated. For developmental studies, freshly harvested and quick-frozen developing COS endosperms or soybean embryos were homogenized (1:2; w/v) with a Polytron for 20 s in 50 mM HEPES-KOH (pH 7.5) containing 0.1% (w/v) Triton X-100, 20% (v/v) glycerol, 4% (w/v) PEG-8000, 1% (w/v) insoluble poly(vinylpolypyrridoline), 5 mM malate, 5 mM thiourea, 1 mM DTT, 5 mM MgCl2, 1 mM EDTA, 1 mM EGTA, 20 mM NaF, 5 μg/ml chymostatin, 2 mM 2,2′-dipyridyl disulfide, 2 mM phenylmethylsulfonyl fluoride and 50 nM microcystin-LR. Homogenates were centrifuged at 16,000g for 6 min and the resulting supernatants used as the enzyme source. To ensure that the measurement of PKc activities in the COS or soybean extracts was not affected by the presence of endogenous enzyme inhibitor(s), equal proportions of COS endosperm or soybean cotyledon tissue at various developmental stages were combined and extracted. Activities of the resultant mixed extracts were always 90–110% of that predicted from the measurements made on separate samples.
PKc was isolated from 120 g of endosperm tissue of stage VII (full cotyledon) developing COS during the concomitant purification of high and low M r PEPC isoforms (PEPC1 and PEPC2) from the same tissue. Initial steps for PEPC/PKc purification involving PEG fractionation, followed by Butyl-Sepharose hydrophobic interaction and Fractogel EMD DEAE-650 (S) anion-exchange FPLC were as previously described (Blonde and Plaxton 2003). Under the conditions employed, PKc did not absorb onto the DEAE-Fractogel column and was thus recovered in the non-binding eluate. This fraction was subsequently brought to 25% (w/v) PEG 8000, stirred for 30 min, and centrifuged at 14,000g for 30 min. The pellet was resuspended in 7 ml of 50 mM HEPES-KOH (pH 7.2) containing 15% (v/v) glycerol, 1 mM DTT, 5 mM MgCl2, 1 mM EDTA, 1 mM EGTA, and 20 mM NaF (buffer A). The sample was incubated at 55°C for 3 min, chilled for 5 min on ice and centrifuged at 14,000g for 10 min. ADP-agarose affinity chromatography was performed at room temperature. The supernatant from the heat treatment step was loaded at 0.3 ml/min onto a column (3 cm long, 1 cm i.d.) of ADP-agarose (Sigma catalogue#A-4398) that had been pre-equilibrated in buffer A. The column was washed with 12 ml of buffer A and the enzyme eluted with 5 ml of buffer A containing 1 mM PEP. Pooled fractions were desalted into buffer A and simultaneously concentrated using an Amicon Centricon-10 ultrafilter, divided into 25 μl aliquots, frozen in liquid N2, and stored at −80°C. The activity of the final preparation was stable for at least 2 months when stored frozen.
Determination of native molecular mass via gel filtration FPLC
This was performed by FPLC on a Superose 6 HR 10/30 column preequilibrated with 50 mM HEPES-KOH (pH 7.2) containing 100 mM KCl, 10 mM MgCl2, 1 mM DTT, 1 mM EDTA, and 10% (v/v) glycerol. Purified PKc (100 μl, 0.45 mg/ml) was chromatographed at 0.2 ml/min. Fractions of 0.25 ml were collected and assayed for PKc activity and A 280. Native M r was calculated from a plot of K D (partition coefficient) versus log M r using the following protein standards: thyroglobulin (669-kDa), apoferritin (443-kDa), catalase (232-kDa), aldolase (158-kDa) and BSA (66-kDa). The column’s void volume was determined using Blue Dextran.
Electrophoresis and immunoblotting
SDS-PAGE was performed using a Bio-Rad minigel apparatus as previously described (Turner and Plaxton 2000). Final acrylamide concentration in the 0.75 mm-thick slab gels was 10% (w/v) for the separating gel and 4% (w/v) in the stacking gel. Extraction of developing COS under denaturing conditions using 10% (w/v) trichloroacetic acid was performed according to Wu and Wang (1984). Prior to electrophoresis all samples were incubated for 3 min at 100°C in the presence of 50 mM KPi (pH 6.8), 50 mM DTT, 10% (v/v) glycerol and 1% (w/v) SDS. SDS-PAGE was performed at 200 V for 45 min and the gel stained with Coomassie blue R-250. Subunit M r was calculated from a plot of relative mobility versus (log M r ) with the following standard proteins: myosin (200-kDa), ß-galactosidase (116-kDa), phosphorylase (97-kDa), BSA (66-kDa), ovalbumin (45-kDa), carbonic anhydrase (31-kDa), soybean trypsin inhibitor (22-kDa), lysozyme (14-kDa) and aprotinin (7-kDa).
Immunoblotting was performed using affinity-purified IgGs raised in rabbits against B. napus PKc (Smith et al. 2000), COS PKc (Plaxton 1989), or COS PKp (Plaxton et al. 1990). Antigenic polypeptides were visualized using an alkaline phosphatase-conjugated secondary antibody and chromogenic detection using 5-Br-4-Cl-3-indoyl phosphate and nitro blue tetrazolium as described in (Plaxton 1989). Phosphatase staining was for 5–10 min at 30°C. Immunological specificity was confirmed by performing immunoblots in which rabbit pre-immune serum was substituted for the anti-(B. napus or COS PKc)-IgG. Densitometric scanning of the blots was performed using an LKB Ultroscan XL Enhanced laser densitometer with data analysis using the LKB GelScan XL software (version 2.1). Immunoreactive polypeptides were quantified in terms of relative absorbance units at 633 nm. Immunoquantification of clarified extracts was linear in the concentration range of 5–50 μg of protein.
Mass spectrometry
The approximate 56-kDa Coomassie blue-staining polypeptide corresponding to purified COS PKc (see below) was excised from a SDS gel, reduced, alkylated, and digested with trypsin according to standard protocols (Shevchenko et al. 1996). Resultant tryptic peptides were mixed 1:1 (v/v) with 5 mg/ml α-cyanohydroxycinnamic acid matrix in 50% (v/v) acetonitrile and 0.1% (v/v) trifluoroacetic acid, and analyzed using a Micromass MALDI-TOF mass spectrometer in positive ion mode. The Aldente search tool (Gasteiger et al. 2005) was used to match peptide mass fingerprints using the non-redundant Viridiplantae UniProt/TrEMBL data set.
Results
Developmental profiles of cytosolic pyruvate kinase: subunit composition, activity, and amount in developing castor and soybean seeds
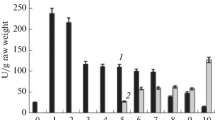
To visualize immunoreactive PKc polypeptides of developing COS endosperms and soybean embryos, immunoblots of the respective developmental time-course extracts were probed with affinity-purified polyclonal antibodies raised against homogeneous PKc from B. napus suspension cells (Fig. 1). The immunoblots uniformly revealed an immunoreactive polypeptide of approximately 56-kDa. A faint immunoreactive polypeptide of about 50-kDa was also detected on immunoblots of stage IX (maturation phase) developing COS, as well as fully mature COS (Fig 1a). Identical results were observed on immunoblots of clarified extracts prepared under denaturing conditions in the presence of 10% trichloroacetic acid according to Wu and Wang (1984), or when parallel immunoblots were probed with affinity-purified rabbit polyclonal antibodies prepared against homogeneous PKc isolated from germinated endosperms of 5-day-old COS seedlings (results not shown). A progressive and marked decrease in the relative quantity of the 56-kDa immunoreactive PKc polypeptide was observed throughout soybean and COS development and this paralleled a similar reduction in extractable PKc activity (Figs. 1 and 2). All PKc activities reported in Fig. 2 were corrected for contaminating PEP phosphatase activity by conducting parallel assays in which the PKc cosubstrate ADP was omitted from the PKc reaction mixture. Developing COS endosperm contains significant levels of both PKc and leucoplast PK (PKp) isozymes (Ireland et al. 1980; Plaxton 1989, Plaxton et al. 1990). However, purified PKp from non-green plant cells including developing COS endosperm and B. napus suspension cells exhibited a sharp alkaline pH activity optima of about pH 8.0 (with undetectable activity at pH ≤ 6.8), whereas the corresponding PKc demonstrates a typically broad pH-activity profile centered around pH 6.4–7.0 (Ireland et al. 1980; Plaxton et al. 2002). As the PK activities reported in Fig. 2 were determined at pH 6.4, they reflect PKc rather than a combination of PKc and PKp activities.

Immunological detection and SDS-PAGE analyses of developing seed PKc. a, b Extracts were subjected to SDS-PAGE (10% (w/v) separating gel) and blot transferred to a PVDF membrane. Immunoblots were probed with an affinity-purified rabbit IgG raised against homogeneous PKc from B. napus suspension cells (Smith et al. 2000). Immunoreactive polypeptides were visualized using an alkaline phosphatase-conjugated secondary antibody and chromogenic staining. Lanes labeled ‘pure PKc’ contain 20 ng of purified PKc from stage VII developing COS. The remaining lanes contain clarified extracts (corresponding to 1.0 mg of fresh tissue) from various stages of COS (a) or soybean (b) development. Lanes labeled “Dry seed” designates a fully mature COS or soybean. a Lanes (developmental stage) III, V, VII, and IX correspond to the heart-shaped embryo, mid-cotyledon, full cotyledon, and maturation stages of COS development, respectively (Greenwood and Bewely 1982). b Lanes R4, R5, and R6 correspond to the beginning seed, full seed, and beginning maturity stages of soybean development, respectively (Fehr et al. 1971). c SDS-PAGE analysis of 2 μg of purified PKcfrom stage VII developing COS. The lane labeled ‘M’ contains 5 μg of various protein standards. Protein staining was performed with Coomassie Blue R-250. O, origin; TD, tracking dye front

Developmental profiles of PKc activity and relative amount, and soluble protein concentration in endosperm of developing COS (a) and cotyledons of developing soybeans (b). PKc activities and soluble protein concentrations represent the means ± SE of n=3 separate extracts. Relative amounts of PKc were estimated by laser densitometry of immunoreactive 56-kDa PKc polypeptides on immunoblots of clarified extracts (equivalent to 1.0 mg fresh weight) that had been probed with anti-(B. napus PKc)-IgG as described in Fig. 1. Values represent the mean of two different extracts
Purification, and physical and immunological characterization of PKc from developing COS endosperm
As shown in Table 1, PKc from endosperms of stage VII developing COS was purified approximately 1,900-fold to a final specific activity of 73.1 units/mg and an overall recovery of 10%. The final preparation was devoid of PEP phosphatase activity. Affinity chromatography on ADP-agarose was particularly effective as it resulted in an approximate 100-fold purification and 60% recovery. SDS-PAGE of the final preparation confirmed that the enzyme was purified to homogeneity (Fig. 1c). Similar to other plant PKcs (Hu and Plaxton 1996; Plaxton 1988, 1989; Podestá and Plaxton 1994; Smith et al. 2000; Turner and Plaxton 2000), the purified COS PKc was relatively heat stable, retaining 100, 85, 48, and 6% of its activity following its 3 min incubation at 55, 60, 65, and 75°C, respectively. This property was exploited during PKc purification, with the 55°C heat treatment step providing a 23% increase in specific activity with a 100% yield (Table 1). By contrast, plant PKp isozymes (including developing COS PKp) are quite heat labile, being completely inactivated following a similar heat treatment (Ireland et al. 1980; Plaxton et al. 1990, 2002; Smith et al. 2000).
The single Coomassie blue staining 56-kDa polypeptide observed following SDS-PAGE of the final COS PKc preparation cross-reacted strongly with anti-(B. napus PKc)-IgG and co-migrated with the immunoreactive 56-kDa polypeptide observed on immunoblots of clarified extracts from developing COS or soybean (Fig. 1). By contrast, no immunoreactive bands were observed when an immunoblot of up to 100 ng of the final preparation was probed with anti-(developing COS PKp)-IgG (results not shown). Tryptic peptides of the 56-kDa polypeptide corresponding to purified COS PKc (Fig. 1c) were analyzed by MALDI-TOF MS. The resultant peptide mass fingerprint best matched three putative PKcs from Arabidopsis (gene loci: At3g52990, At2g36580, and At5g63680) with each having a predicted M r of approximately 56-kDa. The most significant match (At3g52990) exhibited peptide coverage of 17% and an expect value of 10−9 . Taken together, these data indicate that the final preparation consisted only of PKc and that it was not proteolytically degraded. The enzyme’s native M r was estimated to be 230-kDa by gel filtration FPLC on a calibrated Superose-6 HR 10/30 column. Thus, in agreement with an earlier report on partially purified developing COS PKc (Plaxton 1989), this enzyme appears to exist as a homotetramer composed of identical 56-kDa subunits.
Kinetic properties of developing COS PKc
The influence of pH on the activity of the purified enzyme was determined over the pH range of 5.0–8.25 using a mixture of 25 mM MES and 25 mM Bis-Tris propane as the assay buffer. The enzyme exhibited a broad, symmetrical pH/activity profile with maximal activity occurring at about pH 6.4 (results not shown). Subsequent kinetic studies were routinely performed using 50 mM HEPES buffer at pH 6.4 and 7.4 where PKc activity was approximately 80% of the maximum. At either pH value, the enzyme displayed equivalent activity in 50 mM HEPES or a mixture of 25 mM MES and 25 mM Bis-Tris propane as the assay buffer.
Co-factor requirements and substrate saturation kinetics
Similar to most known PKs, the activity of developing COS PKc was absolutely dependent on the presence of a monovalent and bivalent cation with K+ and Mg2+ fulfilling this requirement. At pH 6.4 the enzyme’s apparent K m value for free Mg2+ was 0.32 mM. The enzyme exhibited hyperbolic saturation kinetics for PEP and ADP at both pH 6.4 and 7.4 (results not shown). In contrast to germinating COS endosperm PKc which demonstrated substrate inhibition by ADP at concentrations greater than 1 mM (Podestá and Plaxton 1991), no inhibition of developing COS PKc by ADP (up to 5 mM) was observed. V max,app and K m values obtained for PEP and ADP were respectively about 20% and 50% lower at pH 7.4 than at pH 6.4 (Table 2). As a consequence, the enzyme’s catalytic efficiency (V max,app/K m ) with respect to PEP or ADP was approximately 40% greater at pH 7.4 than at pH 6.4 (Table 2).
Metabolite effects
A wide variety of compounds were tested for effects on PKc activity at pH 6.4 and 7.4 with subsaturating concentrations of PEP and ADP (0.1 mM each). The following compounds exerted little or no influence (±15% control velocity) on enzymatic activity: 10 mM sucrose; glucose, glucose 6-phosphate, glucose 1-phosphate, fructose, fructose 6-phosphate, 6-phosphogluconate, ribose 5-phosphate, dihydroxyacetone-phosphate, glycerol 3-phosphate, 2-phosphoglycerate, phosphoglycolate, 2-oxoglutarate, succinate, glutamine, aspartate, asparagine, alanine, serine, lysine, methionine, cysteine, arginine, histidine, acetate, acetaldehyde, NH4Cl, AMP, and Pi (5 mM each); threonine, valine, isoleucine, tyrosine, phenlyalanine, proline, and acetyl-CoA (0.5 mM each); 0.2 mM rutin; and 0.05 mM fructose 2,6-bisphosphate. Table 3 lists those compounds that significantly influenced the activity of the purified enzyme.
Glutamate was the most effective inhibitor of developing COS PKc with inhibition being somewhat more pronounced at pH 7.4 as compared to pH 6.4 (Table 3). Glutamate significantly reduced PKc’s V max,app and affinity for PEP (Table 2). As reported for some plant PKcs, including purified PKc from expanding castor leaves (Hu and Plaxton 1996), aspartate partially reversed the inhibitory effect of glutamate on PKc at both pH values (Fig. 3). This appears to be a very specific effect, since aspartate did not influence the response of the enzyme to the other inhibitors listed in Table 3, and did not activate the enzyme in the absence of glutamate. As asparagine and glutamine are endproducts of NH +4 assimilation, their influence on glutamate inhibition of PKc was also examined. However, the enzyme’s I T 50 (glutamate) was unchanged by the inclusion of glutamine or asparagine (10 mM each) in the assay mixture.

Influence of aspartate on the inhibition of developing COS endosperm PKc by glutamate. Assays were conducted at pH 6.4 (a) and pH 7.4 (b) in the presence of subsaturating concentrations of PEP and ADP (0.1 mM each). The enzyme’s I 50 values for glutamate (c) were determined in the presence of various concentrations of aspartate. All values are the means of at least three independent determinations and are reproducible to within ±10% (SE) of the mean value
Discussion
Immunoblotting of clarified extracts from soybean cotyledons or developing COS endosperm using monospecific anti-(B. napus PKc)-IgG revealed an immunoreactive 56-kDa polypeptide at all stages of seed development (Fig. 1). Developmental profiles of PKc activity correlated well with the relative amount of PKc (Fig. 2). That the 56-kDa immunoreactive polypeptide observed on immunoblots of developing soybean or COS extracts corresponds to a bona fide PKc polypeptide was corroborated by its co-migration with the single protein-staining and immunoreactive 56-kDa polypeptide of the PKc purified to apparent homogeneity from endosperms of stage VII developing COS (Fig. 1). In a related study, Tang and co-workers (2003) produced a polyclonal antibody against a 15 amino acid peptide corresponding to the N-terminus of a putative soybean seed PKc to monitor PKc subunit composition during soybean embryogenesis. Their results are in accord with those of Figs. 1b and 2b since both studies demonstrated that extractable PKc activity and the amount of an immunoreactive PKc polypeptide of approximately 56-kDa both show a marked reduction during soybean embryogenesis. They also reported a parallel reduction in transcript levels encoding a putative PKc of about 56-kDa in developing soybeans, with these transcripts becoming undetectable during the later stages of seed development (Tang et al. 2003). Likewise, a cDNA microarray study of developing Arabidopsis seeds concluded that PKc gene expression showed a marked reduction as seed development progressed (Ruuska et al. 2002). Taken together, the results imply that oilseed PKc synthesis versus proteolytic turnover is tightly controlled such that net synthesis predominates earlier in seed development with proteolytic turnover prevailing during seed maturation. Tang and coworkers (2003) also observed the progressive accumulation of a 51-kDa immunoreactive polypeptide during soybean embryogenesis. They concluded that this arises from C-terminal cleavage of the 55-kDa full-length PKc polypeptide, and that active soybean seed PKc exists as a heterooligomeric protein composed of varying proportions of 55 and 51-kDa subunits depending upon the developmental stage. PKc phosphorylation was also assessed using phosphopeptide antibodies corresponding to a pair of putative phosphorylation sites of developing soybean seed PKc (Tang et al. 2003). Although anti-(phosphopeptide)-IgG immunoreactive polypeptides corresponding to phosphorylated full length (55-kDa) soybean PKc were not detected, numerous immunoreactive polypeptides in the range of 51–20-kDa were suggested to correspond to phosphorylated truncated forms of PKc (Tang et al. 2003). One would anticipate that any truncated soybean PKc polypeptides should cross-react with the affinity-purified anti-(B. napus and COS PKc)-polyclonal IgGs employed in the present study. Although our immunoblots of developing COS or soybean extracts uniformly revealed a prominent 56-kDa PKc polypeptide, we also observed an additional, but less intense immunoreactive polypeptide of about 50-kDa on immunoblots of stage IX developing COS, together with fully mature COS (Fig. 1a). Further studies are required to determine the generality and/or the physiological role of proteolytic PKc post-translational modification during oil seed development and to what extent this might impact on PEP partitioning due to an altered allosteric response of the enzyme.
COS PK c purification, physical and immunological properties
To further evaluate the physical and kinetic properties of PKc in developing oilseeds, as well as to extend our understanding of comparative PKc biochemistry in different tissues of the castor plant, the enzyme was purified from stage VII (full cotyledon) COS endosperm (Table 1). The COS PKc was purified to electrophoretic homogeneity (Fig. 1c) and a final specific activity of nearly 75 units/mg protein. This value compares favorably to those reported previously for homogeneous PKc from various plant sources (Table 4) (Smith et al. 2000; Turner and Plaxton 2000). Three observations confirm that the final preparation corresponded to PKc and not PKp: (1) as observed with other PKcs the enzyme was relatively heat stable, in contrast to the heat-labile PKp, (2) it exhibited a pH/activity profile typical of plant PKc, but not PKp, and (3) it readily cross-reacted with antibodies raised against plant PKc (Fig. 1a, b), but not the PKp isozyme of developing COS. Immunological distinctiveness of plant PKc and PKp isozymes has been well documented (Plaxton 1989; Plaxton et al. 1990, 2002; Smith et al. 2000). In addition, MALDI-TOF MS analysis of tryptic peptides of the final preparation yielded a mass fingerprint that best matched three putative Arabidopsis PKcs having predicted M r s of about 56-kDa, equivalent to that estimated for the purified COS PKc via SDS-PAGE (Fig. 1c). Predotar (V1.03; http://genoplante-info.infobiogen.fr/predotar/) was used to scan these putative Arabidopsis PKcs for organelle targeting sequences. In each case the algorithm did not indicate a plastid or mitochondrial targeting sequence and predicted (P>0.99) that the protein would be localized in the cytosol.
Native PKc from developing COS exists as a 230-kDa homotetramer as indicated by gel-filtration FPLC and SDS-PAGE analyses of the final preparation. Therefore, the subunit structure of developing COS PKc is similar to the 237-kDa homotetrameric PKc isolated from germinating COS cotyledons (Podestá and Plaxton 1994), but differs from castor leaf or germinating COS endosperm native PKcs which both exist as 240–250-kDa heterotetramers composed of equivalent proportions of 57 and 56-kDa subunits (Table 4) (Hu and Plaxton 1996; Plaxton 1988, 1989).
Kinetic and regulatory properties of purified PK c from developing COS
The activity of the developing COS PKc exhibited: (1) a broad symmetrical pH/activity profile with maximal activity centered around pH 6.4, (2) hyperbolic substrate saturation kinetics, and similar affinities for PEP and ADP (Table 2), (3) absolute dependence on a univalent and bivalent metal cation cofactor, and (4) greater catalytic efficiency (V max,app/K m ) at pH 7.4 (physiological pH) than pH 6.4. These characteristics are similar to PKc preparations from other plant sources (Hu and Plaxton 1996; Podestá and Plaxton 1991, 1994; Smith et al. 2000; Turner and Plaxton 2000), but contrast with PKp isolated from B. napus suspension cells and developing COS which uniformly displayed relatively sharp pH/activity profiles centered at pH 8.0 (Ireland et al. 1980; Plaxton et al. 2002).
ATP inhibition is widespread among plant and non-plant PKs. However, the I 50 values of COS PKc for ATP at pH 6.4 and 7.4 (Table 3) are well in excess of estimated levels of ATP in the plant cytosol (Stitt et al. 1982). This, along with the lack of effect of AMP, indicates that energy charge does not play a major role in regulating the in vivo activity of developing COS PKc. This contrasts with many non-plant PKs, as well as PKc from endosperm of germinated COS endosperms where regulation by energy charge appears to be a key aspect of the enzyme’s in vivo control (Podestá and Plaxton 1991, 1993). Developing COS PKc demonstrated potent inhibition by glutamate, with its response to glutamate being somewhat dampened at pH 6.4 relative to pH 7.4 (Table 3, Fig. 3). No information is available on the cytosolic glutamate concentration during development of this tissue. However, next to sucrose, glutamate is the most highly concentrated compound in the cytosol of many plant cells (Heldt 1997). For example, the glutamate concentration of the leaf cytosol ranges from about 20–90 mM (Heinke et al. 1997). Glutamate is thus expected to have major significance in the in vivo allosteric control of developing COS PKc. It is intriguing that aspartate functioned as an activator of this PKc only in the presence of glutamate (Fig. 3). The inhibition of COS PKc by glutamate and its partial reversal by aspartate was specific for these particular amino acids. This reciprocal allosteric control is remarkable considering that glutamate differs from aspartate only by having an extra carbon atom in its functional group. The data indicate that the cytosolic concentration ratio of aspartate to glutamate may be a major determinant of in vivo PKc activity in developing COS. Both glutamate and aspartate have been implicated in the allosteric control of PKc and PEPC in various vascular plant tissues and green algae (Plaxton 1996), including castor leaves (Hu and Plaxton 1996), ripening banana fruit (Law and Plaxton 1995, 1997), and B. napus suspension cells (Moraes and Plaxton 2000; Smith et al. 2000). Allosteric control of plant PKcs and PEPCs by glutamate provides a tight feedback control that is hypothesized to balance their overall activity with the production of C-skeletons required for NH +4 -assimilation and transamination reactions in tissues active in amino acid and protein synthesis. By contrast, the activity of PKc isolated from endosperms of 5-d-germinated castor seedlings was insensitive to glutamate or aspartate (Table 4) (Podestá and Plaxton 1991). These differences may reflect tissue-specific PKc isozymes that have evolved to suit the distinctive metabolic requirements of developing versus germinating oil seeds (i.e., storage lipid and protein accumulation versus mobilization).
Developing soybean seed PKc was suggested to become inhibited by glutamate and activated by aspartate following its in vivo proteolytic degradation from a 55-kDa full length PKc polypeptide into a 51-kDa truncation product (Tang et al. 2003). However, non-proteolyzed PKcs with subunit M r s in the range of 57–56-kDa that have been fully purified from developing seeds and leaves of the castor plant (Table 4), banana fruit (Turner and Plaxton 2000), and B. napus suspension cells (Smith et al. 2000) all demonstrated aspartate activation by relieving the effect of glutamate inhibition. Therefore, reciprocal allosteric control of plant PKcs by glutamate and aspartate does not necessarily depend upon the enzyme’s limited proteolysis.
Coordinate control of developing COS PKc and PEPC by pH and allosteric effectors
Two PEPC isoforms from the endosperm of stage VII developing COS were recently purified and shown to be highly analogous to green algal ‘Class 1’ and ‘Class 2’ PEPC isoforms (Blonde and Plaxton 2003). The association of a shared 107-kDa catalytic subunit (p107) with an immunologically unrelated, but PEPC-like 64-kDa polypeptide (p64; related to ‘bacterial type’ plant PEPCs (Sanchez and Cejudo 2003)) appears to cause the marked physical and kinetic differences between the COS PEPC1 p107 homotetramer and the novel PEPC2 p107/p64 heterooctamer. We hypothesized that PEPC1 (activated by glucose 6-phosphate; potently inhibited by malate, aspartate and glutamate) and PEPC2 (desensitized to effectors) respectively support storage protein versus storage lipid synthesis in developing COS (Blonde and Plaxton 2003). Based on these differences and on the allosteric properties of PKc (this work) a model can be formulated for the synchronous control of PKc and PEPC in developing COS endosperm.
Glutamate inhibition of COS PKc was positively correlated with pH increases in the physiological range (Table 2, Fig. 3), whereas with PEPC1 and PEPC2 the converse was true (Blonde and Plaxton 2003). Therefore, cytosolic pH may be an important factor in coordinating the relative flux of PEP through PKc and PEPC in developing COS, with the balance moving from PEPC1 or PEPC2 to PKc upon cytosolic acidification and vice versa. As COS PEPC2 is desensitized to effectors relative to PEPC1, the flux of PEP to malate in support of leucoplast fatty acid biosynthesis may be facilitated despite the elevated levels of malate present in developing COS. In contrast, the interactive effects of aspartate and glutamate on PEPC1 and PKc suggest that PEP flux through either enzyme will be coordinated with the specific anaplerotic demands of the cell. Thus, if storage protein biosynthesis uses glutamate or glutamate-derived amino acids, the tricarboxylic acid cycle will be drained from citrate/2-oxoglutarate and necessitate an equal contribution from PKc and PEPC1 for anaplerotic replenishment. In contrast, if aspartate (or aspartate-derived amino acids) are synthesized from oxaloacetate they will only require the contribution of PEPC1. Therefore, feedback inhibition of PKc and PEPC1 by glutamate provides a rationale for the activation of both enzymes that occurs when glutamate concentrations fall as glutamate or glutamate-derived amino acids are utilized in protein biosynthesis. However, when aspartate or aspartate-derived amino acids are required for storage protein biosynthesis, flux through PEPC will be increased (due to the relief of PEPC aspartate inhibition) while flux through PKc will decrease (due to reduced aspartate relief of PKc’s glutamate inhibition). Currently, this model is limited to the known allosteric properties of PKc and PEPC from developing COS. However, developing soybean seed PKc and COS PEPC have been reported to be phosphorylated in vivo (Tang et al. 2003; Tripodi and Plaxton 2005). It will therefore be important for future studies to resolve the role that enzyme phosphorylation plays in controlling carbon partitioning at the cytosolic PEP branchpoint in developing seeds.
Tissue-specific isozymes of PKc in the castor oil plant
Vascular plant PKcs are immunologically related, heat-stable proteins, indicating their close phylogenetic relatedness. However, the present and previous studies of purified PKcs from various castor plant tissues indicate that as with mammalian PKcs, the plant enzyme exists as tissue-specific isozymes that exhibit substantial differences in various physical and kinetic/regulatory properties (Table 4). This conclusion is substantiated by recent genome sequencing initiatives indicating the existence of up to 15 and 8 different PKc genes in Arabidopsis and rice, respectively. In terms of subunit composition, the developing COS PKc most closely resembles the homotetrameric germinating cotyledon enzyme, but differs from the heterotetrameric leaf and germinating endosperm PKcs (Table 4). It is intriguing that developing versus germinated COS endosperm appears to express separate PKc isozymes that display substantial differences in their respective physical and kinetic/regulatory properties. Early in the study of plant glycolysis, PKc was demonstrated to play a fundamental role in regulating the metabolic reorganization from gluconeogenesis to glycolysis that occurs when germinated COS undergoes anaerobiosis (Kobr and Beevers 1971). Unlike the homotetrameric developing COS PKc (Fig. 3), the activity of the heterotetrameric PKc purified from germinated COS endosperm was insensitive to glutamate and aspartate (Podestá and Plaxton 1991). Current evidence suggests that the anoxia-dependent increase in the in vivo activity of germinated COS PKc (Kobr and Beevers 1971) arises from a release of the enzyme from inhibition by ATP and various glyoxylate and tricarboxylic acid cycle intermediates (Podestá and Plaxton 1991). Thus, as highlighted by the marked differences in the physical and kinetic/regulatory properties of PKc from developing versus germinated COS endosperm, the complete understanding of the control and integration of plant glycolysis awaits detailed analyses of the molecular and biochemical features of each key enzyme on a tissue-by-tissue basis.
Abbreviations
- COS :
-
Castor oil seed
- FPLC :
-
Fast protein liquid chromatography
- MALDI-TOF MS :
-
Laser desorption ionization-time of flight mass spectrometry
- PEG :
-
Poly(ethylene)glycol
- PEP :
-
Phosphoenolpyruvate
- PEPC :
-
Phosphoenolpyruvate carboxylase
- PK :
-
Pyruvate kinase
- PK c and PK p :
-
Cytosolic and plastid PK isozymes, respectively.
References
Blonde JD, Plaxton WC (2003) Structural and kinetic properties of high and low molecular mass phosphoenolpyruvate carboxylase isoforms from the endosperm of developing castor oil seeds. J Biol Chem 278:11867–11873
Bollag DM, Rozycki MD, Edelstein SJ (1996) Protein methods. Wiley, New York, pp 62–70
Brooks SPG (1994) A program for analyzing enzyme rate data obtained from a microplate reader. Biotechniques 17:1154–1161
Brooks SPG, Storey KB (1992) Bound and determined: a computer program for making buffers of defined ion concentrations. Anal Biochem 201:119–126
Chollet R, Vidal J, O’Leary MH (1996) phosphoenolpyruvate carboxylase: a ubiquitous, highly regulated enzyme in plants. Ann Rev Plant Phys Plant Mol Biol 47:273–298
Fehr WR, Caviness CE, Burmood DT, Permington JS (1971) Stage of development descriptions for soybean (Glycine max L., Merr). Crop Sci 11:929–931
Gasteiger E, Hoogland C, Gattiker A, Duvaud S, Wilkins MR, Appel RD, Bairoch A (2005) Protein identification and analysis tools on the ExPASy server. In: Walker JM (ed) The proteomics protocols handbook. Humana Press, New Jersey (in press)
Greenwood JS, Bewley JD (1982) Seed development in Ricinus communis (castor bean). I. Descriptive morphology. Can J Bot 60:1751–1760
Grodzinski B, Jiao J, Knowles VL, Plaxton WC (1999) Photosynthesis and carbon-partitioning in transgenic tobacco plants deficient in leaf pyruvate kinase. Plant Physiol 120:887–895
Heineke D, Lohaus G, Winter H, (1997) Compartmentation of C/N metabolism. In: Foyer C, Quick WP (eds) A molecular approach to primary metabolism in higher plants. Taylor & Francis Ltd, London, pp 205–217
Heldt H-W (1997) Plant biochemistry and molecular biology. Oxford University Press, New York, p 131
Hu Z, Plaxton WC (1996) Purification and characterization of cytosolic pyruvate kinase from leaves of the castor oil plant. Arch Biochem Biophys 333:298–307
Ireland RJ, Deluca V, Dennis DT (1980) Characterization and kinetics of isoenzymes of pyruvate-kinase from developing castor bean endosperm. Plant Physiol 65:1188–1193
Knowles VL, McHugh SG, Hu Z, Dennis DT, Miki BL, Plaxton WC (1998) Altered growth of transgenic tobacco lacking leaf cytosolic pyruvate kinase. Plant Physiol 116:45–51
Kobr M, Beevers H (1971) Gluconeogenesis in the castor bean endosperm. I. Changes in glycolytic intermediates. Plant Physiol 47:48–52
Law RD, Plaxton WC (1995) Purification and characterization of a novel phosphoenolpyruvate carboxylase from banana fruit. Biochem J 307:807–816
Law RD, Plaxton WC (1997) Regulatory phosphorylation of banana fruit phosphoenolpyruvate carboxylase by a copurifying phosphoenolpyruvate carboxylase-kinase. Eur J Biochem 247:642–651
Marchler-Bauer A, Panchenko AR, Shoemaker BA, Thiessen PA, Geer LY, Bryant SH (2002) CDD:a database of conserved domain alignments with links to domain three-dimensional structure. Nucleic Acids Res 30:81–283
McCloud SA, Smith RG, Schuller KA (2001) Partial purification and characterization of pyruvate kinase from the plant fraction of soybean root nodules. Physiol Plant 111:283–290
Moraes TF, Plaxton WC (2000) Purification and characterization of phosphoenolpyruvate carboxylase from Brassica napus (rapeseed) suspension cell cultures. Implications for phosphoenolpyruvate carboxylase regulation during phosphate starvation and the integration of glycolysis with nitrogen assimilation. Eur J Biochem 267:4465–4476
Plaxton WC (1988) Purification of pyruvate kinase from germinating castor bean endosperm. Plant Physiol 86:1064–1069
Plaxton WC (1989) Molecular and immunological characterization of plastid and cytosolic pyruvate kinase isozymes from castor-oil-plant endosperm and leaf. Eur J Biochem 181:443–451
Plaxton WC (1996) The organization and regulation of plant glycolysis. Ann Rev Plant Phys Plant Mol Biol 47:185–214
Plaxton WC, Dennis DT, Knowles VL (1990) Purification of leucoplast pyruvate kinase from developing castor bean endosperm. Plant Physiol 94:1528–1534
Plaxton WC, Smith CR, Knowles VL (2002) Molecular and regulatory properties of leucoplast pyruvate kinase from Brassica napus (rapeseed) suspension cells. Arch Biochem Biophys 400:54–62
Podestá FE, Plaxton WC (1991) Kinetic and regulatory properties of cytosolic pyruvate kinase from germinating castor oil seeds. Biochem J 279:495–501
Podestá FE, Plaxton WC (1993) Activation of cytosolic pyruvate kinase by polyethylene glycol. Plant Physiol 103:285–288
Podestá FE, Plaxton WC (1994) Regulation of cytosolic carbon metabolism in germinating Ricinus communis cotyledons. II. Properties of phosphoenolpyruvate carboxylase and cytosolic pyruvate kinase associated with the regulation of glycolysis and nitrogen assimilation. Planta 194:381–387
Ruuska SA, Girke T, Benning C, Ohlrogge JB (2002) Contrapuntal networks of gene expression during Arabidopsis seed filling. Plant Cell 14:1191–1206
Sánchez S, Cejudo FJ (2003) Identification and expression analysis of a gene encoding a bacterial-type phospho enol pyruvate carboxylase from Arabidopsis and rice. Plant Physiol 132:949–957
Schuller KA, Turpin DH, Plaxton WC (1990) Metabolite regulation of partially purified soybean nodule phospho enolpyruvate carboxylase. Plant Physiol 94:1429–1435
Schwender J, Ohlrogge JB, Shachar-Hill Y (2004) Understanding flux in plant metabolic networks. Curr Op Plant Biol 7:309–317
Shevchenko A, Wilm M, Vorm O, Mann M (1996) Mass spectrometric sequencing of proteins from silver-stained polyacrylamide gels. Anal Chem 68:850–858
Smith CR, Knowles VL, Plaxton WC (2000) Purification and characterization of cytosolic pyruvate kinase from Brassica napus (rapeseed) suspension cell cultures. Implications for the integration of glycolysis with nitrogen assimilation. Eur J Biochem 267:4477–4485
Stitt M, Lillet RM, Heldt HW (1982) Adenine nucleotide levels in the cytosol, chloroplasts and mitochondria of wheat leaf protoplasts. Plant Physiol 70:977–983
Tang GQ, Hardin SC, Dewey R, Huber SC (2003) A novel C-terminal proteolytic processing of cytosolic pyruvate kinase, its phosphorylation and degradation by the proteasome in developing soybean seeds. Plant J 34:77–93
Thomas S, Mooney PJ, Burrell MM, Fell DA (1997) Metabolic control analysis of glycolysis in tuber tissue of potato (Solanum tuberosum): explanation for the low control coefficient of phosphofructokinase over respiratory flux. Biochem J 322:119–127
Tripodi KE, Plaxton WC (2005) In vivo regulatory phosphorylation of phosphoenolpyruvate carboxylase in developing castor oil seeds. In: van der Est A, Bruce D (eds) Photosynthesis: fundamental aspects to global perspectives. Allen Press Inc., Lawrence, pp 915–918
Turner WL, Plaxton WC (2000) Purification and characterization of cytosolic pyruvate kinase from banana fruit. Biochem J 352:875–882
Wu FS, Wang MY (1984) Extraction of proteins for sodium dodecyl sulfate-polyacrylamide gel electrophoresis from protease rich plant tissue. Anal Biochem 139:100–103
Zhang X-Q, Li B, Chollet R (1995) In vivo regulatory phosphorylation of soybean nodule phosphoenolpyruvate carboxylase. Plant Physiol 108:1561–1568
Acknowledgment
Financial support was kindly provided by research and equipment grants from the Natural Sciences and Engineering Research Council of Canada (NSERC). We are also grateful to Prof. Steve Huber for helpful discussions, and to the Protein Function Discovery Research and Training Program (Queen’s University) for performing MALDI-TOF MS analysis of the purified COS PKc.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Turner, W.L., Knowles, V.L. & Plaxton, W.C. Cytosolic pyruvate kinase: subunit composition, activity, and amount in developing castor and soybean seeds, and biochemical characterization of the purified castor seed enzyme. Planta 222, 1051–1062 (2005). https://doi.org/10.1007/s00425-005-0044-8
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00425-005-0044-8