
Abstract
We show here that the total invertase activity in developing seeds of maize is due to two cell wall invertase (CWI) genes, Incw1 and Incw2 (Mn1). Our previous results have shown that loss-of-function mutations at the Mn1 locus lead to the miniature-1 (mn1) seed phenotype, marked by a loss of >70% of seed weight at maturity. The mn1 seed mutant is, however, non-lethal presumably because it retains a residual low level, ∼1%, of the total CWI activity relative to the Mn1 endosperm throughout seed development. Evidence here shows that the residual activity in the mn1 mutant is encoded by the Incw1 gene. RNA level analyses, especially quantitative real-time PCR studies, showed significant spatial and temporal heterogeneity in the expression of the two CWI genes in the developing endosperm. The Mn1-encoded Incw2 transcripts were seen at the highest levels in the basal region (the sugar unloading zone) during the early phase of cell division and elongation in the endosperm. In contrast, the highest levels of Incw1 transcripts were seen in the storage phase in both the upper (storage cells) and the lower parts of the endosperm. Protein and enzyme level analyses, however, appeared to show a lack of concordance with the RNA level of expression in both the Mn1 and mn1 endosperms, indicating a possibility of post-transcriptional control in the expression of these two genes. Collectively, the data suggest an important role for apoplastic cleavage of sucrose throughout the duration of seed development; and, of the two isoforms, the INCW2 appears to control metabolic flux of sugar utilization in the developing endosperm.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Cell wall invertase (CWI), localized in the apoplast and ionically bound to the cell wall, catalyzes the irreversible hydrolysis of sucrose to glucose and fructose (reviewed by Sturm 1999). The substrate of CWI, sucrose, is a major form of photosynthate for long distance transport to various sink tissues, including terminal storage organs such as developing seeds and tubers. Thus, CWI-mediated cleavage of sucrose constitutes a major entry point of sucrose carbon into numerous diverse metabolic reactions. Not surprisingly, several correlative data suggest a major physiological role for CWI in maintaining source-to-sink unloading of sucrose and ultimately in determining sink strength. In tomato, three CWIs show unique distribution profiles that are essentially associated with sink tissues (Godt and Roitsch 1997). Transgenic potato plants over-expressing CWI showed elevated tuber/leaf dry weight indicating increased sink strength of tubers and leaves (Sonnewald et al. 1997). Antisense repression of CWI in carrot resulted in altered growth of the transgenic plants at early stages of development, with markedly reduced tap root growth and severely compromised capacity for carbohydrate storage (Tang et al. 1999). Recently, two major studies have further substantiated a correlation between altered CWI activity and a modification of sink strength in transgenic plants. Leaf senescence is believed to occur due to perturbed source–sink relationships such that nutrients from a senescing leaf are diverted to other parts of the plant that have stronger sinks. Lara et al. (2004) show through transgenic approaches that ectopically increased production of CWI can delay senescence. Additionally, the increased CWI is in fact required for cytokinin-mediated delay in senescence; thus providing overall an exquisite system of cross-talk in sugar–cytokinin signaling and sink strength. Similarly, Heyer et al. (2004) demonstrated greatly altered resource allocation and a 30% gain in seed yield (through increased reproductive axillary branches) in transgenic Arabidopsis plants which express the yeast CWI gene under the control of a meristem-specific promoter.
One of the earliest genetic demonstrations on the role of CWI in sink strength in developing seeds is in maize. The miniature1 (hereafter, mn1) seed mutation is associated with a loss of >70% seed weight and is caused by loss-of-function mutation at the Mn1 locus that encodes a major endosperm-specific CWI isoform, INCW2 (Miller and Chourey 1992; Cheng et al. 1996). The highest level of INCW2 protein is seen during the early phase of endosperm development in the basal endosperm transfer cells (BETC), the single port of sugar-entry from maternal pedicel tissue to developing seed in maize (Cheng et al. 1996). Further analyses revealed that the CWI-mediated metabolic release of hexoses in the BETC region, not an exogenous supply of hexoses, is critical for resource allocation of sugars in normal endosperm development (Cheng and Chourey 1999). Recent analyses based on in situ fluorometric assay of glucose on tissue sections suggest that glucose may also enter the filial tissues during very early stages, 0–2 days after pollination (DAP), of seed development (McLaughlin and Boyer 2004). Several studies on the role of CWI in seed development, especially in establishing the roles of hexoses and sucrose in cell division and storage functions, are reported in Vicia faba (Weber et al. 1996, 1997). Consistent with these results are the maize studies which show that the CWI-deficiency in the mn1 endosperm is associated with reduced cell number and cell size when compared with the Mn1 endosperm (Vilhar et al. 2002). In developing seeds of barley, a total of three CWI genes have been identified through conventional gene expression and macro-array expression profiles and one CWI gene, HvCWINV1, is an ortholog of the maize Incw2 gene (Weschke et al. 2003; Sreenivasulu et al. 2004). The highest levels of CWI activities in barley caryopses are at the sites of sugar transport within rapidly importing apoplastic sites (Weschke et al. 2003). In rice, however, only a single CWI gene, OsCIN1, an ortholog of maize Incw1, is expressed in developing seeds (Hirose et al. 2002) with expression profiles that are similar to the maize Incw2 gene (Cheng et al. (1996). On the basis of RT-PCR analyses, Cho et al. (2005) have recently described three CWI genes in developing rice seeds. Significantly, Ohto et al. (2005) have shown seed mass control in Arabidopsis by a loss-of-function mutation at the APETALA 2 (AP2) locus, a transcription factor which causes changes in the ratio of hexose to sucrose in developing seeds. There is thus a strong possibility that AP2 may control seed mass through its effect on sugar metabolism.
In maize, a total of four CWI and two soluble invertase genes (Kim et al. 2000; Xu et al. 1996) respectively are described. Unlike CWI that is bound to the cell wall, soluble invertase is in the cytosol and/or vacuole and is extractable in the crude supernatant after cell disruption. Xu et al. (1996) reported abundant levels of the two soluble invertases, Ivr1 and Ivr2, in developing kernels. However, subsequent analyses (Carlson and Chourey 1999; Anderson et al. 2002; McLaughlin and Boyer 2004) report no Ivr1 expression in this tissue and the Ivr2 expression is predominantly localized in the maternal tissues, nucellus and pedicel, and is temporally restricted to the very early stages (up to the 8 DAP stage) of kernel development. A similar tissue profile of soluble invertase is also described in developing caryopsis of barley (Weschke et al. 2003). The metabolic storage phase of maize endosperm, a major force in sink strength, is in general not initiated until ∼12 DAP. Largely due to these spatial and temporal constraints of Ivr2 expression, it is unlikely to play any major role in the sink strength of the filial tissue, endosperm and embryo. Of the four CWI genes (Kim et al. 2000), Incw3 and Incw4 are putative CWI genes; their expression in various tissues is detectable only by RT-PCR, and neither is expressed in developing seeds. The Incw1 and Incw2 genes are considered to be the major genes and their expressions are readily detectable by Northern and Western blot analyses (Taliercio et al. 1999). Our objectives for this study are: (1) to elucidate a possible genetic basis of the residual low levels of CWI activity in the mn1 seed mutant, and (2) to describe quantitatively the overall patterns of expression of these two CWI genes in developing endosperms using more sensitive approaches, including real-time PCR (q-PCR).
Materials and methods
Plant material
Immature maize (Zea mays L.) kernels of Mn1 and the mn1 reference allele in the W22 inbred line were harvested at 8, 12, 20, 24 and 28 DAP. Plants were grown in the field during 2001 and were either self-pollinated or sib-pollinated. Where indicated, additional plants were grown in the greenhouse, under normal diurnal conditions. At the time of harvest, kernels were excised from the ear, taking care to include the pedicel region. Excised kernels were flash frozen in liquid nitrogen and stored at −80°C until further use.
Northern blot analyses
Kernel samples were harvested from field-grown material and included the entire kernel, pedicel and crown, and used for isolation of total RNA following the method of Wadsworth et al. (1988). RNA was glyoxalated and resolved on a 1.2% (w/v) agarose gel essentially according to Ausubel et al. (1993), followed by transfer to Nytran membrane (Schleicher & Schull, Keene, NH) and UV cross-linked. Blots were prehybridized at 65°C for 3 h in buffer consisting of 50 mM PIPES (pH 6.5), 0.1 M NaCl, 50 mM sodium phosphate buffer (pH 6.5), 1 mM EDTA and 5% (w/v) ultrapure SDS. Probes were prepared from the full-length cDNA clones and labeled using a random priming method (Prime-It RmT Random Primer Labeling Kit, Stratagene, LaJolla, CA, USA). Hybridization was performed in the same buffer with ∼3×106 counts ml−1 of [α-P32]dCTP-labeled probe, at 65°C overnight with constant shaking. Following hybridization, blots were rinsed twice in 6X SSC (1X SSC is 0.15 M NaCl and 0.015 M sodium citrate) solution supplemented with 5 mM sodium phosphate (pH 6.5), 5 mM EDTA (pH 8.0) and 5% (w/v) SDS, followed by two high-stringency washes in 0.2X SSC, 5 mM sodium phosphate (pH 6.5), 5 mM EDTA (pH 8.0) and 1% (w/v) SDS. All rinses were performed at 65°C for 20 min each. The blots were exposed to X-ray film, in between two intensifying screens, for 1–4 days depending on the transcript abundance. Transcript size was estimated to be 2.5–2.7 kb based on the position of the rRNA species visualized in the ethidium bromide-stained gel prior to blotting, and by comparison with the Incw2 transcript (∼2.2 kb).
Preparation of protein extracts and invertase activity assays
Protein extracts enriched for CWI protein were prepared using the basal (approximately the lower one third) sections of the developing kernels as described in Carlson et al. (2002). The basal sections of the frozen kernels were excised and pulverized under liquid nitrogen with a mortar and pestle. The powder was then resuspended in five volumes of 50 mM Tris–maleate (pH 7.0) buffer. The slurry was cleared by centrifugation for 20 min at 27,000×g. The pellets (containing the cell wall fraction) were resuspended in two volumes of extraction buffer supplemented with 1 M NaCl, vigorously agitated for 30 min at 4°C using an Eppendorf shaker, followed by centrifugation for 20 min at 27,000×g. The resulting supernatants, enriched for the CWI fractions, were desalted on MicroBio-Spin 6 chromatography columns (BioRad, Hercules, CA, USA) as per the manufacturer’s recommendations. Acid invertase activity was measured in a spectrophotometric assay as previously described (Tsai et al. 1970; Miller and Chourey 1992), using glucose as the standard in the Nelson’s assay for reducing sugars. Activity assays were performed in triplicate using at least two separate extractions for each sample. Protein concentrations were determined using the BioRad Protein Assay (BioRad), with BSA as the standard.
SDS-PAGE and western blots
Samples prepared for SDS-PAGE electrophoresis and blotting were immediately denatured in 2X SDS-PAGE sample buffer [0.125 M Tris–HCl, 4% (w/v) SDS, 20% (v/v) glycerol, 0.02% (w/v) BPB, pH 6.8] by boiling at 100°C for 5 min. Samples were subjected to electrophoresis immediately or stored at −20°C overnight. SDS-PAGE analyses were done using 7.5% (w/v) acrylamide Tris–HCl gels according to Laemmli (1970). Protein blotting to PVDF membrane (Immobilon-P, Millipore, Billerica, MA, USA) was performed in the BioRad Mini Trans-Blot apparatus, following the manufacturer’s instructions. As an additional blotting control, pre-stained standards were loaded along with the samples (Kaleidoscope Markers, BioRad). Western blots were developed using enhanced chemiluminescent substrate (Pierce Super Signal Substrate, Rockford, IL, USA). The primary polyclonal anti-INCW1 antiserum was used at a dilution of 1:1,000. Production and characterization of anti-INCW1 antiserum has previously been described (Carlson and Chourey 1999). Anti-mouse HRP-labeled secondary antiserum was used for INCW1 detection at a 1:12,500 dilution. As a control, polyclonal anti-UGPase antiserum was used as previously described (Carlson et al. 2002).
Immunolocalization of the CWI protein
Developing kernels at various developmental stages were harvested and immediately fixed in formalin acetic alcohol (FAA), dehydrated through a tertiary butyl alcohol (TBA) series, infiltrated in Paraplast plus paraffin (Fisher Scientific), embedded, sectioned and immunostained essentially following the protocol described previously (Cheng et al. 1996). Polyclonal antibodies of maize CWI were raised in mice (Carlson and Chourey 1999), and the immunogold signal was visualized using a commercial ZYMED kit (Zymed Laboratories Inc, San Francisco, CA, USA).
Reverse transcriptase PCR
The reverse transcription reactions were performed using SuperScript First-Strand Synthesis System (Invitrogen, Carlsbad, CA, USA) essentially according to the manufacturer’s protocol. Five microgram total RNA, 1 mM dNTP mix and 0.5 μg oligo(dT)12-18 primers were mixed in a final volume of 10 μl, incubated at 65°C for 5 min, and immediately chilled on ice. First-strand synthesis buffer was added to a final volume of 20 μl, containing 5 mM MgCl2, 10 mM DTT and RNaseH inhibitor, pre-incubated at 50°C for 2 min prior to addition of 1 μl (50 U μl−1) SuperScript II RT. The RT reactions were incubated for an additional 50 min at 50°C, and terminated at 70°C for 15 min. Finally, the RT reactions were treated with RNaseH at 37°C for 15 min and stored at −20°C. The first-strand cDNA was used within 2–3 days for PCR analyses. Gene-specific primers for RT-PCR and real-time quantitative PCR were designed from the selected gene sequences (Table 1). The PCR Opti-Prime system (Invitrogen) was used for standardizing the optimal pH, magnesium and DMSO concentration. Incw1 and Incw2 were amplified for 35 cycles of 1 min at 94°C for 45 s at 55°C and extension for 1 min at 72°C. Likewise, α -tubulin was amplified for 35 cycles of 1 min at 94°C, 1 min at 52°C, followed by extension for 1 min at 72°C. All the PCR runs were started with an initial step of 5 min at 94°C, and were allowed a final extension of amplification products for 10 min at 72°C. The PCR amplified products (Incw1, ∼650 bp; Incw2, ∼630 bp; and α -tubulin, ∼600 bp) were resolved on 1.2% (w/v) agarose gel and visualized after ethidium bromide staining.
Real-time quantitative PCR
Gene-specific mRNA quantification was performed by real-time quantitative PCR using the Chromo 4 CFD supported by Opticon Monitor Software version 2.03 (MJ Research, Alameda, CA, USA). SYBR Green chemistry was used for cDNA quantification (Applied Biosystems, Foster City, CA, USA). The PCR reaction contained 200 nm of both the forward and reverse gene-specific primers, 1 μl of the original 20 μl RT reaction, in addition to 1X SYBR Green PCR buffer, 3 mM MgCl2, 2 mM each of dATP, dCTP and dGTP and 4 mM dUTP, 1.25 U AmpliTaq Gold DNA polymerase and 0.5 U AmpErase UNG (uracil-N-glycosylase), in a final volume of 20 μl. The thermal cycling protocol entailed 50°C incubation for 2 min to activate AmpErase UNG activity, followed by AmpliTaq Gold DNA polymerase activation at 95°C for 10 min. The PCR amplification was carried out for 40 cycles with denaturation at 95°C for 15 s and primer annealing/extension at 60°C for 1 min. Optical data was acquired during the annealing step, and the fidelity of PCR reaction was monitored by melting curve analysis beginning at 55°C through 95°C, at 0.1°C s−1. Additionally, PCR products were also checked for purity by agarose gel electrophoresis. α -Tubulin was used as the endogenous reference gene for normalizing the Incw1 and Incw2 transcript profiles. The real time PCR data were calibrated relative to Incw1 or Incw2 levels in 12 DAP whole kernel, following the 2− ΔΔCt method for relative quantification (Livak and Schmittgen 2001). The efficiencies of PCR amplification for the reference and the target cDNA templates were validated using tenfold serial dilutions of the template RT reactions, and were found to be approximately similar under our amplification regime. The data is presented as average ± SD of three independently made RT preparations used for PCR run, each having three replicates.
Results
Expression analyses at the RNA level
Figure 1 shows an RNA blot for transcripts of Incw1 and Incw2 genes in developing kernels of lineage-related (W22 inbred line) Mn1 and mn1 genotypes at three developmental stages: 12, 24 and 28 DAP. An RNA sample from a BMS cell suspension line was included as a control because this sample is specific to Incw1 expression (Cheng et al. 1999). Loading controls for RNA levels were represented by ethidium bromide-stained rRNA bands and radiolabeled single copy transcripts sucrose synthase-3 encoded by Sus3, which does not show any temporal variability during kernel development (Carlson et al. 2001). As shown previously (Cheng et al. 1996), Mn1-encoded Incw2 transcripts in 12 DAP kernels were readily detectable in Mn1 but not in mn1 kernels (Cheng et al. 1996; Carlson and Chourey 1999). No Incw2 RNA was detectable in subsequent developmental stages in either genotype. In contrast, the same samples showed readily detectable levels of Incw1 transcripts in all three stages in both the Mn1 and mn1 genotypes. A control sample of RNA from maize suspension-cultured cells showed abundant steady-state levels of Incw1 transcript with no detectable Incw2 transcript. This control established gene-specificity of the cDNA probe, as the Incw2 probe does not show any cross-hybridization with the RNA sample from suspension-culture cells, where there is an abundance of Incw1 transcript (Fig. 1). Furthermore unlike Incw2 expression, there appeared to be no developmental variation at three stages, 12, 24 and 28 DAP, in steady-state levels of Incw1 RNA in wild type Mn1; a somewhat similar response was also seen in mn1 kernels which show reduced levels when compared with the Mn1.

Northern blot analyses of Mn1 and mn1 endosperm samples at various developmental stages. All lanes were loaded with 20 μg unless otherwise noted. For the Incw1 panel, RNA was loaded at 10 μg per lane, except for the cell suspension culture which was loaded at 5 μg level (but 10 μg level for the Incw2 panel). The transcript sizes were estimated to be 2.2 kb for Incw2 and Incw1 and 2.6 kb for Sus3, based on the position of the rRNA in the ethidium bromide-stained gel
RT-PCR analyses
Figure 2 shows a Southern blot of RT-PCR products representing Incw1 and Incw2 cDNA fragments of endosperm RNAs from two developmental stages, 12 and 28 DAP. Further, the endosperms were cut into two parts representing the lower one-third (including the BETC region) and upper two-thirds of an endosperm to test for possible spatial heterogeneity in expression. In contrast with our previous analyses (Cheng et al. 1996; Tailercio et al. 1999), the more sensitive method of RT-PCR used here showed both Incw1- and Incw2-specific products in the upper and lower regions of 12 DAP Mn1 endosperms. Hybridization analyses also revealed that whereas the Incw1 probe detected a single gene-specific fragment, the Incw2 visualized two fragments that were slightly different in size, 650 and 630 bp (the larger fragment of Incw1 showed weaker hybridization intensity with the Incw2 probe). Sequence analyses confirmed that the larger fragment was Incw1 and the smaller fragment was Incw2 (data not shown). At 28 DAP both the lower and the upper parts of the Mn1 endosperm showed the Incw1 fragment but not the Incw2 fragment. In the mn1 mutant, only the Incw1 product was seen in both the upper and the lower parts of the 12 DAP endosperm. As expected, the smaller cDNA fragment (Incw2) was missing in the mn1 mutant.

Southern blot analysis of RT-PCR products from upper and lower endosperms of Mn1 and mn1 at various developmental stages as shown. The lanes marked 1 and 2 contained RT-PCR products amplified using Incw1- and Incw2-gene-specific primers, respectively
q-PCR quantification of Incw1 and Incw2 RNAs
To make valid comparisons on the relative transcript levels of the two Incw genes, we quantified Incw1 and Incw2 expression using q-PCR, as shown in Fig. 3. These analyses revealed, for the first time, a high level of spatial and temporal regulation in the expression of these two genes. At 12 DAP, the lower Mn1 endosperm showed significantly higher abundance of Incw2 transcript relative to Incw1; the same region, however, showed greatly increased levels of Incw1 relative to the Incw2 at 20 DAP, and nearly equal levels at 28 DAP. The 20 DAP stage was also contrasted by nearly an order magnitude reduction in the levels of Incw2 RNA in the lower endosperm. Interestingly, a very high level of Incw1 expression was seen in both the upper and the lower parts of the endosperm in all three stages of development. Overall, both genes were expressed, though differentially, in the upper and lower regions of endosperm throughout development. In 12 DAP mn1 seed mutants, there was no detectable Incw2 expression, and the levels of Incw1 transcripts in the lower parts of the endosperm were higher than the upper region (3.8-, 3.0- and 2.0-fold at 12, 20 and 28 DAP, respectively). The Incw1 transcript levels in the mn1 mutant were relatively higher than the Mn seed throughout endosperm development.

a, b Real-time PCR determinations of the relative expression of Incw1 and Incw2 genes in a Mn1 and b mn1 in 12, 20 and 28 DAP endosperms. The relative transcript abundance was calibrated against 12 DAP Mn1 or mn1 endosperm (whole kernel), as indicated
Expression analyses at protein and enzyme activity levels
Figure 4 shows an immunoblot reacted with the polyclonal INCW1 antisera (Shanker et al. 1995) that recognizes both INCW1 and INCW2 forms (Carlson et al. 1999) estimated to be 68 kDa (p68) and 72 kDa (p72), respectively (Cheng et al. 1996). The Mn1 lower endosperm extracts at 12 DAP showed two polypeptides in highly unequal amounts; the most abundant protein was the 72 kDa INCW2 and the minor form was the INCW1 protein. The extracts from the upper endosperm region, 12 DAP, showed INCW2 and extremely low to undetectable levels of the INCW1 polypeptide. At later stages of development, 20, 24 and 28 DAP, the relative levels of INCW2 appeared to decrease gradually in the lower endosperm extracts, whereas the upper endosperm, in the storage region, showed only INCW1 and no detectable levels of INCW2 even at the maximal loading levels possible. The mn1 endosperm, both in the upper and in the lower regions at 12 DAP, showed a single p68 INCW1 polypeptide, consistent with our RNA level analyses that show no Incw2 transcripts in the mutant. A similar pattern was seen at 20 and 28 DAP stages .

Western blot analysis of CWI proteins in Mn1 and mn1 endosperm extracts as shown. Amounts of total protein loaded were (for lower and upper sections, respectively), Mn1 at 12 DAP: 2.5 and 5.0 μg; Mn1 at 20–28 DAP: 5.0 and 10.0 μg; and mn1 at 12 and 28 DAP: 10 and 20 μg per lane
The qualitative data from Western blots were approximately paralleled in enzyme activity estimates in both Mn1 and mn1 endosperms (Table 2). Specifically, a great bulk of enzyme activity was seen in extracts from the lower part, and relatively very little (5.65% of the lower) was seen in the upper storage region of the 12 DAP Mn1 endosperm. The temporal reductions at 20 and 28 DAP were similar to those reported previously (Cheng et al. 1996); however, in contrast to our previous results which showed no activity in upper regions, some very low level CWI activity was detected in the upper regions at both 20 and 28 DAP stages. It is significant to note that despite the highest levels of Incw1 RNA in both the upper and the lower parts of the Mn1 endosperm at 20 DAP (Fig. 3), the overall levels of total CWI activity (Table 2) and the two corresponding proteins (Fig. 4) were greatly reduced as compared to those seen at 12 DAP stage. Very little or no change in CWI activity was seen in the mn1 mutant throughout the duration of 12–28 DAP; except the upper region which at 28 DAP showed statistically significantly higher levels than the two early stages.
Immunolocalization of the CWI protein
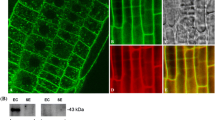
The same INCW1 antibodies as described above detected a strong immuno-reactive signal in the BETC region of 20 DAP Mn1 longitudinal sections (Fig. 5), as shown previously for 12 DAP kernels (Cheng et al. 1996). No such signal was seen in the mn1 mutant (Fig. 5) or with a pre-immune serum in either genotype (data not shown). There was also no detectable signal in the upper regions of the 20 DAP endosperm; apparently, the CWI enzyme activity and the protein seen in Western blots was below the detection limits of this assay.

a–c Immunolocalization of CWI protein in 20 DAP Mn1 and 16 DAP mn1 using INCW1 antibodies (primary antibody used at 1:2,000 dilution). a Mn1, 20 DAP, 40×; b Mn1, 20 DAP, 200× ; c mn1, 16 DAP, 40×
Discussion
Sugar metabolism and its control are critical to all parts of the plants, including developing seeds—a major site of sugar utilization. As discussed previously, the CWIs play a pivotal role in establishing sink strength through irreversible cleavage of sucrose in an apoplast, a major entry point for metabolizable sugars into a cell for both downstream signaling and metabolic functions. Our rationale for subdividing the developing endosperms into lower and upper parts (approximately, one-third and two-third ratio, respectively) is based on our previous observations that bulk of the CWI protein is in the BETC layer and the surrounding cells in the basal region whereas the upper part constitutes the storage cells metabolizing sucrose into storage compounds. The studies here have yielded two major observations on the nature of genetic control and regulation of CWIs in the developing endosperm of maize. Briefly, we show that: (1) the residual CWI activity in the mn1 mutant was due to the Incw1 gene, which is previously shown to be expressed in many tissues of the plant (Taliercio et al. 1999) and (2) the two previously characterized orthologes, Incw1 and Incw2, encoding similar proteins (Taliercio et al. 1999), were expressed with distinctive yet overlapping spatial and temporal specificities in the developing endosperm.
The residual low level of CWI activity in the standard mn1 mutant and several of its allelic EMS mutants has been known for the past several years (Miller and Chourey 1992; Cheng et al. 1996) but a genetic basis of this enzyme has remained unknown until now. As is often the case with many endosperm mutants in maize, the mutational inactivation of a certain gene invariably uncovers another duplicate gene. We describe a second gene, an ortholog that encoded CWI in the developing endosperm with the mutational loss of ∼99% of the enzyme activity in the mn1 mutation. Collective data based on Northern blots (Fig. 1), RT-PCR gels (Fig. 2), sequence analysis of the cloned fragment (data not shown) and Western blot analyses (Fig. 4), establish that the low level CWI activity in the mutant is encoded by the Incw1 gene, the only CWI gene expressed in the mn1 endosperm. The Incw1 gene has been cloned and characterized previously (Shanker et al. 1995; Taliercio et al. 1999); however, its expression in the developing endosperm has remained elusive. The q-PCR data (Fig. 3) show the highest levels of Incw1 transcripts in 28 DAP mn1 relative to both Incw1 and Incw2 at all three stages of the Mn1; yet, the levels of the CWI protein and enzyme activity in the mutant were the lowest relative to Mn1. Similar results showing increased steady-state levels of Incw1 RNA but no associated increases in the levels of the INCW1 protein were reported previously in sugar-starved cell suspension cultured cells of maize (Cheng et al. 1999). The cell culture study suggested that the 3′ untranslated region of the Incw1 gene acts as a regulatory sensor of carbon starvation and may provide a link between sink metabolism and cellular translation in plants (Cheng et al. 1999). A molecular basis of the post-transcriptional control in the regulation of the Incw1 gene in the developing endosperm remains to be analyzed. An alternate possibility on the greatly diminished levels of enzyme activity in the mn1 mutant may be due to increased levels of an invertase inhibitor protein recently described in developing maize endosperm (Bate et al. 2004; McLaughlin and Boyer 2004). The inhibitor protein is postulated to act through protein–protein interactions between CWI and the invertase inhibitor (Greiner et al. 1998).
As for the Mn1 endosperm, the q-PCR results (Fig. 3) suggest a far more complex picture in spatial and temporal heterogeneity of expression of the two CWI genes than our previous results based on the expression of only the single gene, Incw2 (Cheng et al. 1996). As expected, the 12 DAP stage was marked by high levels of both Incw2 RNA (Figs. 1, 3) and protein/enzyme activity in the sugar unloading zone, the BETC region, of the endosperm (Fig. 4, Table 2). A key role of CWI at this early developmental stage is in the maintenance of cell division through hexose signaling and its provision in metabolism, as discussed previously. The irreversible hydrolysis of sucrose to hexoses may also lead to increased osmotic pressure and an increased pull of water into the endosperm. Increased turgor pressure is essential for cell expansion. Not surprisingly, the mn1 mutant is associated with several drastic changes at both phenotypic (Miller and Chourey 1992; Cheng et al. 1996) and cellular levels (Vilhar et al. 2002). Largely due to the severity of these phenotypic changes, we suggest that the Mn1-encoded INCW2 protein must control the metabolic flux of sugars entering a developing endosperm.
The q-PCR results also show greater abundance of Incw1 RNA than Incw2 at the storage phase, 20–28 DAP, of endosperm development. Despite the greatly elevated levels of the Incw1 transcript, very little or no change was seen in the levels of the INCW1 polypeptide, which indicated that either the Incw1 RNA was not translated and/or the protein was preferentially unstable in the lower endosperm (the BETC region). Conversely, the greater relative abundance of INCW2 than INCW1, but not the respective transcripts, indicated a much slower turnover rate of the INCW2 protein in the lower endosperm region. The upper endosperm showed no detectable levels of the INCW2 protein despite low levels of Incw2 RNA (relative to the Incw1). Clearly, there was a cell-specific intergenic cross-talk in the regulation of the two CWI genes in the ultimate expression at the protein level; Incw2 was partly epistatic to Incw1 in the lower endosperm and the reverse was true in the upper endosperm. A similar post-transcriptional or translational control is also described for the two sucrose synthase orthologs in developing embryos in maize (Chourey and Taliercio 1994).
A strong immuno-signal for CWI protein in the BETC layer in 20 DAP Mn1 kernel sections, but not in the mn1 mutant (Fig. 5), indicates that sucrose hydrolysis in the basal part of the endosperm remained a critical function, as is the case of the 12 DAP kernels (Cheng et al. 1996). Although not detectable by immunolocalization, CWI activity was also seen in the upper region, albeit greatly reduced when compared with the lower endosperm, throughout the duration of seed development in both Mn1 and mn1 endosperms. Collectively, these results show that although symplastic sucrose transport throughout the developing endosperm is not ruled out, the apoplastic cleavage of sucrose and the entrance of hexose as the primary sugar in endosperm metabolism is also a strong possibility. Whether the apoplastic or the symplastic sucrose has differential metabolic fates in endosperm cells is unknown.
Abbreviations
- BETC:
-
Basal endosperm transfer cells
- CWI:
-
Cell wall invertase
- DAP:
-
Days after pollination
- INCW:
-
Invertase cell wall
- mn :
-
Miniature
References
Anderson MN, Asch F, Wu Y, Jensen CR, Naested H, Mogensen VO, Koch KE (2002) Soluble invertase expression is an early target of drought stress during the critical, abortion-sensitive phase of young ovary development in maize. Plant Physiol 130:591–604
Ausubel FM, Brent R, Kingston RE, Moore DD, Seidman JG, Smith JA, Struhl K (1993) Current protocols in molecular biology. Wiley, New York
Bate NJ, Niu X, Wang Y, Reimann KS, Helentjaris TG (2004) An invertase inhibitor from maize localizes to the embryo surrounding region during early kernel development. Plant Physiol 134:246–254
Carlson SC, Chourey PS (1999) A re-evaluation of the relative roles of two invertases, INCW2 and IVR1, in developing maize kernels and other tissues. Plant Physiol 121:1025–1035
Carlson SJ, Chourey PS, Helentjaris T, Datta R (2002) Gene expression studies on developing kernels of maize sucrose synthase (SuSy) mutants show evidence for a third SuSy gene. Plant Mol Biol 49:15–29
Cheng WH, Chourey PS (1999) Genetic evidence that invertase mediated release of hexoses is critical for appropriate carbon partitioning and normal seed development in maize. Theor Appl Genet 98:485–495
Cheng WH, Taliercio EW, Chourey PS (1996) The Miniature1 seed locus of maize encodes a cell wall invertase required for normal development of endosperm and maternal cells in the pedicel. Plant Cell 8:971–983
Cheng WH, Taliercio EW, Chourey PS (1999) Sugars modulate an unusual mode of control of the cell-wall invertase gene (Incw1) through its 3′ untranslated region in a cell suspension culture of maize. Proc Natl Acad Sci USA 96:10512–10517
Cho JI, Lee SK, Ko S, Kim HK, Jun SH, Lee YH, Bhoo SH, Lee KW, An G, Hahn TR, Jeon JS (2005) Molecular cloning and expression analysis of the cell wall invertase gene family in rice (Oryza sativa L.) Plant Cell Rep 10:1–22
Chourey PS, Taliercio EW (1994) Epistatic interaction and functional compensation between the two tissue- and cell-specific sucrose synthase genes in maize. Proc Natl Acad Sci USA 91:7917–7921
Godt DE, Roitsch T (1997) Regulation and tissue specific distribution of mRNA for three extracellular invertase isozymes in tomato suggests an important function in establishing and maintaining sink metabolism. Plant Physiol 115:273–282
Greiner S, Krausgrill S, Rausch T (1998) Cloning of a tobacco apoplasmic invertase inhibitor. Proof of function of the recombinant protein and expression analysis during plant development. Plant Physiol 116:733–742
Heyer AG, Raap M, Schroeer B, Marty B, Willmitzer L (2004) Cell wall invertase expression at the apical meristem alters floral, architectural, and reproductive traits in Arabidopsis thaliana. Plant J 39:161–169
Hirose T, Takano M, Terao T (2002) Cell wall invertase in developing rice caryopsis: molecular cloning of OsCIN1 and analysis of its expression in relation to its role in grain filling. Plant Cell Physiol 43:452–459
Kim JY, Mahe A, Guy S, Brangeon J, Roche O, Chourey PS, Prioul JL (2000) Characterization of two members of the maize gene family, Incw3 and Incw4, encoding cell-wall invertases. Gene 245:89–102
Laemmli UK (1970) Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature 227:680–685
Lara MEB, Garcia MCG, Fatima T, Ehne R, Lee TK, Proels R, Tanner W, Roitsch T (2004) Extracellular invertase is an essential component of cytokinin-mediated delay of senescence. Plant Cell 16:1276–1287
Livak KJ, Schmittgen TD (2001) Analysis of relative gene expression data using real-time quantitative PCR and the 2\Delta\Delta ct method. Methods 25:402–408
McLaughlin JE, Boyer JS (2004) Sugar-responsive gene expression, invertase activity, and senescence in aborting maize ovaries at low water potentials. Ann Bot (Lond) 94:675–689
Miller ME, Chourey PS (1992) The maize invertase-deficient miniature-1 seed mutation is associated with aberrant pedicel and endosperm development. Plant Cell 4:297–305
Ohto M, Fisher RL, Goldberg RB, Harada JJ (2005) Control of seed mass by APETALA2. Proc Natl Acad Sci USA 102:3123–3128
Shanker S, Salazar RW, Taliercio EW, Chourey PS (1995) Cloning and characterization of full-length cDNA encoding cell-wall invertase from maize. Plant Physiol 108:873–874
Sonnewald U, Hajirezaei MR, Kossmann J, Heyer A, Trethewey RN, Willmitzer L (1997) Increased potato tuber size resulting from apoplastic expression of a yeast invertase. Nat Biotechnol 15:794–797
Sreenivasulu N, Altschmied L, Radchuk V, Gubatz S, Wobus U, Weschke W (2004) Transcript profiles and deduced changes of metabolic pathways in maternal and filial tissues of developing barley grains. Plant J 37:539–553
Sturm A (1999) Invertases: primary structures, functions, and roles in plant development and sucrose partitioning. Plant Physiol 121:1–8
Taliercio EW, Kim JY, Mahé A, Shanker S, Choi J, Cheng WH, Prioul JL, Chourey PS (1999) Isolation, characterization and expression analyses of two cell wall invertase genes in maize. J Plant Physiol 155:197–204
Tang GQ, Luscher M, Sturm A (1999) Antisense repression of vacuolar and cell wall invertase in transgenic carrot alters early plant development and sucrose partitioning. Plant Cell 11:177–189
Tsai CY, Salamini F, Nelson OE (1970) Enzymes of carbohydrate metabolism in the developing endosperm of maize. Plant Physiol 46:299–306
Vilhar B, Kladnik A, Blejec A, Chourey PS, Dermastia M (2002) Cytometrical evidence that the loss of seed weight in the miniature1 seed mutant of maize is associated with reduced mitotic activity in the developing endosperm. Plant Physiol 129:23–30
Wadsworth GJ, Redinbaugh MG, Scandalios JG (1988) A procedure for the small-scale isolation of plant RNA suitable for RNA-blot analysis. Anal Biochem 172:279–283
Weber H, Buchner P, Borisjuk L, Wobus U (1996) Sucrose metabolism during cotyledon development of Vicia faba L. is controlled by the concerted action of both sucrose-phosphate synthase and sucrose synthase: expression patterns, metabolic regulation and implications for seed development. Plant J 9:841–850
Weber H, Borisjuk L, Heim U, Sauer N, Wobus U (1997) A role for sugar transporters during seed development: molecular characterization of a hexose and a sucrose carrier in fava bean seeds. Plant Cell 9:895–908
Weschke W, Panitz R, Gubatz S, Wang Q, Radchuk R, Weber H, Wobus U (2003) The role of invertases and hexose transporters in controlling sugar ratios in maternal and filial tissues of barley caryopses during early development. Plant J 33:395–411
Xu J, Avigne WT, McCarty DR, Koch KE (1996) A similar dichotomy of sugar modulation and developmental expression affects both paths of sucrose metabolism: evidence from a maize invertase gene family. Plant Cell 8:1209–1220
Acknowledgements
Mention of trade names or commercial products in this publication is solely for the purpose of providing specific information and does not imply recommendation or endorsement by the U.S. Department of Agriculture. We thank Drs. S. LeClere, D. R. Pring and E. W. Taliercio for critical reading of the manuscript. This was a cooperative investigation of the U.S. Department of Agriculture—Agricultural Research Service and the Institute of Food and Agricultural Science, University of Florida. This paper is Florida Agricultural Experiment Station Journal Series Number R-10959.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Chourey, P.S., Jain, M., Li, QB. et al. Genetic control of cell wall invertases in developing endosperm of maize. Planta 223, 159–167 (2006). https://doi.org/10.1007/s00425-005-0039-5
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00425-005-0039-5