
Abstract
A novel WRKY-like transcription factor was isolated from a screen for weakly expressed mRNAs in ovules in the self-incompatible wild potato species Solanum chacoense Bitt. This protein, named ScWRKY1, consisted of 525 amino acids and can be classified as a WRKY group-I member, having two WRKY domains. It is expressed at low levels in stems, roots, and petals, and expressed at much higher levels in leaves. Interestingly, although barely detectable in developing seeds, it is strongly and transiently expressed in fertilized ovules bearing late torpedo-staged embryos, suggesting a specific role during embryogenesis.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
During embryogenesis, the architecture of the mature plant is established through a series of developmental steps during which the zygote undergoes a complex series of morphological and cellular changes that results in the formation of a mature embryo comprised of an embryonic axis with shoot and root poles, and cotyledons. These transitions are orchestrated by numerous master regulatory genes, which comprise mainly transcription factors and genes involved in cellular signaling (Raghavan 1997; Chaudhury et al. 2001). Transcription factors of the WRKY-type family have, so far, only been isolated from plants, and with the completion of the Arabidopsis genome about 74 proteins containing the WRKY motif have been predicted (Dong et al. 2003; Kalde et al. 2003). WRKY proteins possess a characteristic and highly conserved 60-amino-acid domain that includes the WRKY motif, hence the family name, and a novel zinc-finger-like motif. Although most WRKY genes have been shown to be involved in the plant defense response (Eulgem et al. 2000; Dong et al. 2003; Kalde et al. 2003), many WRKY genes have also been found to be expressed during various developmental or physiological processes including leaf senescence (Hinderhofer and Zentgraf 2001), dormancy and drought tolerance (Pnueli et al. 2002), thermal hysteresis in the bittersweet nightshade, Solanum dulcamara (Huang and Duman 2002), and in orchard grass somatic embryogenesis (Alexandrova and Conger 2002). The TRANSPARENT TESTA GLABRA 2 mutant of Arabidopsis shows disruptions in trichome development and to tannin and mucilage production in the seed coat. It also encodes a WRKY protein of the group I with two WRKY domains, and is strongly expressed in trichomes throughout their development, as well as in the endothelium of developing seeds and later in other layers of the seed coat (Johnson et al. 2002). In this paper we present the preliminary characterization of a WRKY transcription factor, isolated from an ovule library made from tissues collected from 7 to 17 days post-pollination, bearing embryos from the globular to the bent-cotyledon/walking stick stages in the self-incompatible wild potato species Solanum chacoense.
Materials and methods
Plant material
The diploid (2n=2x=24) Solanum chacoense Bitt. self-incompatible genotypes used include line PI 458314 (S11 and S12 self-incompatibility alleles) and line PI 230582 (S13 and S14 self-incompatibility alleles) originally obtained from the Potato Introduction Station (Sturgeon Bay, WI, USA). Plants were grown on soil in a greenhouse with an average of 14 h of natural light per day supplemented with light from sodium lamps.
cDNA library construction and selection of weakly expressed genes
Poly(A)+mRNA (5 μg) was isolated from ovules hand-dissected from ovaries 7 to 17 days after pollination, and used to make a cDNA library in the Uni-ZAP vector following the manufacturer’s instruction (Stratagene). The cDNA phage library was converted to a plasmid cDNA library by mass excision. Plasmid cDNA library colonies (3,840) were transferred to positively charged nylon membranes and denatured in 0.5 N NaOH and 1.5 M NaCl. The resulting membranes were hybridized with a probe derived from α-[P32]dATP random-labeled cDNA leftovers (library target tissues) obtained during the library construction and were exposed at −85°C with one intensifying screen on Kodak Biomax MR film (Interscience, Markham, ONT, Canada).
The cDNA probes produced corresponded exactly with the abundance of each mRNA species found in the tissue tested. Rare mRNA species are under-represented and thus made poor probes. This enabled the selection of weakly expressed genes. Of the 3,840 colonies plated, only the lowest fifth in expression level (as determined through densitometric scans and through inspection by eye) were selected for sequencing on an automated sequencer (ABI Prism 310).
Isolation and gel blot analysis of RNA and DNA
Total RNA was isolated as described previously (Jones et al. 1985). RNA concentration was determined by measuring its absorbance at 260 nm and verified by agarose gel electrophoresis and ethidium bromide staining. Equal loading of total RNA on RNA gel blots was verified with an S. chacoense 18S RNA probe. Genomic DNA was isolated via a modified CTAB extraction method (Reiter et al. 1992). DNA gel blot analysis, including restriction, electrophoresis, and capillary transfer to a positively charged nylon membrane, (Hybond N+; Amersham Pharmacia Biotech) were performed as described by Sambrook et al. (1989). Membranes were hybridized at 42°C for 16–24 h in 50% deionized formamide, 5× Denhardt’s solution, 0.5% SDS, 200 μg ml−1 denatured salmon sperm DNA, and 6× SSC. Following hybridization, the membranes were washed twice for 30 min in 2× SSC/0.1% SDS at 35°C, followed by washes in 1× SSC/0.1% SDS at 45°C and 55°C for 30 min, and finally once with 0.1× SSC/0.1% SDS at 55°C for 10 min (1× SSC is 0.15 M NaCl, 0.015 M sodium citrate, pH 7.0). RNA gel blot analyses were performed as described in Sambrook et al. (1989), following the formaldehyde denaturing protocol. RNAs were capillary-transferred and cross-linked (120 mJ cm−2) to Hybond N+ nylon membranes. Membranes were hybridized under high-stringency conditions at 45°C in the same solution as for DNA gel blots. The membranes were washed as mentioned for the DNA gel blots. Probes for both DNA and RNA gel blot analyses were synthesized from random-labeled isolated DNA inserts (Roche Diagnostic, Laval, Qc, Canada) with α-[32P]dATP or α-[32P]dCTP (ICN Biochemicals, Irvine, CA, USA). The membranes were exposed at −85°C with one intensifying screen on Kodak Biomax MR film (Interscience).
Results and discussion
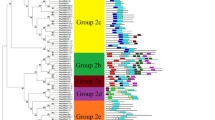
In order to isolate master regulatory genes expressed during embryo and seed development we used a negative selection screen based on the virtual subtraction method of Li and Thomas (1998) to isolate genes expressed at low levels in ovule tissues. Using a modified screening procedure with probes derived from the target-tissue cDNA library, we were able to isolate numerous proteins involved in transcriptional regulation and cellular signaling in our gene pool (see Materials and methods). One of the isolated cDNas corresponded to a WRKY-type transcription factor. The Solanum chacoense WRKY1 cDNA clone (ScWRKY1) is 1,801 bp (excluding the poly(A)-tail) with a very short 5′ untranslated leader of 8 nucleotides (nt), and a 3′ untranslated region (UTR) of 215 nt. Although the cDNA is incomplete (the length of the hybridizing mRNA is estimated at 2.06 kb on gel), an in-frame stop codon is found immediately upstream of the first ATG, indicating that the ScWRKY1-encoded protein is complete. The ScWRKY1 cDNA is predicted to encode a 525-amino-acid polypeptide of 58.3 kDa with a predicted isoelectric point (pI) of 8.4. Two WRKY domains are present on ScWRKY1. The first WRKY domain is centered around amino acid 201 and the second domain is centered around amino acid 366 (Fig. 1, inset). This conserved 60-amino-acid WRKY domain is suggested to be the DNA-binding domain that interacts with the core DNA sequence motif TGAC, called the W-box (Rushton and Somssich 1998). The ScWRKY1 protein can be classified as a group-I WRKY protein, having two WRKY domains and a C-X4-C-X22-H-X1-H zinc-finger-like motif, positioned 17 amino acids downstream of the first WRKY motif, and a C-X4-C-X23-H-X1-H zinc-finger-like motif, positioned 16 amino acids downstream of the second WRKY motif (Eulgem et al. 2000). Three serine-rich islands comprising 24% of the first 200 amino acids and a glutamine-rich island are also found in the N-terminal region immediately upstream of the first WRKY domain, which is also characteristic of many WRKY proteins (Fig. 1, inset). Overall sequence alignments with the closest ScWRKY1 homologues of comparable length show that ScWRKY1 shared 88% amino acid sequence identity (92% similarity) with the NtWRKY1 gene from tobacco (accession number AB022693); 81% amino acid sequence identity (86% similarity) with the NtWRKY6 gene from tobacco (Maeo et al. 2001); 52% amino acid sequence identity (63% similarity) with the parsley PcWRKY1 protein (Rushton et al. 1996); 51% amino acid sequence identity (63% similarity) with the WRKY1 protein from Pimpinella brachycarpa (accession number AF080595); 50% amino acid sequence identity (61% similarity) with the SPF1 DNA-binding protein from sweet potato (Ishiguro and Nakamura 1994); and 49% amino acid sequence identity (61% similarity) with the closest Arabidopsis homologue AtWRKY33 gene (accession number Q8S8P5). All of the above-mentioned WRKY proteins also fall into the group-I family. Figure 1 shows an alignment that includes the region covered by the two WRKY domains from the WRKY proteins most closely related to ScWRKY1. Although the WRKY domains are strongly conserved among all these proteins, the sequence between these domains is quite divergent, mainly in the region immediately following the first WRKY domain. This divergence outside the WRKY domains is also reflected by the overall sequence similarity levels mentioned before.

Alignment of the Solanum chacoense ScWRKY1 deduced protein sequence with closely related WRKY proteins covering the two WRKY domains. The deduced amino acid sequence of ScWRKY1 was aligned (with MacVector 7.2) with those of the following WRKY proteins: tobacco (Nicotiana tabacum) NtWRKY1 (accession number AB022693); N. tabacum NtWRKY2 (accession number AB020590); N. tabacum NtWRKY6 (Maeo et al. 2001); parsley (Petrosilenum crispum) PcWRKY1 (Rushton et al. 1996); Pimpinella brachycarpa PbWRKY1 (accession number AF080595); sweet potato (Ipomea batata) SPF1 (Ishiguro and Nakamura 1994); Arabidopsis thaliana AtWRKY33 (accession number Q8S8P5); rice (Oryza sativa) OsABF1(accession number NM190410); Capsella rubella CrWRKY33 (accession number AY314971). A consensus sequence is shown underneath the alignment. Bottom right Schematic of the overall structure of the ScWRKY1 gene, indicating the number and relative position of the WRKY domains, and the serine-rich (S) and glutamine-rich (Q) islands
Tissue-specific expression of ScWRKY1 was determined using RNA extracted from various tissues of S. chacoense. Figure 2a shows that ScWRKY1 is expressed in leaves, and to lower levels in petals, stems and roots. Interestingly, ScWRKY1 is also strongly expressed but only transiently in ovules bearing mainly late-torpedo-stage embryos, 16 days after fertilization (Fig. 2a,b). Few WRKY target genes have been characterized to date, but from the above-mentioned closest homologues, the sweet potato SFB1 and the parsley PcWRKY1 have been shown to regulate the expression of a pathogenesis-related gene (PR10) and a β-amylase gene, respectively (Ishiguro and Nakamura 1994; Eulgem et al. 1999). In order to determine if the expression of ScWRKY1 could also be involved in the expression of these genes, we used a potato PR10a cDNA, also known to contain a W-box in its promoter (Matton and Brisson 1989; Matton et al. 1993), and a potato β-amylase cDNA fragment (isolated from our gene pool). As shown in Fig. 2a, the potato β-amylase gene is only expressed in pollen and anthers, while the PR10a gene is only detectable in non-stressed leaves although it is strongly induced following elicitor treatments or pathogen infections in both leaves and tubers (Matton and Brisson 1989; Constabel and Brisson 1992). The constitutive expression of the ScWRKY1 gene in ovules 16 days after pollination, in leaves, petals, roots or stems is clearly quite different than for the two putative targets of ScWRKY1 homologues, suggesting that it is probably not involved in the regulation of these genes. To determine the ScWRKY1 gene copy number, a DNA gel blot analysis was performed of genomic DNA with various restriction enzymes (Fig. 2c). The restriction enzyme Eco RI cuts 4 times in the cDNA, between positions 44 and 456, Hind III cuts once at position 586, Xba I cuts once at position 1673, while Eco RV does not cut in the ScWRKY1 cDNA sequence. Since we have not determined the genomic organization of the ScWRKY1 gene, we cannot exclude the presence of restriction sites in putative introns, but the overall digestion pattern suggests that the ScWRKY1 gene is most probably present as a single-copy gene or, at the most, by two related copies in the S. chacoense diploid genome.

RNA expression analysis of ScWRKY1 transcript levels in mature tissues and in fertilized ovules. Ten μg of total RNA from various tissues was probed with the ScWRKY1 cDNA insert, stripped and re-probed with S. tuberosum PR10a and β-amylase cDNAs, as well as with a PCR product corresponding to the 18S rRNA gene as a control. b Hand-dissected embryos from compatibly pollinated plants, from 15 to 17 days after pollination. c DNA gel blot analysis of the ScWRKY1 gene. Genomic DNA (10 μg) isolated from S. chacoense leaves was digested with Eco RI, Eco RV, Hind III and Xba I restriction enzymes and probed with the complete ScWRKY1 cDNA insert. Molecular weights of the fragments as determined from the DNA MW ladder appear on the left
Although most WRKY genes have been shown to be involved in the plant defense response ( Eulgem et al. 2000; Dong et al. 2003; Kalde et al. 2003), WRKY proteins have also recently been found to be involved in plant developmental processes such as somatic embryogenesis (Alexandrova and Conger 2002) or trichome development (Johnson et al. 2002). These two examples, combined with the expression pattern observed for ScWRKY1 in torpedo-stage embryos, suggest that transcription factors of the WRKY family could be involved in crucial events during embryogenesis and seed development in plants. Further clarification of WRKY function in processes other than defense responses awaits the characterization of new mutant phenotypes involving WRKY proteins.
References
Alexandrova KS, Conger BV (2002) Isolation of two somatic embryogenesis-related genes from orchardgrass (Dactylis glomerata). Plant Sci 162:301–307
Chaudhury AM, Koltunow A, Payne T, Luo M, Tucker MR, Dennis ES, Peacock WJ (2001) Control of early seed development. Annu Rev Cell Dev Biol 17:677–699
Constabel CP, Brisson N (1992) The defense-related STH-2 gene product of potato shows race-specific accumulation after inoculation with low concentrations of Phytophthora infestans zoospores. Planta 188:289–295
Dong J, Chen C, Chen Z (2003) Expression profiles of the Arabidopsis WRKY gene superfamily during plant defense response. Plant Mol Biol 51:21–37
Eulgem T, Rushton PJ, Schmelzer E, Hahlbrock K, Somssich IE (1999) Early nuclear events in plant defence signalling: rapid gene activation by WRKY transcription factors. EMBO J 18:4689–4699
Eulgem T, Rushton PJ, Robatzek S, Somssich IE (2000) The WRKY superfamily of plant transcription factors. Trends Plant Sci 5:199–206
Hinderhofer K, Zentgraf U (2001) Identification of a transcription factor specifically expressed at the onset of leaf senescence. Planta 213:469–473
Huang T, Duman JG (2002) Cloning and characterization of a thermal hysteresis (antifreeze) protein with DNA-binding activity from winter bittersweet nightshade, Solanum dulcamara. Plant Mol Biol 48:339–350
Ishiguro S, Nakamura K (1994) Characterization of a cDNA encoding a novel DNA-binding protein, SPF1, that recognizes SP8 sequences in the 5′ upstream regions of genes coding for sporamin and beta-amylase from sweet potato. Mol Gen Genet 244:563–571
Johnson CS, Kolevski B, Smyth DR (2002) TRANSPARENT TESTA GLABRA 2, a trichome and seed coat development gene of Arabidopsis, encodes a WRKY transcription factor. Plant Cell 14:1359–1375
Jones JDG, Dunsmuir P, Bedbrook J (1985) High level expression of introduced chimeric genes in regenerated transformed plants. EMBO J 4:2411–2418
Kalde M, Barth M, Somssich IE, Lippok B (2003) Members of the Arabidopsis WRKY group III transcription factors are part of different plant defense signaling pathways. Mol Plant Microbe Interact 16:295–305
Li Z, Thomas TL (1998) PEI1, an embryo-specific zinc finger protein gene required for heart-stage embryo formation in Arabidopsis. Plant Cell 10:383–398
Maeo K, Hayashi S, Kojima-Suzuki H, Morikami A, Nakamura K (2001) Role of conserved residues of the WRKY domain in the DNA-binding of tobacco WRKY family proteins. Biosci Biotechnol Biochem 65:2428–2436
Matton DP, Brisson N (1989) Cloning, expression, and sequence conservation of pathogenesis-related gene transcripts of potato. Mol Plant Microbe Interact 2:325–331
Matton DP, Prescott G, Bertrand C, Camirand A, Brisson N (1993) Identification of cis-acting elements involved in the regulation of the pathogenesis-related gene STH-2 in potato. Plant Mol Biol 22:279–291
Pnueli L, Hallak-Herr E, Rozenberg M, Cohen M, Goloubinoff P, Kaplan A, Mittler R (2002) Molecular and biochemical mechanisms associated with dormancy and drought tolerance in the desert legume Retama raetam. Plant J 31:319–330
Raghavan V (1997) Molecular embryology of flowering plants. Cambridge University Press, Cambridge, UK
Reiter RS, Young RM, Scolnik PA (1992) Genetic linkage of the Arabidopsis genome: methods for mapping with recombinant inbreds and random amplified polymorphic DNAs (RAPDs). In: Koncz C, Chua N-H, Schell J (eds) Methods in Arabidopsis research. World Scientific Publishing, Singapore, pp 170–190
Rushton PJ, Somssich IE (1998) Transcriptional control of plant genes responsive to pathogens. Curr Opin Plant Biol 1:311–315
Rushton PJ, Torres JT, Parniske M, Wernert P, Hahlbrock K, Somssich IE (1996) Interaction of elicitor-induced DNA-binding proteins with elicitor response elements in the promoters of parsley PR1 genes. EMBO J 15:5690–5700
Sambrook J, Fritsch EF, Maniatis T (1989) Molecular cloning: a laboratory manual. Cold Spring Harbor Laboratory Press, Cold Spring Harbor, NY
Acknowledgements
This work was supported by the Natural Sciences and Engineering Research Council of Canada (NSERC) and by Le Fonds pour la Formation des Chercheurs et l’Aide à la Recherce (FCAR-FQRNT, Québec). We thank Sier-Ching Chantha and Martin O’Brien for providing pictures of hand-dissected embryos. We also thank Gabriel Téodorescu for plant care and maintenance.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Lagacé, M., Matton, D.P. Characterization of a WRKY transcription factor expressed in late torpedo-stage embryos of Solanum chacoense . Planta 219, 185–189 (2004). https://doi.org/10.1007/s00425-004-1253-2
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00425-004-1253-2