
Abstract
A protein from Arabidopsis thaliana (L.) Heynh. showing homology to animal proteins of the NaPi-1 family, involved in the transport of inorganic phosphate, chloride, glutamate and sialic acid, has been characterized. This protein, named ANTR2 (for anion transporters) was shown by chloroplast subfractionation to be localized to the plastid inner envelope in both A. thaliana and Spinacia oleracea (L.). Immunolocalization revealed that ANTR2 was expressed in the leaf mesophyll cells as well as in the developing embryo at the upturned-U stage. Five additional homologues of ANTR2 are found in the Arabidopsis genome, of which one was shown by green fluorescent protein (GFP) fusion to be also located in the chloroplast. All ANTR proteins share homology to the animal NaPi-1 family, as well as to other organic-anion transporters that are members of the Anion:Cation Symporter (ACS) family, and share the main features of transporters from this family, including the presence of 12 putative transmembrane domains and of a 7-amino acid motif in the fourth putative transmembrane domain. ANTR2 thus represent a novel protein of the plastid inner envelope that is likely to be involved in anion transport.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
The plastid is the site for a number of biosynthetic functions essential to plants, including photosynthesis, nitrogen assimilation and the synthesis of amino acids, as well as the synthesis of starch, fatty acids and numerous secondary products. These diverse biochemical functions rely on the controlled exchange of metabolites between the plastid and the surrounding cytosol. Plastids are double-membrane organelles. Although the outer envelope membrane contains a number of proteins that have been shown to have channel-like activities (Pohlmeyer et al. 1997), and therefore may act as a selectivity filter, the major site of the controlled exchange of metabolites across the plastid is the inner envelope (Fischer and Weber 2002). Numerous proteins located in the inner envelope membrane are needed for the transport of a variety of metabolites, including inorganic anions such as phosphate, sulfate, nitrite and chloride, organic anions such as amino acids and pyruvic acid, as well as adenylates and phosphorylated intermediates (Fischer and Weber 2002; Neuhaus and Wagner 2000).
To date, only a small subset of transporters of the plastid inner envelope has been biochemically characterized and the corresponding gene identified. These include the triose phosphate/phosphate exchanger, phosphoenolpyruvate/phosphate exchanger, 2-oxoglutarate/malate and glutamate/malate translocators, ADP/ATP translocator, glucose-6-phosphate/phosphate translocator, pentose phosphate/phosphate translocator and the glucose translocator (Flügge et al. 1989; Weber at al. 1995, 2000; Fischer et al. 1997; Neuhaus et al. 1997; Kammerer et al. 1998; Eicks et al. 2002; Taniguchi et al. 2002; Renné et al. 2003). It is thus clear that numerous additional transporters of the plastid inner envelope remain to be identified and characterized.
Several approaches can be used to identify and characterize transporters present in the plastid inner envelope. These range from the classical approach of biochemical purification of proteins followed by reconstitution into artificial membranes, to approaches using genomics, including the in silico analysis of the genome of Arabidopsis thaliana for the presence of genes encoding putative plastid membrane proteins as well as the analysis of the proteome of the plastid inner envelope using organic solvent extraction and MALDI–TOF analysis (Ferro et al. 2002, 2003; Koo and Ohlrogge 2002).
In this work, we demonstrate the identification of a novel protein present in the plastid inner envelope of A. thaliana and Spinacia oleracea that shows homology to animal transporters involved in inorganic phosphate (Pi), chloride and glutamate transport.
Materials and methods
Plant material
Wild-type Arabidopsis thaliana (L.) Heynh., accession Columbia (Arabidopsis Biological Resources Center, Ohio), was grown under continuous light on a mixture of 2/3 soil and 1/3 perlite at 19 °C and 60% relative humidity. Spinach (Spinacia oleracea L.) leaves were purchased at the local supermarket.
DNA constructs and expression in Escherichia coli
The full-length ANTR1 cDNA was isolated by screening the PRL-2 cDNA library (Newman et al. 1994) with a probe derived from a partial expressed sequence tag (EST). A similar screen for ANTR2 gave rise only to an incomplete cDNA. The full-length ANTR2 cDNA was isolated by a 5′-RACE–PCR approach (Frohman et al. 1988) on the Arabidopsis PRL-2 cDNA library. The PCR product corresponding to the missing 5′ end was inserted at the BglII site located near the 5′ end of the incomplete cDNA using standard molecular biology procedures.
The plasmid psmRS–GFP was used for the construction of the ANTR1–GFP fusion (Davis and Vierstra 1998). A PCR fragment including the whole coding region of ANTR1 was inserted in the XbaI–BamHI sites of psmRS–GFP in order to create an in-frame fusion protein with the ANTR1 protein sequence at the N-terminus and green fluorescent protein (GFP) at the C-terminus. The cassette containing the CaMV 35S promoter-ANTR1–GFP fusion-Nos terminator was excised by HindIII–EcoRI and inserted in the binary vector pBI121 (Clontech, Palo Alto, CA, USA). As a control the GFP from the plasmid psmRS–GFP was also subcloned in the binary vector pBI121.
Polypeptides representing the N-terminal hydrophilic portion of the mature ANTR1 and ANTR2 proteins were expressed in E. coli using the pET expression system (Novagen, Madison, WI, USA). Briefly, DNA fragments encoding portions of ANTR1 and ANTR2 were amplified by PCR using oligonucleotides creating NdeI and BamHI restriction sites at the 5′ and 3′ ends, respectively, and cloned in the same sites in the pET14b vector. The proteins were expressed in the E. coli strain BL21(DE3)LysE growing in media containing isopropyl β-d-thiogalactopyranoside (IPTG). Total protein extracts were analyzed by Western blotting using ANTR2 antibodies.
Plant transformation
Arabidopsis was transformed with the GFP and ANTR1–GFP constructs by the floral-dip method using Agrobacterium tumefaciens pGV3101 (Clough and Bent 1998). Transgenic plants were selected on Murashige and Skoog media containing 1% sucrose and 50 μg/ml kanamycin. Epidermal peels of the abaxial surface of leaves of transgenic plants were analyzed by epifluorescence microscopy.
Chloroplast isolation
Chloroplasts were isolated according to Douce and Joyard (1982) on a Percoll gradient. Leaves were cut from young plants (less than 4 weeks old) and stored in air at 4 °C overnight to reduce the starch content of the chloroplasts. For spinach leaves, the mid-rib of each leaf was removed before storing overnight at 4 °C. All further steps were performed at 4 °C. Leaves were homogenised with two short bursts (3–5 s each) in a Waring Blender in 3 vol. of Buffer H [0.33 M sucrose, 30 mM Na-pyrophosphate (pH 7.8), 0.1% BSA]. The homogenate was filtered through one layer of Miracloth and centrifuged for 10 min at 1,200 g. The pellet was gently resuspended in Buffer J [0.33 M sucrose, 10 mM MOPS–KOH (pH 7.8)] with a paintbrush. The suspension was filtered through one layer of Miracloth, layered on a 40/80% (v/v) Percoll step gradient in Buffer J and centrifuged at 3,000 g for 20 min. The intact chloroplasts were collected from the 40/80% interphase, diluted at least 5 times with Buffer J and centrifuged at 3,000 g for 5 min. Finally the chloroplasts were resuspended with a paintbrush in an appropriate volume of Buffer J. The amount of chlorophyll in the chloroplast preparation was determined by mixing 10 μl of purified chloroplasts with 10 ml of 80% (v/v) acetone. Aliquots of 1 ml were centrifuged for 30 s. The absorbance at 652 nm was determined and the amount of chlorophyll calculated with the formula: chlorophyll (mg/ml) = OD652×28.99.
Isolation of chloroplast envelopes
Envelopes were separated by methods adapted from Joyard et al. (1982). Intact chloroplasts were resuspended in a hypertonic medium [0.66 M sucrose, 10 mM MOPS–KOH (pH 7.8), 4 mM MgCl2], frozen at −20 °C for at least 1 h and thawed at room temperature for 1 h. The chloroplast envelopes were then stripped-off by homogenisation in a Dounce (Sigma, St. Louis, MO, USA) homogeniser (20–40 strokes). The freezing and homogenising were repeated once more. The homogenate was supplemented with sucrose to a final concentration of 1.1 M, overlaid with hypertonic medium (0.66 M sucrose) and Buffer J [0.33 M sucrose, 10 mM MOPS–KOH (pH 7.8)]. This gradient was centrifuged at 113,000 g at 4 °C for at least 3 h. Highly purified outer envelopes were isolated from the 0.33/0.66 M interphase, diluted at least 3 times with Buffer J and centrifuged at 208,000 g. Enriched inner envelopes were collected from the 0.66/1.1 M interphase, diluted at least 3 times with Buffer J and centrifuged for 60 min at 208,000 g. The envelope pellets were resuspended in an appropriate volume of Buffer J.
Total protein extraction and quantification
For small-scale extractions, 100 mg of plant material was ground with 100 μl of extraction buffer [80 mM Tris–HCl (pH 8.0), 5 mM DTT, 0.1 mM EDTA, 1 mM phenylmethylsulfonyl fluoride (PMSF), 1% Triton X-100] in an Eppendorf tube. For larger amounts the plant material was ground in liquid nitrogen, and the powder extracted with the same amount (w/v) of extraction buffer. Cell debris was separated by centrifugation at 10,000 g in a tabletop centrifuge. The supernatant was stored at −80 °C. The amount of protein in the different organelle fractions and from the total protein extractions was determined with the Bradford assay from BioRad (Richmond, CA, USA). In the Western analysis of crude extracts from various tissues, care was taken to load equal amounts of proteins on the gel, and the quality of the extracts and of the transfer was controlled by Ponceau Red staining.
Analysis of organelle and membrane fractions
The purity of organelles and membrane fractions was tested by Western blot analysis. Proteins were fractionated by SDS–PAGE. The sample denaturation was done in Laemmli buffer on ice for 1 h. The proteins were transferred onto nitrocellulose in a Trans-Blot electrophoretic cell (BioRad). Free binding sites were blocked with blocking buffer (8 g/l NaCl, 0.2 g/l KCl, 1.44 g/l Na2HPO4, 0.24 g/l KH2PO4, 0.1% Tween-20, 5% milk powder). The nitrocellulose membranes were incubated with the following organelle-specific antibodies: aquaporin RD28 for plasma membranes (Daniels et al. 1994), pyrophosphatase VPPase326 for tonoplast (Kim et al. 1994), porin for mitochondria (Heins et al. 1994), SEC12 for the ER (Bar-Peled and Raikhel 1997), TOC75 for outer-envelope proteins (Tranel et al. 1995), TIC110 for inner-envelope proteins (Lübeck et al. 1996) and LHCII for thylakoids. The antibody dilutions used were 1:500 for RD28, TIC110 and AtSEC12, 1:800 for porin, 1:1,000 for VPPase326 and TOC75, and 1:2,000 for LHCII. AtSEC12 antibody was purchased from Rose Biotechnology (Palo Alto, CA, USA). The ANTR2 rabbit antibody was made against the peptide NSIKINRSRAYYKSEESDIA that was coupled to keyhole-limpet protein (Eurogentec, Belgium). Antibodies were visualized with horseradish peroxidase conjugated to protein A and the Enhanced Chemiluminescence Kit (ECL; Amersham, UK) as recommended by the manufacturer.
Immunolocalization
Tissues were fixed in a solution containing 3.7% (w/v) formaldehyde, 5% (v/v) acetic acid, 50% (v/v) ethanol, for 4 h at room temperature, followed by 3 days at 4 °C. Tissues were then dehydrated in an ethanol gradient, cleared in xylol and embedded in paraffin wax. Section of 10 μm were dewaxed, hydrated and washed in water, and incubated in blocking buffer containing 10% (w/v) BioRad blotting-grade blocker in 30 mM Tris–HCl (pH 7.5), 37 mM NaCl and 0.5% (v/v) Tween-80. Sections were incubated with immune or pre-immune sera in blocking buffer for 2 h at room temperature followed by washes in TBS-T buffer containing 30 mM Tris–HCl (pH 7.5), 37 mM NaCl and 0.5% (v/v) Tween-80. The secondary antibody was a goat anti-rabbit immunoglobulin linked to alkaline phosphatase (Dako, Switzerland). The secondary antibody was diluted in blocking buffer and incubated with the sections for 1 h at room temperature. After washing (first washing with TBS-T buffer with Tween-80 followed by buffer without Tween), colour was developed using the Dako NBT/BCIP system. Sections were counterstained with 0.1% (w/v) alcian green for 20 s, dehydrated in an ethanol series, cleared in xylene and mounted with Eukitt (Kindler, Germany).
Results
Identification of two genes homologous to animal anion transporters
NaPi-1 was initially characterized as a gene expressed in rabbit kidney cells that confers sodium-dependant phosphate transport when expressed in Xenopus oocytes (Werner et al. 1991). Further studies have shown that in Xenopus oocytes, NaPi-1 acts as anion channel permeable to chloride and organic anions (Busch et al. 1996). More recently, the Caenorhabditis elegans EAT4 homologue was shown to be involved in glutamatergic neurotransmission (Lee et al. 1999), while the human homologue VGLUT1 has been shown to be a neuronal glutamate transporter that also exhibits a conductance for chloride (Bellocchio et al. 2000; Takamori et al. 2000). Another human homologue of NaPi-1, named AST, has been implicated in the transport of sialic acid (Verheijen et al. 1999).
We initially selected two expressed sequence tags from Arabidopsis that showed homology to the rabbit NaPi-1 gene. Two full-length cDNAs were subsequently isolated and named ANTR1 and ANTR2 for anion transporters. ANTR1 corresponds to the gene At2g29650, while ANTR2 corresponds to the gene At4g00370. ANTR1 and ANTR2 share 70% amino acid identity and 80% amino acid similarity (Fig. 1). The homology is high throughout the proteins except at the N-termini where the sequences are highly divergent.

Alignment of the Arabidopsis thaliana ANTR protein family. The consensus sequence for ACS family members is indicated by asterisks. The putative transit-peptide cleavage sites for the proteins ANTR1, ANTR2 and ANTR3 are indicated by the digits 1, 2 and 3, respectively, placed over the sequences. Alignment was made with the ClustalW1.74 program
Analysis of ANTR1 and ANTR2 with the BlastP program for homology with proteins having a characterized function reveals highest homology with the human VGLUT1 and AST, the C. elegans EAT4 and the rabbit NaPi-1. Alignment of ANTR2 protein with these animal proteins, as well as with a rice homologue, is shown in Fig. 2. The level of similarity between ANTR2 and the animal homologues ranges from 47% for NaPi-1 to 53% for VGLUT1 or AST, with the N-terminal hydrophilic domain being poorly conserved for all proteins. All these proteins belong to the Anion:Cation Symporter (ACS) family which itself is included in the major facilitator superfamily (MFS; Pao et al. 1998). Analysis of the hydropathy profiles of ANTR1 and ANTR2 revealed the presence of 12 putative transmembrane-spanning domains separated in two clusters of six domains by a larger hydrophilic stretch (Fig. 3), a feature common to members of the MFS superfamily. Both ANTR1 and ANTR2 also contain the seven residues that form a characteristic motif present in the members of the ACS family (Fig. 2; Pao et al. 1998).

Alignment of the Arabidopsis thaliana ANTR2 with a rice (Oryza sativa) homologue (OsANTR), and with the human (Homo sapiens) VGLUT1 and AST, the Caenorhabditis elegans EAT-4 and the rabbit (Oryctolagus cuniculus) NaPi-1 proteins. The consensus sequence for ACS family members is indicated by asterisks. Alignment was made with the ClustalW 1.74 program

Hydropathy profiles of ANTR1, ANTR2 and the human VGLUT1 homologue. Evaluation of the potential transmembrane-spanning domains was performed using the programs HMMTOP (http://www.enzim.hu/hmmtop), TMHMM (http://www.cbs.dtu.dk/services/TMHMM) and Tmpred (http://www.ch.embnet.org/software/TMPRED_form.html). The results were transposed on a Kyte–Doolittle plot using a window of 11 amino acids. Transmembrane segments predicted by at least two programs are indicated by black boxes while segments predicted by only one program are indicated by open boxes
Analysis of the Arabidopsis proteins with the ChloroP (Emanuelsson et al. 1999) and TargetP (Emanuelsson et al. 2000) programs revealed a putative chloroplast transit peptide for ANTR1 (ChloroP score 0.58, TargetP score 0.99), with a cleavage site between amino acid 59 (Arg) and 60 (Ser; Fig. 1). Use of the same programs for the analysis of ANTR2 gave ambiguous and non-conclusive results (ChloroP score 0.49, TargetP score 0.68).
ANTR2 is a chloroplast inner-envelope protein
The intracellular localization of the ANTR2 protein was examined by organelle and membrane fractionation. Analysis of the purity of the fractions was tested using antibodies recognizing proteins specific to various membranes. Antibodies against ANTR2 were raised against a unique 19-amino-acid peptide present in the N-terminal hydrophilic domain (see Materials and methods).
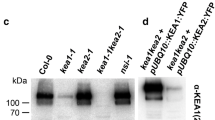
Preliminary analysis, by Western blotting, of fractions derived from Arabidopsis leaves and enriched in either plasma membrane, tonoplast, chloroplast or mitochondria, indicated that ANTR2 was mainly a chloroplast protein (data not shown). Subsequent analysis was thus focused on purification and fractionation of the chloroplast. Figure 4a shows that ANTR2 protein is enriched in the chloroplast fraction relative to a crude extract from whole shoots. ANTR2 antibodies recognize a single protein of 45 kDa. Analysis of the chloroplast fractions by Western blotting with antibodies raised against the endoplasmic reticulum protein Sec12, the tonoplast pyrophosphatase, the plasma membrane aquaporin RD28 and the mitochondrial porin, revealed that the chloroplast fraction was depleted of proteins derived from the membranes of other organelles (data not shown).

ANTR2 is localized in the chloroplast inner envelope. a Proteins (25 μg) from whole-leaf extracts (wl) and purified chloroplasts (pc) were separated on an SDS–PAGE gel and analyzed by Western blotting using polyclonal rabbit antibodies against ANTR2. The asterisk denotes 45 kDa. b Proteins were isolated from purified chloroplasts as well as inner and outer chloroplast envelopes and analyzed by Western blotting with antibodies against ANTR2, the outer envelope protein TOC75, the inner envelope protein TIC110 and the thylakoid protein LHCII. Lane 1, Arabidopsis purified chloroplasts (10 μg); lane 2, Arabidopsis outer envelopes (3 μg); lane 3, Arabidopsis inner envelopes (3 μg); lane 4, spinach (Spinacia oleracea) purified chloroplasts (30 μg); lane 5, spinach outer envelopes (16 μg); lane 6, spinach inner envelopes (16 μg)
Chloroplasts were then fractionated to separate the inner and outer envelopes. The extent of enrichment and contamination of the two fractions were analyzed by Western blotting with antibodies against the inner-envelope protein TIC110, the outer-envelope protein TOC75 and the thylakoid protein LHCII. Figure 4b (lanes 1–3) shows that ANTR2 is found enriched in the inner-envelope fraction and depleted in the outer-envelope fraction of Arabidopsis chloroplasts. Both envelope fractions had undetectable contamination from thylakoids, excluding the localization of the ANTR2 protein in these membranes. Similar results were obtained in experiments with envelopes isolated from spinach chloroplasts (Fig. 4b, lanes 4–6).
The localization of the ANTR2 protein to the plastid inner envelope indicated that the N-terminal hydrophilic domain may contain a cleavable transit peptide. Analysis of the protein with ChloroP and TargetP, combined with manual inspection of the N-terminal hydrophilic domain, revealed the presence of two potential cleavage sites, one between amino acids 28 (Ser) and 29 (Gly) and a second site between 87 (Arg) and 88 (Ala; Fig. 1). Since the second potential cleavage site is within the peptide that was used for production of antibodies, the ability of the antibodies to detect the hydrophilic peptide included between amino acids 88 and 139 of ANTR2 was checked. Figure 5 shows that the ANTR2 antibodies can detect the ANTR2 peptide expressed in E.coli while they failed to recognize a peptide representing the mature hydrophilic portion of ANTR1 comprising the region between amino acids 60 and 110.

Specificity of the ANTR2 antibodies. Proteins (5 μg) from whole Escherichia coli transformed with the pET14b vector (lanes 1) or with constructs leading to the expression of peptides representing the N-terminal hydrophilic portion of the mature ANTR1 (amino acids 60–110; lanes 2) or ANTR2 (amino acids 88–139; lanes 3) were separated on an SDS–PAGE gel. a Western blot analysis using polyclonal rabbit antibodies against ANTR2. b Coomassie blue staining of the SDS–PAGE gel showing the overexpressed ANTR1 and ANTR2 peptides. The asterisk denotes 6 kDa
Expression pattern of ANTR2
Western analysis of crude extracts from different tissues of Arabidopsis was performed with antibodies against ANTR2. Figure 6 shows that ANTR2 is predominantly expressed in leaves, stems and developing siliques, while expression in roots and flowers was below detection.

Expression pattern of ANTR2 in Arabidopsis. Western blotting was performed with equal loading (50 μg) of protein isolated from roots (lane 1), leaves (lane 2), inflorescence stems (lane 3), green siliques (lane 4), and flowers (lane 5). The arrow indicates 45 kDa
Expression of ANTR2 was further examined in leaves and developing seeds by immunolocalization. Cross-sections through leaves reveal expression of ANTR2 in both palisade and spongy mesophyll cells (Fig. 7a, b). No expression of ANTR2 protein is evident in cells of the leaf epidermis. In 7- to 9-day-old siliques containing embryos at the upturned-U stages, expression of ANTR2 is strongest in the embryo and weak but detectable in the cells of the outer integument, endosperm and cells of the silique walls (Fig. 7c, d).

Immunolocalization of ANTR2 in leaves and seeds of Arabidopsis. Paraffin sections of leaves (a, b) and 7- to 9-day-old siliques (c, d) were analyzed with antibodies against ANTR2 (b, d), as well as the corresponding pre-immune serum (a, c)
ANTR1 is a chloroplast protein
Rabbit antibodies raised against the entire N-terminal hydrophilic domain (110 amino acids) of ANTR1were found to significantly cross-react with the ANTR2 N-terminal hydrophilic domain of the mature peptide (amino acids 88–139), making its use inappropriate for localization of ANTR1 (data not shown). A hybrid protein was thus created whereby GFP was translationally fused to the C-terminal end of the full-length ANTR1. GFP and the ANTR1–GFP constructs were expressed under the control of the CaMV 35S promoter and stably transformed into Arabidopsis. Epidermal peels for the abaxial surface of leaves were examined by epifluorescence microscopy (Fig. 8). While GFP was found in the cytoplasm and nucleus but not in plastids of epidermal cells of transgenic plants expressing the unfused GFP, the green fluorescence was clearly localized in plastids in epidermal cells of transgenic plants expressing the ANTR1–GFP fusion. These data indicated that ANTR1 was localized to the plastids.

Localization of an ANTR1–GFP fusion in chloroplasts. Epidermal pavement cells from the abaxial surface of transgenic Arabiodopsis leaves expressing GFP (a, b) and the fusion protein ANTR1–GFP (c, d) were examined by epifluorescence microscopy. Autofluorescence from the plastids (a, c) was visualized with a Leitz I2 filter (excitation filter 450–490 nm and long-pass emission filter 515 nm) while GFP (b, d) was visualized with a Leitz L3 filter (excitation filter 450–490 nm, band-pass emission filter 520–525 nm)
The ANTR gene family in Arabidopsis
Analysis of the genome of Arabidopsis for homologues of ANTR1 and ANTR2 revealed the presence of four additional genes homologous to ANTR1 and ANTR2, namely At2g38060 (ANTR3), At3g46980 (ANTR4), At5g44370 (ANTR5) and At5g20380 (ANTR6). No complete cDNAs were available for ANTR3 and ANTR6. The protein sequences of ANTR3 and ANTR6 were thus deduced using a combination of approaches, including the annotation provided in TAIR (http://www.arabidopsis.org) or TIGR (http://www.tigr.org), alignment with ESTs from Arabidopsis and other plants, as well as alignments with the ANTR members for which the annotation has been confirmed with full-length cDNAs (ANTR1, ANTR2, ANTR4 and ANTR5). Analysis of the gene structure of ANTR6 and comparison with the exon/intron structure of ANTR1, ANTR2 and ANTR4 led to a predicted protein sequence slightly different from the TAIR annotation. The difference is mainly due to a selection of splice acceptor sites. Although a full-length cDNA was reported for ANTR6 (accession AY095993), its structure appeared aberrant since only a short open reading frame is generated. Analysis of this sequence indicates an insertion of 16 bases, which leads to a change in reading frame and a premature stop codon. The prediction of the ANTR6 protein is thus based on the exon/intron structure of the cDNA with the removal of the 16 bases.
The alignment of all ANTR proteins sequences is shown in Fig. 1.The level of homology between the various members of the family ranges from 29% amino acid identity and 49% similarity between ANTR4 and ANTR5 to 70% identity and 80% similarity between ANTR1 and ANTR2. The homology between ANTR members is lowest at the N-terminal hydrophilic region. The programs ChloroP and TargetP both predict a chloroplast targeting sequence for ANTR3 (ChloroP score 0.543, TargetP score 0.905; Fig. 1). The predictions with the same programs for ANTR4 and ANTR6 are more ambiguous. The structure of the ANTR5 protein appears unusual in that it lacks an extensive N-terminal hydrophilic domain common to all other ANTR proteins. This indicates that the protein may be located in a different sub-cellular compartment compared to other members of the family. The SignalP program (Nielsen et al. 1997) predicts an anchoring signal peptide at the N-terminal end of ANTR5. The gene structure of ANTR5 is also unusual in that it is composed of a single exon while all other ANTR genes are composed of at least eight exons. The various members of the ANTR family contain at least five of the seven residues that form the characteristic motif present in the members of the ACS family (Fig. 1).
Discussion
Chloroplast purification and membrane subfractionation have shown that ANTR2 is a protein located in the inner envelope of chloroplasts in both Arabidopsis and spinach. ANTR2 has not been previously identified as a plastid protein either through proteomic approaches combining membrane protein extraction with organic solvents and analysis of peptides with MALDI–TOF (Ferro et al. 2002, 2003), or in silico searches of the Arabidopsis genome using programs such as ChloroP or TargetP (Ferro et al. 2002; Koo and Ohlrogge 2002). This may be due, in part, to the failure of the ChloroP and TagetP programs to recognize the chloroplast targeting sequence.
Western analysis of crude protein extracts revealed that ANTR2 is mainly expressed in leaves, stems and developing siliques, while expression in roots and flowers is below detection. Immunolocalization revealed that ANTR2 is expressed in the leaf spongy and palisade mesophyll cells while expression in the leaf epidermal cells is undetectable. In developing seeds, ANTR2 expression is mainly located in the embryo with some weak expression in the integument, endosperm and cells of the silique wall. Although the developing embryo of Arabidopsis at the upturned-U stage is green and could be competent at photosynthesis, the integument and endosperm are non-photosynthetic. ANTR2 is thus expressed in both autotrophic and heterotrophic tissues. Although the expression pattern of the ANTR1 protein could not be determined, the localization of an ANTR1–GFP fusion protein to the chloroplast shows that ANTR1 is also a chloroplast protein.
Four additional homologues of ANTR1 and ANTR2 are found in the Arabidopsis genome. Although the localization of these additional proteins within the cell is unknown, the apparent lack of a chloroplast-targeting signal for ANTR5 suggests that not all ANTR proteins may be chloroplastic. All ANTR proteins are homologous to the animal NaPi-1 protein family as well as to other organic-anion transporters found in the ACS family of transporters, albeit at a lower level, including bacterial hexunorate and galactonate transporters. ANTR proteins share the main features of members of the ACS family, including the presence of 12 putative membrane-spanning domains clustered in 2 groups of 6, as well as the presence of a 7-amino-acid motif in the fourth putative transmembrane domain. To our knowledge, the ANTR genes from Arabidopsis are the first characterized plant genes that are members of the ACS family of permeases (Pao et al. 1998).
The ANTR1 and ANTR2 genes of Arabidopsis were isolated based on the high homology of the encoded proteins to the rabbit and human NaPi-1. Although NaPi-1 was initially identified as a protein expressed in rabbit kidney that induced a sodium-dependant Pi transport when expressed in Xenopus oocytes (Werner et al. 1991), the role of this protein in vivo still remains unclear. In contrast to NaPi-2, a protein unrelated to NaPi-1 but also expressed in rabbit kidney and mediating a strong sodium-dependant Pi transport in Xenopus ooyctes, the rabbit NaPi-1 shows only low affinity to Pi and the gene is poorly regulated by Pi supply (Werner et al. 1998). Expression of NaPi-1 in Xenopus has been shown to induce a chloride conductance that was inhibited by chloride-channel blockers, as well as a conductance for organic anions, such as benzylpenicillin (Busch et al. 1996). Detailed studies of the Pi transport induced by NaPi-1 expression in Xenopus oocytes indicated that NaPi-1 might be up-regulating a Pi transporter that is endogenous to Xenopus oocytes (Bröer et al. 1998). Mutation in a homologue of NaPi-1 in humans results in a deficiency in the transport of the organic anion sialic acid, reinforcing the concept of NaPi-1 as an organic-anion transporter (Verheijen et al. 1999). More recently, the human brain-specific NaPi-1 homologue (BNPI) has been shown to be responsible for the transport of glutamate into synaptic vesicles (Bellocchio et al. 2000; Takamori et al. 2000). In addition, this protein, which has been renamed VGLUT1, was shown to exhibit a conductance for chloride that is blocked by glutamate. VGLUT1 is expressed specifically in glutamatergic neurons (Bellocchio et al. 1998), and mutation in the NaPi-1 homologue EAT4 in C. elegans results in a defect in neurotransmission from these neurons (Lee et al. 1999). Together, these results indicate that NaPi-1 homologues in mammalian cells are primarily involved in the transport of glutamate (in neurons) and of other organic acids, as well as acting as a chloride channel. Although the evidence for a role of mammalian NaPi-1 homologues in Pi transport is unclear, it remains possible that these proteins may have Pi transport activity under some conditions. It is also possible that NaPi-1 may have multiple activities, similar to the animal EAAT5/ASCT1 protein, which exhibits both glutamate/amino acid transport activity and a chloride conductance (Zerangue and Kavanaugh 1996), and the chloroplast triose phosphate translocator, which also exhibits channel activity towards anions (Schwarz et al. 1994).
In view of the homology between the Arabidopsis ANTR proteins and the mammalian NaPi-1, VGLUT and AST proteins, three putative functions may be assigned to the plant chloroplast proteins, namely Pi transport, glutamate/organic-anion transport or chloride channel.
Several proteins have been identified in the plastid inner envelope that mediate the 1:1 counter-exchange of Pi with phosphorylated compounds. These include the triose phosphate/phosphate translocator (Flügge et al. 1989), the phosphoenolpyruvate/phosphate translocator (Fischer et al. 1997), the glucose-6-phosphate/phosphate translocator (Kammerer et al. 1998) and the pentose phosphate/phosphate translocator (Eicks et al. 2002). A proton-dependant phosphate transporter has also been recently identified in chloroplasts (Versaw and Harrison 2002). There is evidence for the presence of additional Pi transporters in the plastid envelope. For example, a transporter exchanging malate for Pi has been identified at the biochemical level in leukoplast envelopes from castor seed endosperm (Eastmond et al. 1997) while sulfate transport across the envelope of spinach chloroplast was linked to counter-transport of Pi (Mourioux and Douce 1979). In non-green plastids, synthesis of starch from glucose-6-phosphate and ATP leads to a 2:1 imbalance between phosphate import and phosphate export. A Pi transporter would be required for the exit of Pi out of the plastid since a high level of Pi would inhibit starch biosynthesis. Unidirectional transport of Pi across the envelope membrane of cauliflower-bud amyloplasts has been measured, although the corresponding protein has not been identified (Neuhaus and Maass 1996).
The inner envelope of the plastid contains transport systems for the entry and exit of numerous non-phosphorylated organic anions. These include the translocators for the exchange of glutamine and glutamate (Yu and Woo 1988) or glycerate and glycolate (Howitz and McCarty 1985), for which no proteins have been identified. Furthermore, unidentified transporters are also thought to be involved in the translocation of numerous amino acids synthesized in the plastids.
Chloride channels have been characterized in the envelopes of the chloroplast, either after reconstitution into artificial membranes (Heiber et al. 1995) or in native membranes (van den Wijngaard and Vredenberg 1997). One of these channels was implicated in chloroplast protein import (van den Wijngaard and Vredenberg 1997). Chloride and anion channels may be involved in the maintenance of charge balance and pH across the inner envelope. Besides the inner envelope triose phosphate/phosphate translocator, which has been shown to have anion channel activity in reconstituted membranes (Schwarz et al. 1994), the proteins involved in the various chloride channel activities of the plastid inner envelope have not been identified.
It is difficult at this point to define which of the multiple potential candidate solutes is most likely transported by the ANTR1 and ANTR2 proteins. Functional characterization of transport activity of a chloroplast protein through heterologous expression is made difficult by the inappropriate targeting of the proteins to endomembranes. Expression of chloroplast membrane transporters in yeast followed by protein purification and/or reconstitution into artificial liposomes has been a method of choice used to characterize a number of plastid transporters. Use of this reconstitution system, as well as analysis of transport activity in whole yeast expressing the ANTR 1 and ANTR2 proteins without the transit peptide, has however failed to detect any significant transport activity of ANTR1 and ANTR2 towards either Pi, chloride, glutamate or glutamine (data not shown). Reverse genetics and the inactivation of the ANTR genes may give some clues as to the role of the proteins in the plastids. The potential expression of several ANTR homologues in the plastids may complicate the analysis by reverse genetics and require the simultaneous inactivation or down-regulation of several members of the ANTR gene family. Although challenging, the study of the ANTR genes promises to bring novel information on the biochemistry of metabolites across the envelope of the plastids, and perhaps also of other membranes.
Abbreviations
- ACS:
-
Anion:Cation Symporter
- GFP:
-
green fluorescent protein
- Pi:
-
inorganic phosphate
References
Bar-Peled M, Raikhel NV (1997) Characterization of AtSEC12 and AtSAR1. proteins likely involved in endoplasmic reticulum and golgi transport. Plant Physiol 114:315–324
Bellocchio EE, Hu H, Pohorille A, Chan J, Pickel VM, Edwards RH (1998) The localization of the brain-specific inorganic phosphate transporter suggests a specific presynaptic role in glutamatergic transmission. J Neurosci 18:8648–8659
Bellocchio EE, Reimer RJ, Fremeau RT, Edwards RH (2000) Uptake of glutamate into synaptic vesicles by an inorganic phosphate transporter. Science 289:957–960
Bröer S, Schuster A, Wagner CA, Bröer A, Forster I, Biber J, Murer H, Werner A, Lang F, Busch AE (1998) Chloride conductance and Pi transport are separate functions induced by the expression of NaPi-1 in Xenopus oocytes. J Membr Biol 164:71–77
Busch AE, Schuster A, Waldegger S, Wagner CA, Zempel G, Broer S, Biber J, Murer H, Lang F (1996) Expression of a renal type I sodium/phosphate transporter (NaPi-1) induces a conductance in Xenopus oocytes permeable for organic and inorganic anions. Proc Natl Acad Sci USA 93:5347–5351
Clough SJ, Bent AF (1998) Floral dip: a simplified method for Agrobacterium-mediated transformation of Arabidopsis thaliana. Plant J 6:735–43
Daniels MJ, Mirkov TE, Chrispeels MJ (1994) The plasma membrane of Arabidopsis thaliana contains a mercury-insensitive aquaporin that is a homologue of the tonoplast water channel protein TIP. Plant Physiol 106:1325–1333
Davis SJ, Vierstra RD (1998) Soluble, highly fluorescent variants of green fluorescent protein (GFP) for use in higher plants. Plant Mol Biol 36:521–52
Douce R, Joyard J (1982) Purification of the chloroplast. In: Edelman M, Hallick RB, Chua NH (eds) Methods in chloroplast molecular biology. Elsevier, Amsterdam, pp 239–256
Eicks M, Maurino V, Knappe S, Flugge U-I, Fischer K (2002) The plastidic pentose phosphate translocator represents a link between the cytosolic and the plastidic pentose phosphate pathways in plants. Plant Physiol 128:512–522
Eastmond PJ, Dennis DT, Rawsthorne S (1997) Evidence that a malate/inorganic phosphate exchange translocator imports carbon across the leucoplast envelope for fatty acid synthesis in developing castor seed endosperm. Plant Physiol 114:851–856
Emanuelsson O, Nielsen H, von Heijne G (1999) ChloroP, a neural network-based method for predicting chloroplast transit peptides and their cleavage sites. Protein Sci 8:978–984
Emanuelsson O, Nielsen H, Brunak S, von Heijne G (2000) Predicting subcellular localization of proteins based on their N-terminal amino acid sequence. J Mol Biol 300:1005–1016
Ferro M, Salvi D, Rivière-Rolland H, Vermat T, Seigneuri-Berny D, Grunwald D, Garin J, Joyard J, Rolland N (2002) Integral membrane proteins of the chloroplast envelope: identification and subcellular localization of new transporters. Proc Natl Acad Sci USA 99:11487–11492
Ferro M, Salvi D, Brugière S, Miras S, Kowalski S, Louwagie M, Garin J, Joyard J, Rolland N (2003) Proteomics of the chloroplast envelope membranes from Arabidopsis thaliana. Mol Cell Proteomics 2:325–345
Fischer K, Weber A (2002) Transport of carbon in non-green plastids. Trends Plant Sci 7:345–351
Fischer K, Kammerer B, Gutensohn M, Arbinger B, Weber A, Häusler RE, Flügge UI (1997) A new class of plastidic phosphate translocators: a putative link between primary and secondary metabolism by the phosphoenolpyruvate/phosphate antiporter. Plant Cell 9:453–462
Flügge UI, Fischer K, Gross A, Sebald W, Lottspeich F, Eckerskorn C (1989) The triose phosphate-3-phosphoglycerate-phosphate translocator from spinach chloroplasts: nucleotide sequence of a full-length cDNA clone and import of the in vitro synthesized precursor protein into chloroplasts. EMBO J 8:39–46
Frohman MA, Dush MK, Martin GR (1988) Rapid production of full-length cDNAs from rare transcripts: amplification using a single gene-specific oligonucleotide primer. Proc Natl Acad Sci USA 85:8998–9002
Heiber T, Steinkamp T, Hinnah S, Schwarz M, Flügge UI, Weber A, Wagner R (1995) Ion channels in the chloroplast envelope membrane. Biochemistry 34:15906–15917
Heins L, Mentzel H, Schmid A, Benz R, Schmitz UK (1994) Biochemical, molecular, and functional characterization of porin isoforms from potato mitochondria. J Biol Chem 269:26402–26410
Howitz KT, McCarty RE (1985) Substrate specificity of the pea chloroplast glycolate transporter. Biochemistry 24:3645–3650
Joyard J, Grossman A, Bartlett SG, Douce R, Chua NH (1982) Characterization of envelope membrane polypeptides from spinach chloroplasts. J Biol Chem 257:1095–1101
Kammerer B, Fischer K, Hilpert B, Schubert S, Gutensohn M, Weber A, Flügge UI (1998) Molecular characterization of a carbon transporter in plastids from heterotrophic tissues: the glucose 6-phosphate/phosphate antiporter. Plant Cell 10:105–117
Kim EJ, Zhen RG, Rea PA (1994) Heterologous expression of plant vacuolar pyrophosphatase in yeast demonstrates sufficiency of the substrate-binding subunit for proton transport. Proc Natl Acad Sci USA 91:6128–6132
Koo AJK, Ohlrogge JB (2002) The predicted candidates of Arabidopsis plastid inner envelope membrane proteins and their expression profiles. Plant Physiol 130:823–836
Lee RY, Sawin ER, Chalfie M, Horvitz HR, Avery L (1999) EAT-4, a homolog of a mammalian sodium-dependent inorganic phosphate cotransporter, is necessary for glutamatergic neurotransmission in Caenorhabditis elegans. J Neurosci 19:159–167
Lübeck J, Soll J, Akita M, Nielsen E, Keegstra K (1996) Topology of IEP110, a component of the chloroplastic protein import machinery present in the inner envelope membrane. EMBO J 15:4230–4238
Mourioux G, Douce R (1979) Transport du sulfate à travers la double membrane limitante, ou enveloppe, des chloroplastes d’épinard. Biochimie 61:1283–1292
Neilsen H, Engelbrecht J, Brunak S, von Heijne G (1997) Identification of prokaryotic and eukaryotic signal peptides and prediction of their cleavage sites. Protein Eng 10:1–6
Neuhaus HE, Maass U (1996) Unidirectional transport of orthophosphate across the envelope of isolated cauliflower-bud amyloplasts. Planta 198:542–548
Neuhaus HE, Wagner R (2000) Solute pores, ion channels, and metabolite transporters in the outer and inner envelope membranes of higher plant plastids. Biochim Biophys Acta 1465:307–323
Neuhaus HE, Thom E, Möhlmann T, Steup M, Kampfenkel K (1997) Characterization of a novel ATP/ADP translocator located in the plastid envelope of Arabidopsis thaliana L. Plant J 11:73–82
Newman T, de Bruijn FJ, Green P, Keegstra K, Kende H, McIntosh L, Ohlrogge J, Raikhel N, Somerville SC, Thomashow M, Retzel E, Somerville CR (1994). Genes galore: a summary of methods for accessing results from large-scale partial sequencing of anonymous Arabidopsis cDNA clones. Plant Physiol 106:1241–1255
Pao SS, Paulsen IT, Saier MH (1998). Major facilitator superfamily. Microbiol Mol Biol Rev 62:1–34
Pohlmeyer K, Soll J, Steinkamp T, Hinnah S, Wagner R (1997) Isolation and characterization of an amino acid-selective channel protein present in the chloroplastic outer envelope membrane. Proc Natl Acad Sci USA 94:9504–9509
Renné P, Dressen U, Hebbeker U, Hille D, Flügge UI, Westhoff P, Weber APM (2003) The Arabidopsis mutant dct is deficient in the plastidic glutamate/malate translocator DiT2. Plant J 35:316–331
Schwarz M, Gross A, Steinkamp T, Flügge UI, Wagner R (1994) Ion channel properties of the reconstituted chloroplast triose phosphate/phosphate translocator. J Biol Chem 269:29481–29489
Takamori S, Rhee JS, Rosenmund C, Jahn R (2000) Identification of a vesicular glutamate transporter that defines a glutamatergic phenotype in neurons. Nature 407:189–194
Taniguchi M, Taniguchi Y, Kawasaki M, Takeda S, Kato T, Sato S, Tabata S, Miyake H, Sugiyama T (2002) Identifying and characterizing plastidic 2-oxoglutarate/malate and dicarboxylate transporters in Arabidopsis thaliana. Plant Cell Physiol 43:706–717
Tranel PJ, Froehlich J, Goyal A, Keegstra K (1995) A component of the chloroplastic protein import apparatus is targeted to the outer envelope membrane via a novel pathway. EMBO J 14:2436–2446
van den Wijngaard PWJ, Vredenberg WJ (1997) A 50-picosiemens anion channel of the chloroplast envelope is involved in chloroplast protein import. J Biol Chem 272:29430–29433
Verheijen FW, Verbeek E, Aula N, Beerens CEMT, Havelaar AC, Joose M, Peltonen L, Aula P, Galjaard H, van der Spek PJ, Mancini GMS (1999) A new gene, encoding an anion transporter, is mutated in sialic acid storage diseases. Nature Biotechnol 23:462–465
Versaw WK, Harrison MJ (2002) A chloroplast phosphate transporter, PHT2;1, influences allocation of phosphate within the plant and phosphate-starvation responses. Plant Cell 14:1751–1766
Weber A, Menzlaff E, Arbinger B, Gutensohn M, Eckerskorn C, Flügge UI (1995) The 2-oxoglutarate/malate translocator of chloroplast envelope membranes: molecular cloning of a transporter containing a 12-helix motif and expression of the functional protein in yeast cells. Biochemistry 34:2621–2627
Weber A, Servaites JC, Geiger DR, Kofler H, Hille D, Gröner F, Hebbeker U, Flügge UI (2000) Identification, purification and molecular cloning of a putative plastidic glucose translocator. Plant Cell 12:787–801
Werner A, Moore ML, Mantei N, Biber J, Semenza G, Murer H (1991) Cloning and expression of cDNA for a Na/Pi cotransport system of kidney cortex. Proc Natl Acad Sci USA 88:9608–9612
Werner A, Dehmelt L, Nalbant P (1998) Na+-dependent phosphate cotransporters: the NaPi protein families. J Exp Biol 201:3135–3142
Yu J, Woo KC (1988) Glutamine transport and the role of the glutamine translocator in chloroplasts. Plant Physiol 88:1048–1054
Zerangue N, Kavanaugh MP (1996) Flux coupling in a neuronal glutamate transporter. Nature 383:634–637
Acknowledgements
C. Roth and G. Menzel contributed equally to this work and are first co-authors. We thank M.J. Chrispeels (University of California-San Diego), P. Rea, (University of Pennsylvania), K. Keegstra (Michigan State University), U.K. Schmitz (Universität Hannover) and R. Leech (University of York) for providing antibodies against organelle proteins. We are also very grateful to A. Weber and I. Flügge (Universität zu Köln) for their help with the reconstitution of ANTR in artificial membranes. This work was funded, in part, with grants from the Fonds National Suisse de la Recherche Scientifique (Grant 3100-041903.94 and 31-61731.00), the Université de Lausanne and the Canton de Vaud.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Roth, C., Menzel, G., Petétot, J.MC. et al. Characterization of a protein of the plastid inner envelope having homology to animal inorganic phosphate, chloride and organic-anion transporters. Planta 218, 406–416 (2004). https://doi.org/10.1007/s00425-003-1121-5
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00425-003-1121-5