
Abstract
The effect of temperature on skeletal muscle ATP turnover, pulmonary oxygen uptake and single fibre ATP and PCr content was studied during intense cycling exercise. Six healthy male subjects performed 6-min intense (Δ50%LT-VO2peak) cycling, at 60 rpm, under conditions of normal (N) and elevated muscle temperature (ET). Muscle biopsies obtained from the vastus lateralis at rest, 2 and 6 min were analysed for homogenate ATP, PCr, lactate and glycogen, allowing estimation of anaerobic ATP turnover. Freeze-dried single fibres from biopsies were characterised according to their myosin heavy chain composition (type I, IIA or IIAX) and analysed for ATP and PCr content. Pulmonary gas exchange was measured throughout. There was no difference in pulmonary oxygen uptake between the trials. The elevation of muscle temperature resulted in a lower (P < 0.05) PCr content, higher (P < 0.05) lactate content and greater (P < 0.05) anaerobic ATP turnover after 2 min of exercise. There was no effect of temperature on these measures at 6 min. In single fibres it was observed that in ET, there was a lower (P < 0.05) PCr content in type I fibres after 2 min with no differences between conditions after 6 min. The present study demonstrates that elevation of muscle temperature results in a greater anaerobic ATP turnover and type I fibre PCr degradation during the initial 2 min of intense exercise.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
It has been well established that muscle temperature (T m) is an important determinant of the mechanical and metabolic performance in both amphibian and mammalian skeletal muscle (e.g. [7, 37]). This is significant because muscle temperature can vary considerably depending on the environmental conditions and the metabolic heat liberated from the muscle itself [1, 39]. Early work in this area by Edwards et al. [13] showed, in humans, that during isometric exercise, there was a greater anaerobic ATP turnover, when performed at elevated muscle temperatures. However, the majority of contractile activity, particularly during athletic events and daily activities, involves dynamic contractions, and during isometric contractions, energy turnover has been demonstrated in frog muscle to be different than during dynamic exercise [17].
In another investigation in humans, during 2-min cycling exercise, Febbraio et al. [16] found that there was greater glycogenolysis, glycolysis and ATP hydrolysis when exercise was performed after passive warming of the active musculature. Further human work has demonstrated that during 20 min of moderate intensity cycling, increasing skeletal muscle temperature results in an increased glycogenolysis, with no change in high energy phosphate degradation [44]. During exercise of this duration, however, the main energy source is oxidative phosphorylation [34, 36]. Investigations into the effects of muscle temperature on oxygen uptake have revealed somewhat ambiguous results. Indeed, it has been shown in two investigations, both in humans, that passively elevating muscle temperature has no appreciable effect on the oxygen uptake response during moderate and heavy cycling exercise [10, 31]. However, Ferguson et al. [18] found in humans that at 60 rpm, there was an elevation in oxygen uptake and a decrease in mechanical efficiency, whereas at 120 rpm, the converse was true. On a cautionary note, Ferguson and colleagues used blood lactate as a measure of anaerobic ATP turnover [12]. This may not reflect actual muscle ATP turnover since lactate can be taken up and metabolised by various tissues, including the heart and inactive muscles [20, 21].
It has been proposed that these differences may be due to a fibre-type-specific effect of alterations in skeletal muscle temperature [23, 41]. It is known that phosphocreatine (PCr) splits immediately upon contraction in amounts energetically sufficient to account for the amount of external work done [28]. Based on this, we have previously employed single fibre measures of PCr to show a preferential effect of elevations in skeletal muscle temperature on human muscle fibres with a predominance of myosin heavy chain (MHC IIA). This effect was noted during a 6-s maximal sprint where the average pedal rate was 120–140 rpm [24]. However, at lower pedal rates (60–100 rpm), it has been suggested that the effect of muscle temperature is directed at type I fibres [41]. There is no data investigating the effect of passive elevation of muscle temperature on the response of different fibre types during exercise, performed at relatively slow contraction frequencies.
Therefore, the aim of this study was to investigate the effects of passive elevation of T m on skeletal muscle metabolism and single fibre metabolism during intense cycling exercise performed at 60 rpm. It was hypothesised that elevating T m would lead to a greater anaerobic ATP turnover, concomitant with a greater PCr degradation, specifically in type I fibres after the elevation of T m.
Methods
Subjects
Six healthy male subjects (age, 25 ± 3 years; height, 1.85 ± 0.06 m; mass, 87 ± 16 kg; means±SD) with no history of muscle or metabolic disorders, volunteered for the study. All subjects were habitually physically active but not specifically trained. The subjects were fully informed of the purposes, risks and discomfort associated with the experiment before providing written, informed consent. This study conformed to current local guidelines and the Declaration of Helsinki and was approved by the University of Strathclyde Ethics Committee.
Pre-experimental procedures
Prior to the experimental, trials subjects’ \( \dot{V}{{\text{O}}_{{2{\text{peak}}}}} \) and lactate threshold (LT) were determined on an electromagnetically braked cycle ergometer (Excalibur Sport, Lode, BV, Groningen) at 60 rpm using a continuous incremental test to volitional fatigue, modified from the treadmill protocol described previously [29]. Power output was increased every 3 min by between 20 and 35 W. A fingertip blood sample was taken at the end of each 3 min stage and analysed immediately for lactate concentration (Lactate Pro, Akray KDK, Koyota, Japan). Heart rate and rate of perceived exertion were also recorded at the end of each stage with pulmonary gas exchange continuously measured breath-by-breath using an online gas analysis system (Oxycon Pro, Jaeger, Hoechberg, Germany). \( \dot{V}{{\text{O}}_{{2{\text{peak}}}}} \) was taken as the highest recording of \( \dot{V}{{\text{O}}_2} \) over a 30-s period. Lactate threshold was calculated by simultaneously solving two linear regression equations relating blood lactate to workload following identification of the visually observed breakpoint in blood lactate by two experienced reviewers [3] and is expressed as power output at LT (W). \( \dot{V}{{\text{O}}_2} \) at LT was calculated by solving the linear regression equation relating workload to \( \dot{V}{{\text{O}}_2} \) using power output at LT as the x variable. An external power output half way between LT and \( \dot{V}{{\text{O}}_{{2{\text{peak}}}}} \) (Δ50%) was then calculated and used in the subsequent experimental trials. At least 7 days later, a familiarisation trial was carried out to fully habituate the subjects to the experimental exercise trials.
Experimental protocol
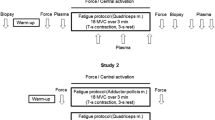
Subjects attended the laboratory on four separate occasions, each separated by a minimum of 7 days, to perform a 6- and a 2-min cycle exercise bout under normal (N) and elevated muscle temperature (ET) conditions. Prior to these trials, participants were instructed to refrain from strenuous physical exercise, caffeine and alcohol for the preceding 48-h period. During this period, they were also instructed to record their dietary intake and replicate this prior to subsequent trials. The order of conditions was counterbalanced, with the 6-min trial always preceding the 2-min trial in each condition.
On each occasion, subjects arrived at the laboratory following an overnight fast. On arrival, they inserted a rectal thermistor probe to allow continuous monitoring of rectal temperature. In N, subjects then rested for 30 min at normal room temperature (20–22°C) during which time the muscle biopsy site was prepared and a flexible muscle temperature probe inserted into the vastus lateralis. A resting biopsy was not taken in the 2-min trial to minimise the number of biopsies taken from each subject. Pilot work demonstrated that with the nutritional replication employed, a second resting biopsy was not necessary. In ET, after insertion of the rectal probe muscle, temperature was elevated through a combination of hot water and subsequently electrically heated blankets as described previously [23]. The same biopsy and temperature probe preparation to the legs was performed by opening the electric blanket for a minimal duration to avoid excessive heat loss. The heating protocol lasted approximately 1 h.
In the 6-min trials, under N or ET, a resting biopsy was taken at this point. Subjects then moved to the cycle ergometer where they sat at rest for 3 min (in ET the blankets remained wrapped around the legs). They then began cycling at the predetermined power output (Δ50%) at 60 rpm, for either 2 or 6 min. All subjects completed the full duration of every trial. Immediately after exercise, in all trials, a post-exercise biopsy was taken with the subject remaining seated on the bike. Pulmonary gas exchange was recorded continuously at rest and throughout the 6-min trials (the mouthpiece was removed ~15 s before the end of exercise to allow muscle biopsies to be taken).
Measurements
Muscle temperature
Muscle temperature (T m) was measured with a flexible muscle temperature probe (Ellab, Copenhagen, Denmark), which was inserted to a depth of 3 cm in the medial portion of the vastus lateralis muscle. The probe was inserted through a flexible venflon cannula (18 G, Becton Dickinson, Dublin, Ireland) and advanced ~0.5 cm beyond the end of the cannula into the muscle.
Muscle biopsies
Muscle samples were obtained from the medial part of the vastus lateralis using the needle biopsy technique [8]. Briefly, after injection of local anaesthetic (1% lidocaine), an incision (~1 cm) was made through the skin and fascia. When two biopsies were taken in the same trial, two separate incisions were made approximately 2–3 cm apart. The biopsy needle was introduced, sample collected and frozen immediately in liquid nitrogen and stored at −80°C for subsequent analyses. At all time points, including after the cessation of exercise, samples were frozen in <10 s, with no difference in time to freezing between treatments or timepoints. Contralateral legs were used for sampling in each of the N and ET trials, the order of which was counterbalanced.
Homogenate muscle analyses
The samples were freeze-dried and dissected free of blood and connective tissue, powdered and prepared for metabolite analysis. Approximately 25 mg of powdered tissue was extracted in 2 ml of 0.5 M perchloric acid and 1 mM EDTA, neutralised to pH 7.0 with 2.2 M KHCO3, and stored at −80°C. PCr and ATP content was determined in duplicate (CV of 1.3 and 1.4% for ATP and PCr, respectively) spectrophotometrically [26]. Lactate content was determined in duplicate (CV of 1.5%) flourometrically [33]. To determine muscle glycogen, approximately 4 mg tissue was extracted in 300 μl 1 M HCl and determined in duplicate (CV of 1.3%) flourometrically [33].
Single fibre analyses
In four subjects, selected because all biopsies for those subjects were deemed a suitable size to allow a sufficient number of single fibres to be dissected, approximately 40–50 fragments (3–4 mm) from each biopsy, were manually dissected under a low powered light microscope (Leica Microsytems, Wetzlar, Germany). Each fibre was then cut in half, with the first fragment used for MHC determination and the second used for single fibre ATP and PCr measurements.
MHC content was determined by sodium dodecyl sulphate (SDS) polyacrylamide gel electrophoresis using a method derived from Fauteck and Kandarian [14]. For approximately each millimetre of fibre fragment, 10 μl of SDS buffer was added. Electrophoresis (Bio-Rad Mini-Protean 3) was then carried out on 6% (crosslinking 2.7%) polyacrylamide resolving gels with 4% (crosslinking 2.7%) stacking gels, which were electrophoresed at 8°C for 18 h at a constant 100 V. Protein bands were visualised by silver staining, using a method modified from Oakley et al. [35], and quantified by densitometry (Bio-Rad GS8000 calibrated densitometer). Fibres were classified as either type I (100% MHC I), IIA (100% MHC IIA) or IIAX (all IIAX hybrid fibres).
The second fibre fragment was weighed on a quartz fibre fish-pole balance [33] calibrated by the spectrophotometrical determination of weighed p-nitrophenol crystals. Fibre fragments, weighing 1–5 μg, were then analysed luminometrically for ATP and PCr content using the luciferase method described by Wibom et al. [46]. Briefly, each fibre was extracted in 200 μl of trichloroacetic acid (2.5%) followed by neutralisation in 20 μl 2.2 M KHCO3. Fifty microlitres of the extract was then added to a sucrose buffer containing D-luciferin and the assay carried out on a luminometer (1251, Bio Orbit Oy, Turku, Finland). The coefficient of variation (CV) for duplicate determination of ATP and PCr content in the same fibre extracts (n = 26) was 7.2% and 4.5%, respectively.
Calculations
Anaerobic ATP turnover was calculated as: ΔPCr + 1.5Δ muscle lactate + 2 ΔATP [42]. The small quantity of ATP produced, or utilised, relating to the accumulation of glycolytic metabolites (e.g. pyruvate and glyceraldehyde-3-phosphate) are neglected as they represent <2% of ATP turnover in all cases [43]. The estimation of anaerobic ATP turnover is an underestimation since neither lactate efflux nor the amount metabolised by other tissues is measured [2]. Lactate release has, however, been shown to be the same when T m is passively elevated by about 3°C during knee extensor exercise [19] so this will not significantly affect comparisons between conditions.
Statistical analysis
Data were analysed by either two-way (temperature and time) analysis of variance (ANOVA) with repeated measures or paired t tests. Where a significant effect was detected, differences were located with post hoc t tests with Bonferroni correction. Significance was accepted at P < 0.05. Data are presented as means±SD.
Results
\( \dot{V}{{\text{O}}_{\text{2peak}}} \) and LT
\( \dot{V}{{\text{O}}_{\text{2peak}}} \) was 3.4 ± 0.5 l min−1 and was achieved at an external power output of 281 ± 54 W. LT occurred at 164 ± 49 W (58 ± 11% of \( \dot{V}{{\text{O}}_{\text{2peak}}} \)), with the external power output required to elicit Δ50% was calculated to be 219 ± 50 W. At exhaustion, the average heart rate was 181 ± 12 bpm (92 ± 5% of predicted max heart rate), and average respiratory exchange rate was 1.19 ± 0.04.
Dietary data
Energy intake for the 24-h period prior to experimental trials was 2432 ± 204 kcal, with 26 ± 3% fat, 59 ± 4% carbohydrate and 15 ± 1% fat.
Rectal and muscle temperature
The heating protocol resulted in a higher T m prior to the onset of exercise (34.7 ± 1.1°C in N vs. 37.4 ± 0.2°C in ET; P < 0.05). Rectal temperature was not different (P > 0.05) between trials (37.1 ± 0.2 and 37.3 ± 0.2°C in N and ET, respectively).
Homogenate muscle metabolites and ATP turnover
Elevation of T m had no effect (P > 0.05) on the resting content of any metabolites (Table 1). ATP content did not change during exercise under either condition.
After 2 min of exercise, PCr content was lower (P < 0.05) in ET compared to N, decreasing by 32.0 ± 8.3 mmol kg−1 (dm) in N and by 37.8 ± 10.6 mmol kg−1 (dm) in ET. PCr content continued to decrease (P < 0.05) and was the same after 6 min in both conditions. Lactate content was higher (P < 0.05) at 2 min in ET compared to N, increasing by 13.4 ± 3.9 mmol kg−1 (dm) and 8.6 ± 3.6 mmol kg−1 (dm), respectively. Lactate content continued to increase (P < 0.05) with no further difference at 6 min. Muscle glycogen content was decreased (P < 0.05) as a result of exercise. There were no differences in glycogen content between N and ET at any time point. After 6 min of exercise, there were no differences in the concentrations of metabolites between temperature conditions.
From these changes in metabolites, it was calculated that during the first 2 min of exercise, there was a 28% greater (P < 0.05) total anaerobic ATP turnover during ET (Fig. 1). This increase was the result of a greater (P < 0.05) glycolytic ATP turnover and a greater (P = 0.05) ATP turnover from PCr hydrolysis. Between 2 and 6 min as well as the overall period of exercise (0–6 min), elevation of T m had no further effect on anaerobic ATP turnover, although there was a tendency (P = 0.06) for a greater rate of glycolytic ATP turnover between 0 and 6 min in ET, compared to N (Fig. 1).

Glycolytic, PCr, and total anaerobic ATP turnover between 0 and 2, 2–6 and 0–6 min during heavy exercise under conditions of normal (N) and elevated (ET) muscle temperature. Values are means±SD (N = 6). Significant difference († P < 0.05) between conditions. Values are expressed as mmol kg−1 (dm) s−1
Single fibre ATP and PCr content
Elevation of T m had no effect (P > 0.05) on resting single fibre ATP content (Table 2). There was also no difference in resting ATP content between fibre groups. After 2 min of exercise, in both N and ET, ATP content was lower (P < 0.01) compared to rest in all fibre types. Thereafter, ATP content was only reduced in type IIA fibres in both N and ET. When muscle temperature was elevated, ATP content was 3.0 ± 0.6 mmol kg−1 (dm) lower (P < 0.01), compared to N, in type I fibres after 6 min of exercise.
Elevation of T m had no effect (P > 0.05) on resting single fibre PCr content, with this being higher in type IIA and IIAX compared to type I fibres (Table 3). After 2 min of exercise, PCr content was lower (P < 0.01) in all fibre types in both N and ET. In N PCr content reduced (P < 0.01) further from 2 to 6 min in type I and IIA fibres, with PCr content only reduced in IIA fibres at this time point.
The main finding of the single fibre analysis was that after 2 min of exercise, there was a 6.7 ± 1.5 mmol kg−1 (dm) lower (P < 0.01) PCr content in type I fibres in ET compared to N. This reflects a 25% greater PCr degradation in type I fibres during ET, compared to N. There were no differences between conditions after 6 min of exercise.
Pulmonary \( \dot{V}{{\text{O}}_{{2}}} \)
When pulmonary oxygen uptake was compared between N and ET, no difference was observed at any time point (Fig. 2).

Pulmonary \( \dot{V}{{\text{O}}_2} \) during 6 min of heavy exercise under conditions of normal (N, filled circles) and elevated (ET, closed circles) muscle temperature
Discussion
The present investigation has demonstrated that passively elevating the temperature of exercising muscle before the performance of sustained heavy exercise results in an increased anaerobic ATP turnover, with no differences in muscle glycogen, during the initial 2 min of exercise. The increase in anaerobic ATP turnover was associated with a greater PCr utilisation in fibres with a predominance of MHC I.
Previous work investigating the effects of elevating muscle temperature on anaerobic ATP turnover has demonstrated a greater anaerobic metabolism during intense isometric exercise [13], maximal sprint exercise [23], and intense cycling exercise [16]. The present investigation observed similar results, although muscle glycogen levels in the current study were unaffected by temperature, during the initial 2 min of exercise, however, when exercise continued, there were no differences in anaerobic ATP turnover between temperature conditions. This is a similar finding to that of Ferguson et al. [19] who did not observe an effect of passively increasing muscle temperature on energy turnover during 10 min of knee extensor exercise. It is worth pointing out that in that study, muscle biopsies were only obtained at the end of exercise, potentially missing any temperature induced changes present in the initial few minutes of exercise.
The reason for the greater anaerobic ATP turnover observed at higher T m is likely to be due to a greater cross-bridge cycling between actin and myosin. Indeed, Karatzaferi et al. [30] have clearly demonstrated the temperature dependency of cross-bridge cycling in relation to the force produced during the power stroke of the cycle. This itself is likely to be the result of an effect of the increase in temperature on mATPase activity [27, 45] as well as several other enzymes involved in ATP turnover. For example, creatine kinase is known to have an optimum temperature of around 42°C [47], and its activity is, therefore, likely to be enhanced after heating. The finding of a similar glycogen breakdown when comparing N and ET was a surprising result. An increase in lactate accumulation with heating has been demonstrated in previous studies [13, 16], although interestingly other work has found an increase in glycogen breakdown without a concomitant increase in muscle lactate [44]. The authors of the latter study suggested that this could be due to an increase in muscle lactate efflux in heated trials, although this was not directly measured. This assertion would not explain our findings, and work from our group has found that lactate efflux from muscle is not altered with heating during exercise [19].
As the exercise duration proceeds beyond the initial period, contribution from aerobic sources becomes more significant, as can be seen from the reduced anaerobic ATP turnover from 2 to 6 min of exercise. Previous work has demonstrated that during sustained cycling at a similar intensity and pedal cadence (60 rpm), passively elevating T m resulted in a greater pulmonary \( \dot{V} \)O2 [18]. The present study, however, found that elevated T m does not increase pulmonary \( \dot{V}{{\text{O}}_{{2}}} \), an observation also made by Burnley et al. [9] and Koga et al. [31], albeit at slightly faster pedal rates, as well as during single leg dynamic knee-extensor exercise [19]. The reason for these contradictory results is unclear, as the mode and intensity of exercise were similar between these investigations. It had previously been suggested that elevations in T m could increase \( \dot{V}{{\text{O}}_{{2}}} \) via a Q10 effect on mitochondrial respiration and/or through the back-leak of protons through the inner mitochondrial membrane. The current study would support the observation that mitochondrial function and physical performance only appear to be compromised at muscle temperature exceeding 40°C [15].
One possible explanation for the lack of T m effect on pulmonary \( \dot{V}{{\text{O}}_{{2}}} \), as well as anaerobic ATP turnover as exercise progresses may be related to the exercise induced rise in T m [39]. It is possible that a temperature effect is observed at the onset of exercise but not at the end as T m in N will approach the levels reached in ET, due to metabolic heat production. Whilst, due to technical difficulties, no measure of T m was made post-exercise in the present study, previous work has shown that when T m is raised by ~3°C prior to exercise, it is still approximately 1.5°C higher at the end of heavy knee-extensor [19] and cycling exercise [31]. It may therefore be the case that this smaller T m difference may not be of sufficient magnitude to alter energy turnover in the latter part of exercise, as was the case in the first 2 min.
Whilst the majority of previous research has focused on investigating the effect of alterations in muscle temperature in homogenate muscle samples, it is important to take into account the different muscle fibres that have diverse contractile and metabolic properties. It has been estimated that at the workload of the current study, the power required will be <50% of maximal power [40], which could likely be generated exclusively from type I fibres. However, it is clear from the single fibre data of the present study that type II fibres are recruited during this exercise. It is therefore prudent to discuss the effect of temperature on different fibre types. Measurements of single fibre metabolism have now been used in a variety of studies and are providing more detailed information on the metabolic response to exercise [5, 6, 25]. These methods are based on the knowledge that the PCr content of single muscle fibres gives a good indication of the activity of muscle fibres [11, 28].
Based on the early findings of Ranatunga et al. [38], in animal muscle, it was hypothesised that type I fibres were more sensitive to perturbations in temperature. This assertion was supported, in humans, by the findings of Sargeant and Rademaker [41] and Ferguson et al. [18]. However, both these studies relied on correlations between data to show a cause and effect, making the reliability of the thesis unclear. Furthermore, we have recently shown that during short-term (6 s) maximal sprint exercise (cadence of ~160 rpm) fibres with a predominance of MHC IIA have a greater PCr and ATP utilisation when muscle temperature is passively elevated [24]. From this, it appears likely that there may be a velocity dependent effect of muscle temperature on different fibre types. Such a view is supported by the current findings. During cycling at 60 rpm, the current study has shown that single fibre PCr utilisation was greater in type I fibres in the initial 2 min during ET. This coincides with the greater anaerobic ATP turnover observed in the analysis of muscle homogenates in the initial part of the exercise (Fig. 1). One caveat to this analysis was that due to lack of muscle tissue collected, we were only able to carry out the single fibre analysis in four subjects, and so further work in a greater number of subjects is necessary to support these assertions. Furthermore, it is worth pointing out that the results of the single fibres do not entirely match with the homogenate data. However, making comparisons between these two sets of data is problematic, as the numbers of each fibre type used in the analysis does not correlate with the average MHC composition of our subjects (e.g. 38.2 ± 12.5% MHC I, 58.7 ± 11.4% MHC IIA and 3.1 ± 2.7% MHC IIX).
The greater PCr utilisation in type I fibres may be explained further by considering the individual efficiency–velocity relationships of different fibre types. In the present study, the pedal rate during cycling was maintained at 60 rpm. This cadence is thought to be close to the optimum velocity for maximum efficiency of type I fibres [40]. An increase in temperature would induce a shift to the right of the efficiency–velocity relationship for the recruited fibre population, reflecting the change in the force–velocity and power–velocity relationships of muscle that occurs when temperature is elevated [38]. For type I fibres, such a shift would mean the fibres would be working on the ascending curve of the efficiency–velocity relationship. This would therefore lead to a decrease in efficiency and an increased energy turnover for a given mechanical power output. On the other hand, at this cadence, no such affect is seen in type II fibres. These fibres are likely, at 60 rpm, to be operating at the lower part of the ascending limb of the efficiency–velocity curve where a rightward shift will have minimal effect.
From a practical view point, it could be considered that as anaerobic energy turnover, and type I fibre metabolism, is elevated in the initial part of exercise in ET, warming the muscles before an athletic event may not be advised. There may be particularly deleterious effects for endurance athletes who have a high percentage of type I fibres [22]. In support of this, a previous work has found that temperature induced changes in mechanical efficiency correlates with the proportion of type I fibres [4]. In the current study, if we estimate mechanical efficiency (Table 4), as we have previously [23], we do not observe any correlations between the magnitude of change in anaerobic ATP turnover or mechanical efficiency. The reasons for this are not clear but may be due to the participants in the study of Bell and Ferguson [4] being female, although further work would be needed to confirm this. Another point of consideration for athletes would also be the contraction frequency at which exercise is performed as the response to changes in muscle temperature has been shown to be velocity dependent, with efficiency being higher at greater cadences [18]. This may also explain why endurance athletes tend to adopt such high cadences, which may have a beneficial effect on mechanical efficiency. The current study is not without limitations. It is well established now that a continuum of different fibre types with a variety of contractile properties exist [32]. Whilst we have separated out fibres into three discreet groups, including hybrid fibres, there is still a relatively wide range of MHC type content within each group. Ideally, one would be able to acquire enough fibres to further subdivide each group into pure and hybrid fibre subtypes, allowing a more detailed analysis. This was not possible in the current study as the amount of tissue was limited and the dissection of the single fibres is a laborious and skilled task. A further limitation to the current investigation is the relatively low subject numbers, due to the laborious nature of the methods and the requirements for large amounts of quality muscle tissue. Whilst the current single fibre analysis has provided valuable information on the metabolic response in single fibres to exercise at elevated muscle temperature, a greater number of subjects would have reduced the chances of a type II error.
In conclusion, the present study has demonstrated that passively increasing the temperature of the working muscle leads to an increase in anaerobic ATP turnover during the first 2 min of heavy exercise. This occurs alongside a greater PCr degradation in muscle fibres expressing a predominance of MHC type I. There was, however, no effect of elevating T m on anaerobic ATP turnover as exercise progressed or on pulmonary \( \dot{V}{{\text{O}}_{{2}}} \) at any point throughout the exercise period.
References
Asmussen E, Boje O (1945) Body temperature and the capacity for work. Acta Physiol Scand 10:1–22
Bangsbo J, Gollink PD, Graham TE, Juel C, Kiens B, Mizuno M, Saltin B (1990) Anaerobic energy production and O2 deficit–debt relationship during exhaustive exercise in humans. J Physiol 422:539–559
Beaver WL, Wasserman K, Whipp BJ (1985) Improved detection of lactate threshold during exercise using a log–log transformation. J Appl Physiol 59:1936–1940
Bell MP, Ferguson RA (2009) Interaction between muscle temperature and contraction velocity affects mechanical efficiency during moderate-intensity cycling exercise in young and older women. Journal of Applied Physiology 107:763–769
Beltman JG, de Haan A, Haan H, Gerrits HL, van Mechelen W, Sargeant AJ (2004) Metabolically assessed muscle fibre recruitment in brief isometric contractions at different intensities. Eur J Appl Physiol 92:485–492
Beltman JG, Sargeant AJ, Haan H, van Mechelen W, de Haan A (2004) Changes in PCr/Cr ratio on single characterized muscle fibre fragments after only a few maximal voluntary contractions in humans. Acta Physiol Scand 180:187–193
Bennett AF (1984) Thermal dependence of muscle function. Am J Physiol 247:R217–R229
Bergstrom J (1962) Muscle electrolytes in man. Scand J Clin Lab Invest suppl 68:1–101
Burnley M, Doust JH, Ball D, Jones AM (2002) Effects of prior heavy exercise on Vo2 kinetics during heavy exercise are related to changes in muscle activity. J Appl Physiol 93:167–174
Burnley M, Doust JH, Jones AM (2002) Effects of prior heavy exercise, prior sprint exercise and passive warming on oxygen uptake kinetics during heavy exercise in humans. Eur J Appl Physiol 87:424–432
Conjard A, Pette D (1999) Phosphocreatine as a marker of contractile activity in single muscle fibres. Pflugers Archiv 438:278–282
di Prampero PE, Ferretti G (1999) The energetics of anaerobic muscle metabolism: a reappraisal of older and recent concepts. Respiration Physiology 118:103–115
Edwards RHT, Harris RC, Hultman E, Kaijser L, Koh D, Nordesjo L-O (1972) Effect of temperature on muscle energy metabolism and endurance during successive isometric contractions, sustained to fatigue, of the quadriceps muscle in man. J Physiol 220:335–352
Fauteck SP, Kandarian SC (1995) Sensitive detection of myosin heavy chain composition in skeletal muscle under different loading conditions. Am J Physiol 268:C419–C424
Febbraio MA (2000) Does muscle function and metabolism affect exercise performance in the heat? Exerc Sport Sci Rev 28:171–176
Febbraio MA, Carey MF, Snow RJ, Stathis CG, Hargreaves M (1996) Influence of elevated muscle temperature on metabolism during intense, dynamic exercise. Am J Physiol 271:1251–1255
Fenn WO (1923) A quantitative comparison between the energy liberated and the work performed by the isolated sartorius muscle of the frog. J Physiol 58:175–203
Ferguson RA, Ball D, Sargeant AJ (2002) Effect of muscle temperature on rate of oxygen uptake during exercise in humans at different contraction frequencies. J Exp Biol 205:981–987
Ferguson RA, Krustrup P, Kjaer M, Mohr M, Ball D, Bangsbo J (2006) Effect of temperature on skeletal muscle energy turnover during dynamic knee-extensor exercise in humans. J Appl Physiol 101:47–52
Gladden LB (2000) Muscle as a consumer of lactate. Med Sci Sports Exerc 32:764–771
Gladden LB (2004) Lactate metabolism: a new paradigm for the third millennium. J Physiol 558:5–30
Gollnick PD, Armstrong RB, Saubert CW, Piehl K, Saltin B (1972) Enzyme activity and fiber composition in skeletal muscle of untrained and trained men. Journal of Applied Physiology 33:312–319
Gray SR, De Vito G, Nimmo MA, Farina D, Ferguson RA (2006) Skeletal muscle ATP turnover and muscle fiber conduction velocity are elevated at higher muscle temperatures during maximal power output development in humans. Am J Physiol 290:R376–R382
Gray SR, Soderlund K, Ferguson RA (2008) ATP and phosphocreatine utilization in single human muscle fibres during the development of maximal power output at elevated muscle temperatures. J Sports Sci 26:701–707
Greenhaff PL, Nevill AM, Söderlund K, Bodin K, Boobis LH, Williams C, Hultman E (1994) The metabolic responses of human type I and II muscle fibres during maximal treadmill sprinting. J Physiol 478:149–155
Harris RC, Hultman E, Nordesjö L-O (1974) Glycogen, glycolytic intermediates and high-energy phosphates determined in biopsy samples of musculus quadriceps femoris of man at rest. Methods and variance of values. Scand J Clin Lab Invest 33:109–120
He Z-H, Bottinelli R, Pellegrino MA, Ferenczi MA, Reggiani C (2000) ATP consumption and efficiency of human single muscle fibres with different myosin isoform composition. Biophysical Journal 79:945–961
Infante AA, Klaupiks D, Davies RE (1965) Phosphorylcreatine consumption during single-working contractions of isolated muscle. Biochim Biophys Acta 94:504–515
Jones. AM (1998) A five year physiological case study of an olympic runner. Br J Sp Med 32:39–43
Karatzaferi C, Chinn MK, Cooke R (2004) The force exerted by a muscle cross-bridge depends directly on the strength of the actomyosin bond. Biophysical Journal 87:2532–2544
Koga S, Shiojiri T, Kondo N, Barstow TJ (1997) Effect of increased muscle temperature on oxygen uptake kinetics during exercise. J Appl Physiol 83:1333–1338
Larsson L, Moss RL (1993) Maximum velocity of shortening in relation to myosin isoform composition in single fibres from human skeletal muscles. J Physiol 472:595–614
Lowry OH, Passonneau JV (1972) A flexible system of enzymatic analysis. Academic, New York
Medbo JI, Tabata I (1993) Anaerobic energy release in working muscle during 30s to 3 min of exhausting bicycling. J Appl Physiol 75:1654–1660
Oakley BR, Kirsch DR, Morris NR (1980) A simplified ultrasensitive silver stain for detecting proteins in polyacrylamide gels. Analytical Biochemistry 105(1):361–363
Parolin ML, Chesley A, Matsos MP, Spriet LL, Jones NL, Heigenhauser GJF (1999) Regulation of skeletal muscle glycogen phoshorylase and PDH during maximal intermittent exercise. Am J Physiol 277:E890–E900
Rall JA, Woledge RC (1990) Influence of temperature on mechanics and energetics of muscle contraction. Am J Physiol 259:197–203
Ranatunga KW (1984) The force–velocity relation of rat fast- and slow-twitch muscles examined at different temperatures. J Physiol 351:517–529
Saltin B, Gagge AP, Stolwijk JAJ (1968) Muscle temperature during submaximal exercise in man. J Appl Physiol 25:679–688
Sargeant AJ, Jones DA (1995) The significance of motor unit variability in sustaining mechanical output of muscle. In: Gandevia SC, Enoka RM, McComas AJ, Stuart DG, Thomas CK (eds) Fatigue: neural and muscular mechanisms. Plenum, New York, pp 323–338
Sargeant AJ, Rademaker A (1996) Human muscle power in the locomotory range of contraction velocities increases with temperature due to an increase in power generated by type I fibres. J Physiol 491:128P
Spriet LL (1995) Anaerobic metabolism during high-intensity exercise. In: Hargreaves M (ed) Exercise metabolism. Human Kinetics, Illinois, pp 1–40
Spriet LL, Söderlund K, Bergstrom M, Hultman E (1987) Anaerobic energy release in skeletal muscle during electrical stimulation in men. J Appl Physiol 62:611–615
Starkie RL, Hargreaves M, Lambert DL, Proietto J, Febbraio MA (1999) Effect of temperature on muscle metabolism during submaximal exercise in humans. Exp Physiol 84:775–784
Steinen GJM, Kiers JL, Bottinelli R, Reggiani C (1996) Myofibrillar ATPase activity in skinned human skeletal muscle fibres: fibre types and temperature dependence. J Physiol 493:299–307
Wibom R, Söderlund K, Lundin A, Hultman E (1991) A luminometric method for the determination of ATP and phosphocreatine in single human skeletal muscle fibres. J Biolumin and Chemilumin 6:123–129
Wyss M, Schlegel J, James P, Eppenberger HM, Wallimann T (1990) Mitochondrial creatine kinase from chicken brain. Purification, biophysical characterization, and generation of heterodimeric and heterooctameric molecules with subunits of other creatine kinase isoenzymes. J Biol Chem 265:15900–15908
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Gray, S.R., Soderlund, K., Watson, M. et al. Skeletal muscle ATP turnover and single fibre ATP and PCr content during intense exercise at different muscle temperatures in humans. Pflugers Arch - Eur J Physiol 462, 885–893 (2011). https://doi.org/10.1007/s00424-011-1032-4
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00424-011-1032-4