
Abstract
We addressed the question of whether hypertrophy induced by compensatory overload differs according to innervation status, and how fibroblast growth factors (FGF) and hepatocyte growth factor (HGF) mRNAs are expressed in the rat plantaris muscle during overload (OL) and/or denervation. Male Wistar rats were divided into four groups (Normal-Cont, Normal-OL, Denervated-Cont, and Denervated-OL). according to the plantaris denervation and/or overload. Three weeks later, plantaris weight in Denervated-Cont and Denervated-OL was significantly lower than in the Normal-Cont. The muscle weights in the Normal-OL were higher than in the Normal-Cont. The muscle weights in the Denervated-OL were higher than in the Denervated-Cont. Three days after the treatment, FGF-2, FGF-6, FGF-7 and HGF mRNAs in the Normal-OL were significantly higher than those in the Normal-Cont. FGF-2, FGF-6, FGF-7 and HGF mRNAs in the Denervated-OL were also significantly higher after 3 days than those in the Denervated-Cont. After 7 days, FGF-2, FGF-5, FGF-6, FGF-7 and HGF mRNAs were significantly higher in the Normal-OL than those in the Normal-Cont. At 21 days, FGF-1, FGF-6 and HGF mRNA levels were significantly increased. In the Denervated-OL, FGF-2, FGF-7 and HGF mRNAs at 7 days, and FGF-2 mRNA at 21 days were significantly higher than those in the Denervated-Cont. FGF-2 and FGF-6 mRNA levels decreased significantly following denervation; however, FGF-1, FGF-5, FGF-7 and HGF mRNA levels increased and maintained this increase for the 21-days treatment period. Muscle hypertrophy was thus induced by compensatory overload irrespective of innervation status, possibly in association with certain FGFs and HGF. The differential mRNA expression patterns of FGFs and HGF observed following compensatory overload and/or denervation suggest distinct roles for individual FGFs and HGF in muscle hypertrophy and/or atrophy.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Mammalian skeletal muscles undergo hypertrophy following stretch or compensatory overload induced by removal of synergists [40, 46]. Satellite cells, the normally inactive myogenic nuclei that participate in developmental and regenerative growth of skeletal muscle [35], are known to play an important role in hypertrophy. In response to stretch or compensatory overload, satellite cells are activated to enter the mitotic cycle and proliferate, and activated satellite cells fuse into preexisting fibres or form new fibres [35]. Inactivation of satellite cells by gamma irradiation prevents muscle hypertrophy following compensatory overload [26, 29].
The most prominent feature of denervated skeletal muscle is rapid atrophy of muscle fibres [10, 24, 25]. Short-term denervation induces satellite cells to enter the mitotic cycle [20, 21, 23], whereas long-term denervation decreases the number of both myonuclei and satellite cells [6, 28, 33, 42]. Satellite cells isolated from denervated muscle for 1 week or longer are resistant to recruitment into the mitotic cycle [17]. Thus, it is well established that the behaviour of satellite cells is influenced by muscle denervation, although little is known about the factors influencing such behaviour.
Fibroblast growth factors (FGFs) stimulate proliferation and inhibit differentiation in myogenic cell lines [1, 15, 27], and FGF-1, FGF-2, FGF-5, FGF-6, FGF-7 and hepatocyte growth factor (HGF) are expressed in rat satellite cells isolated from myofibres [16]. FGFs are considered to be powerful regulators of skeletal muscle in vitro and may be associated with muscle hypertrophy and atrophy in vivo. However, the expression patterns of FGFs and HGF during overload-induced hypertrophy in mammalian skeletal muscle are not well understood, and even less is known about these patterns in denervated skeletal muscle.
In cultured myofibres, continuous rhythmic stretching results in increased protein synthesis and prevents the atrophy that is noted in non-stimulated muscle cells [41]. Low-frequency electrical stimulation transforms fast muscle fibres into slow muscle fibres in denervated muscle, whilst high-frequency stimulation produces a change in the reverse direction [12]. These results suggest that changes in fibre sizes and types in skeletal muscle are important to muscle activity itself. However, there is no evidence that passive stretching independent of neural input in vivo causes muscle hypertrophy. We have developed a model of passive stretching of the plantaris muscle independent of neural input by combining ablation of the synergists with surgical denervation of the tibial nerve, which leaves intact the neural input and contractile activity of the antagonists. Using this model, we determined whether hypertrophy induced by compensatory overload differs according to innervation status and the mRNA expression patterns of FGFs and HGF during overload and/or denervation.
Materials and methods
Animal care and surgical procedures
Male Wistar rats aged 10 weeks and weighing 200–230 g were used. All animals were housed in cages at a constant room temperature (22±2 °C) and fed ad libitum. All surgical procedures were performed under anaesthesia. All experiments and procedures conformed to the Guideline for the Care and Use of Laboratory Animals of the Health Sciences University of Hokkaido. Experiments were initiated by the unilateral removal of the gastrocnemius and soleus muscles [45], and sham operations were carried out on the contralateral leg, which served as the control (Cont). Also, the animals were allocated randomly into innervated and denervated groups. Rat calf muscles (plantaris, soleus and gastrocnemius) are innervated by the tibial nerve whilst the antagonists (tibialis anterior and extensor digitorum longus muscles) are innervated by the common fibular nerve. Denervation was performed by cutting the tibial nerve bilaterally. For the denervated control leg (Denervated-Cont), the common fibular nerve was also cut. For the denervated overloaded leg (Denervated-OL), the common fibular nerve was left intact. Consequently, the plantaris muscle in the Denervated-OL underwent passive stretch via the contractile activities of its antagonists. In the Denervated-Cont, the passive stretch of the plantaris muscle was prevented by simultaneous denervation of its antagonists. To prevent reinnervation, 5-mm segments of the nerves were removed and the proximal stumps ligated. The overloaded and control legs in the innervated group served as Normal-OL and Normal-Cont, respectively. This study thus comprised four groups: Normal-Cont, Normal-OL, Denervated-Cont, and Denervated-OL, according to denervation and/or overload status. The animals were sacrificed 3, 7 or 21 days after treatment. The plantaris muscle was dissected quickly, freed of any fat and connective tissue, weighed, frozen in liquid nitrogen and stored at −80 °C.
Analysis of myosin heavy chain (MHC) isoforms
Six muscles of each group 21 days after compensatory overload and/or denervation were used for the analysis of MHC isoforms by SDS-PAGE according to [30]. Small muscle pieces were homogenized in 40 vol 5 M urea, 2 M thiourea, 10 mM sodium pyrophosphate and 0.1% 2-mercaptoethanol using a glass homogenizer. The homogenates were diluted 75-fold in a solution composed of 62.5 mM TRIS-HCl (pH 6.8), 2% SDS, 10% glycerol, 5% 2-mercaptoethanol and 0.02% bromophenol blue and loaded onto the gel. The stacking gel comprised 3.5% acrylamide, 0.07% bis-acrylamide, 70 mM TRIS-HCl (pH 6.8), 0.4% SDS, 0.1% N,N,N′,N′-tetramethylethylenediamine (TEMED) and 0.1% ammonium persulphate. The separating gel was 7.5% acrylamide, 0.2% bis-acrylamide, 30% glycerol, 200 mM TRIS/100 mM glycine (pH 8.6), 0.4% SDS, 0.1% TEMED and 0.06% ammonium persulphate. SDS-PAGE was carried out at a constant voltage of 200 V (4 °C) for 36 h. After SDS-PAGE, gels were silver-stained using a Wako Silver Stain Kit (Wako, Tokyo, Japan). The percentage distribution of MHC isoforms was estimated using NIH Image 1.54.
Analysis of gene expression by RT-PCR
Six muscles of each group 3, 7 or 21 days after compensatory overload and/or denervation were used for the analysis of the mRNA expression of FGFs and HGF. Total RNA was extracted by the guanidinium isothiocyanate-phenol-chloroform method [5]. RNA concentrations were determined spectrophotometrically at 260 nm and the integrity of the RNAs was determined by gel electrophoresis. Total RNA (1 µg) was used to synthesize first-strand cDNA in reverse transcriptase (RT) buffer containing 1×PCR buffer (Applied Biosystems, Tokyo, Japan), 5 mM MgCl2, 1 mM deoxynucleic acid triphosphates (dNTPs), 50 ng/µl oligo(dT) primer (Novagen, Darmstadt, Germany), 1 U/µl ribonuclease inhibitor (Takara, Tokyo, Japan), and 0.25 U/µl avian myeloblastosis virus (AMV)-reverse transcriptase XL (Takara) in a total volume of 20 µl. For amplification, a PCR master mixture containing 1×PCR buffer (Applied Biosystems), 1.5 mM MgCl2, 0.2 mM dNTPs, 0.025 U/µl Taq polymerase (Ampli Taq Gold, Applied Biosystems) and 0.4 µM of each sense and antisense primer specific for FGF-1, FGF-2, FGF-5, FGF-6, FGF-7, FGF-8 and hepatocyte growth factor (HGF) was prepared just before use. From this mixture, a 29-µl aliquot was removed and added to 1 µl cDNA and then placed in a temperature-controlled system (PCR Thermal Cycler SP, Takara). The PCR cycling program consisted of denaturation at 94 °C for 30 s, annealing at various temperatures (53.5–58 °C) for 30 s and elongation at 72 °C for 90 s. The PCR products were electrophoresed through 2% agarose gels, stained with ethidium bromide, photographed and quantified using NIH Image 1.54.
Specific primers used are summarized in Table 1. Each PCR primer was designed using OLIGO software (Medprobe) or published sequences [16] and the oligonucleotides were purchased from Hokkaido System Science (Sapporo, Japan). The RT-PCR products were sequenced by a double-strand sequencing service (Hokkaido System Science) to confirm the corresponding rat sequences. In a preliminary experiment, each cDNA was amplified for various cycle numbers (cycles 33–37 for FGF-1, cycles 29–33 for FGF-2, cycles 29–36 for FGF-5, cycles 34–38 for FGF-6, cycles 26–30 for FGF-7, cycles 30-36 for FGF-8 and cycles 32–36 for HGF) to establish a suitable PCR amplification (cDNAn) and amplification efficiency (R) of the unsaturated logarithmic phase. R remained constant during the unsaturated logarithmic phase and did not differ between groups. The estimation was repeated using all the primers, and identical results were obtained. The cDNA value at cycle 0 (cDNA0) was calculated from the equation log cDNAn=log cDNA0+n·log(1+R), where n is the cycle of amplification [11, 14]. As the cDNA0 is proportional to the quantity of starting mRNA in the sample [11, 14], this allowed quantitative comparisons of the PCR product between groups. The cDNA0 value for each PCR product is shown relative to the value in Normal-Cont group (=1.0). In our preliminary experiments, FGF-4 was expressed only in rat embryonic calf muscle and not in adult muscle, whilst FGF-3 was not detectable in embryonic or adult calf muscles (A. Yamaguchi, unpublished observations). Consequently, FGF-3 and FGF-4 were not analysed in this study.
Statistics
Standard procedures were employed to calculate the means and SD. One-way ANOVA was used for the comparison between groups. Differences between the means were regarded as significant at P<0.05.
Results
Figure 1 shows the weights of the plantaris muscle in each group 21 days after compensatory overload and/or denervation. The muscle weight in the Normal-OL group was significantly higher than in the Normal-Cont group. The muscle weights in the Denervated-Cont and Denervated-OL groups were significantly lower than in the Normal-Cont group. The muscle weight in the Denervated-OL group was significantly higher than in the Denervated-Cont group.

The weights of rat plantaris muscles 21 days after compensatory overload (OL) and/or denervation. *P<0.05 vs. non-denervated, non-overloaded muscle (Normal-Cont); † P<0.05 vs. the denervated non-overloaded muscle (Denervated-Cont)
MHC isoforms in the plantaris muscle 21 days after compensatory overload and/or denervation are shown in Table 2. The Normal-OL group possessed a higher percentage of the type-I MHC isoform than the Normal-Cont group. The Denervated-OL group also possessed a higher percentage of type-I MHC than the Denervated-Cont group. The Denervated-Cont and Denervated-OL groups possessed higher percentages of types-I and -IIa MHC isoforms and a smaller percentage of the type-IIb MHC isoform than the Normal-Cont group.
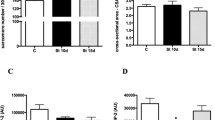
Figure 2 shows FGF-1 mRNA expression in the plantaris muscle 3, 7 and 21 days after compensatory overload and/or denervation. FGF-1 mRNA expression 3 days after treatment did not differ among groups. FGF-1 mRNA levels in the Normal-OL group 21 days after compensatory overload were significantly higher than those in the Normal-Cont group. After 7 and 21 days, FGF-1 mRNAs in the Denervated-Cont and Denervated-OL groups were significantly higher than those in the Normal-Cont group.

Fibroblast growth factor-1 (FGF-1) mRNA expression in rat plantaris muscles 3, 7 and 21 days after compensatory overload and/or denervation. A Ethidium-stained products; B FGF-1 mRNA expression relative to that in the Normal-Cont group (=1.0). *P<0.05 vs. Normal-Cont
FGF-2 mRNA levels in the Normal-OL group 3 and 7 days after compensatory overload were significantly higher than those in the Normal-Cont group. FGF-2 mRNA levels in the Denervated-OL group after 3, 7 and 21 days were significantly higher than those in the Denervated-Cont group. FGF-2 mRNA levels in the Denervated-Cont group after 3 and 21 days were significantly lower than those in the Normal-Cont group (Fig. 3).

FGF-2 mRNA expression in the plantaris muscle 3, 7 and 21 days after compensatory overload and/or denervation. A Ethidium-stained products; B FGF-2 mRNA expression relative to that in the Normal-Cont group (=1.0). *P<0.05 vs. Normal-Cont. † P<0.05 vs. Denervated-Cont
FGF-5 mRNA levels in the Normal-Cont group 3 and 21 days after compensatory overload did not differ from those in the Normal-OL group, however, FGF-5 mRNA level in the Normal-Cont group after 7 days was significantly higher than those in the Normal-Cont group. FGF-5 mRNA levels in the Denervated-Cont group after 3, 7 and 21 days did not differ from those in the Denervated-OL group. FGF-5 mRNA levels in the Denervated-Cont and Denervated-OL groups after 3, 7 and 21 days were significantly higher than those in the Normal-Cont group (Fig. 4).

FGF-5 mRNA expression in the plantaris muscle 3, 7 and 21 days after compensatory overload and/or denervation. A Ethidium-stained products; B FGF-5 mRNA expression relative to that in the Normal-Cont group (=1.0). *P<0.05 vs. Normal-Cont
FGF-6 mRNA levels in the Normal-OL group 3, 7 and 21 days after compensatory overload were significantly higher than those in the Normal-Cont group. Also, the FGF-6 mRNA level in the Denervated-OL group after 3 days was significantly higher than that in the Denervated-Cont group. FGF-6 mRNA levels in the Denervated-Cont group after 3, 7 and 21 days were significantly lower than those in the Normal-Cont group. FGF-6 mRNA levels in the Denervated-OL group 7 and 21 days after the treatments were significantly lower than those in the Normal-Cont group (Fig. 5).

FGF-6 mRNA expression in the plantaris muscle 3, 7 and 21 days after compensatory overload and/or denervation. A Ethidium-stained products; B FGF-6 mRNA expression relative to that in the Normal-Cont group (=1.0). *P<0.05 vs. Normal-Cont. † P<0.05 vs. Denervated-Cont
FGF-7 mRNA levels in the Normal-OL group 3, 7 and 21 days after compensatory overload were significantly higher than those in the Normal-Cont group. Also, FGF-7 mRNA levels in the Denervated-OL group after 3 and 7 days were significantly higher than those in the Denervated-Cont group. FGF-7 mRNA levels in the Denervated-Cont and Denervated-OL groups after 3, 7 and 21 days were significantly higher than those in the Normal-Cont group (Fig. 6).

FGF-7 mRNA expression in the plantaris muscle 3, 7 and 21 days after compensatory overload and/or denervation. A Ethidium-stained products; B FGF-7 mRNA expression relative to that in the Normal-Cont group (=1.0). *P<0.05 vs. Normal-Cont. † P<0.05 vs. Denervated-Cont
FGF-8 mRNA expression in the plantaris muscle 3, 7 and 21 days after compensatory overload and/or denervation did not differ among the groups.
HGF mRNA levels in the Normal-OL group 3, 7 and 21 days after compensatory overload were significantly higher than those in the Normal-Cont group. Also, HGF mRNA levels in the Denervated-OL group after 3 and 7 days were significantly higher than those in the Denervated-Cont group. HGF mRNA levels in the Denervated-Cont group after 3 and 7 days were significantly higher than those in the Normal-Cont group. HGF mRNA levels in the Denervated-OL group after 3, 7 and 21 days were significantly higher than those in the Normal-Cont group (Fig. 7).

Hepatocyte growth factor (HGF) mRNA expression in the plantaris muscle 3, 7 and 21 days after compensatory overload and/or denervation. A Ethidium-stained products; B HGF mRNA expression relative to that in the Normal-Cont group (=1.0). *P<0.05 vs. Normal-Cont. † P<0.05 vs. Denervated-Cont
Discussion
In this study, the expression of FGF-2, FGF-6, FGF-7 and HGF mRNAs increased shortly after initiating the compensatory overload, but not that of FGF-1, FGF-5 and FGF-8 mRNAs. Denervation reduced the FGF-2 and FGF-6 mRNA levels, increased the FGF-1, FGF-5, FGF-7 and HGF mRNA levels but did not change the FGF-8 mRNA level, in comparison with the innervated plantaris. Adding FGFs and HGF to cultured rat myofibres, FGF-1, FGF-2, FGF-4, FGF-6 and HGF enhance satellite cell proliferation, whereas FGF-5 and FGF-7 do not [16]. Moreover, several, but not all, FGFs are expressed in response to stretch-induced hypertrophy in avian muscle [22]. Thus, even if they belong to the FGF family, the gene expression differs in response to different stimuli. It is likely that individual FGFs do not always play the same role in the morphologic growth or maintenance of skeletal muscle.
In this study, the expression of FGF-2, FGF-6, FGF-7, and HGF mRNAs increased with overload-induced hypertrophy. Although the roles of FGFs and HGF in mammalian skeletal muscle in vivo remain obscure, these observations raise the possibility that overload-induced muscle hypertrophy may be influenced by certain FGFs. FGFs stimulate proliferation and inhibit differentiation in myogenic cell lines [1, 15, 27]. HGF also plays a role in enhancing proliferation in satellite cells and in inhibiting cell differentiation [9]. FGF-2 and FGF-4 are also chemotactic for limb myogenic cells [18, 43] and HGF induces increased chemotaxis of satellite cells [4]. Addition of anti-HGF antibodies to cultured satellite cells eliminates their activation in response to mechanical stretching [39]. In adult muscle, satellite cells are normally quiescent, but become mitotically active in response to various stimuli, including stretching and compensatory overload [35, 36, 44]. The activated satellite cells fuse into preexisting fibres or form new muscle fibres [35]. The inactivation of satellite cells by gamma irradiation prevents muscle hypertrophy following compensatory overload [26, 29]. Because FGF-2 and FGF-6 proteins are expressed in adult rat skeletal muscle [16, 31, 32] and the presence of HGF protein in satellite cells isolated from adult rat muscle has been detected by immunofluorescence [39], the enhanced expression of FGF-2, FGF-6, FGF-7, and HGF during overload in this study may be associated with muscle hypertrophy, possibly by activation of satellite cells.
The FGF-5 mRNA level did not change 3 days after compensatory overload, but was increased at 7 or more days. FGF-2, FGF-6 and HGF mRNA levels were increased at 3 days and maintained this increase during compensatory overload. It is difficult to explain what roles the prolonged expressions of FGFs and HGF mRNAs might have. FGFs and HGF may play an important role in the continuing muscle fibre hypertrophy as well as in the activation of satellite cells. FGFs and HGF may also play a role in immature fibre formation, which is often observed in overloaded muscles [46]. Further studies are needed to elucidate these phenomena
Little is known about the expression pattern of FGFs and HGF in denervated skeletal muscle. The observations in this study indicate that mRNAs of individual FGFs and HGF were expressed very differently patterns following denervation; FGF-2 and FGF-6 mRNA levels were decreased, FGF-1, FGF-5, FGF-7 and HGF mRNA levels increased and FGF-8 mRNA level unchanged following denervation. The functional meaning of these changes following denervation are unknown. FGF-2 and FGF-6 are the most prominent factors in satellite cell activation [1, 8, 16, 27]. As reduced expression of FGF-2 and FGF-6 mRNAs was shown shortly after denervation in this study, these FGFs are most closely associated with denervation atrophy. Little is known about the factors involved in muscle atrophy following denervation. Proliferating cell nuclear antigen-positive and myogenin-positive cells, the proliferative and differentiative compartments of cells, in an isolated cell culture from denervated muscle are decreased after 1 week or more, and denervation for 1 week or longer hinders recruitment into the mitotic cycle [17]. Prolonged denervation decreases the number of myonuclei and satellite cells [6, 28, 33, 42]. These observations suggest that muscle atrophy following denervation may be related to inactivation of myonuclei and satellite cells. Collectively, the reduced expression of FGF-2 and FGF-6 may induce the inactivation of myonuclei and satellite cells and may be related to muscle atrophy following denervation.
In previous studies, satellite cell proliferation has been observed during short-term denervation in vivo and mitotic activation is found in the first few days or weeks after denervation using the [3H]-thymidine incorporation strategy [20, 21, 23]. In this study, FGF-5, FGF-7, and HGF mRNAs increased shortly after denervation. Thus, some FGFs may be associated with the activation of satellite cells after short-term denervation.
The increased expression of FGF-1, FGF-5, and FGF-7 mRNAs was maintained during denervation in this study. Myotubes begin to appear a few weeks after denervation and are present for several months [13, 28, 34]. The myotubes observed during denervation fail to grow and to mature into normal muscle fibres. As two important effects of FGFs observed in in vitro studies are the suppression of cell differentiation and the acceleration of cell proliferation, the enhanced expression of FGFs may also play a role in preventing new fibres from maturing. Ectopic application of FGFs can prevent apoptotic cell death [7] and support cell survival in various organisms [38]. Long-term denervation causes remarkable atrophy of muscle fibres and muscle fibre degeneration, but even though degeneration progressed for 25 months, surviving muscle fibres were found [6]. Thus, FGFs may be related to the survival of fibres in muscle with long-term denervation. Denervation causes a progressive increase in connective tissue [13, 19]. Since FGFs are expressed in connective tissue as well as in muscle fibres or satellite cells [16], they may play roles not only in the events observed in the denervated muscle fibres, but also in the connective tissue.
An interesting finding of this investigation was that a change in muscle fibre size due to compensatory overload occurred in both denervated and innervated muscle. As the overload in this study resulted from ablation of the plantaris synergists, the hypertrophy of the remaining plantaris muscle resulted from the increased active shortening from various daily activities in compensation for the ablated synergists. However, as the denervated muscle cannot contract spontaneously, the ablation of the denervated synergists could not increase the compensatory active shortening of the plantaris muscle. The only possible effect counteracting atrophy in the denervated plantaris muscle following compensatory overload is increased passive stretching associated with activities of the innervated antagonists. Stretching of the anterior latissimus dorsi muscle in birds reportedly also produces muscle enlargement [2, 3]. In mammalian skeletal muscles, short-term stretching due to plaster cast immobilization in the lengthened position causes an increase of muscle mass [46]. In cultured myofibres, continuous rhythmic stretching increases protein synthesis and prevents the atrophy noted in unstimulated muscle cells [41]. These observations would support the notion that the counteracting effect of the overload on the denervated atrophy is caused by increased passive stretch, independent of neural input. However, the fact that compensatory overload in this study only partly prevented the atrophy in a denervated plantaris muscle should be addressed.
Denervation in this study increased the percentage of types-I and -IIa MHC isoforms at the expense of type-IIb. As we did not estimate the sizes of each fibre type, it is unknown whether the changes in MHC isoforms were dependent on fibre type conversion or selective atrophy of fibre types. Muscle fibre composition following denervation changes slow fibres into fast fibres in slow muscle [10, 12, 37], but not in fast muscle, including the plantaris muscle [10]. Also, type-IIB fibres atrophy more severely than type-I fibres [10, 19, 24, 25]. Taken together, it appears that the decreased percentage of the type IIb MHC isoform resulted from the greater atrophy of type IIB fibres following denervation. As the changes in MHC isoforms in this study were very small, it is difficult to find a relationship between MHC isoforms and FGFs.
Finally, our results suggest that the ablation of synergists in the plantaris muscle during denervation, the model of passive stretching independent of neural input, gave rise to muscle hypertrophy, and that the hypertrophy may be associated with certain FGFs and HGF. The differential mRNA expression patterns of FGFs and HGF observed following compensatory overload and/or denervation suggest distinct roles for individual FGFs and HGF in muscle hypertrophy and atrophy.
References
Allen RE, Rankin LL (1990) Regulation of satellite cells during skeletal muscle growth and development. Proc Soc Exp Biol Med 194:81–86
Alway SE, Gonyea WJ, Davis ME (1990) Muscle fiber formation and fiber hypertrophy during the onset of stretch-overload. Am J Physiol 259:C92–C102
Antonio J, Gonyea WJ (1993) Role of muscle fiber hypertrophy and hyperplasia in intermittently stretched avian muscle. J Appl Physiol 74:1893–1898
Bischoff R (1997) Chemotaxis of skeletal muscle satellite cells. Dev Dyn 208:505–515
Chomczynski P, Sacchi N (1987) Single step method of RNA isolation by guanidium thiocyanate-phenol-chloroform extraction. Anal Biochem 162:156–159
Dedkov EI, Kostrominova TY, Borisov AB, Carlson BM (2001) Reparative myogenesis in long-term denervated skeletal muscles of adult rats results in a reduction of the satellite cell population. Anat Rec 263:139–154
Fallon JF, López A, Ros MA, Savage MP, Olwin BB, Simandl BK (1994) FGF-2: apical ectodermal ridge growth signal for chick limb development. Science 264:104–106
Floss T, Arnold H-H, Braun T (1997) A role for FGF-6 in skeletal muscle regeneration. Gene Dev 11:2040–2051
Gal-Levi R, Leshem Y, Aoki S, Nakamura T, Halevy O (1998) Hepatocyte growth factor plays a dual role in regulating skeletal muscle satellite cell proliferation and differentiation. Biochim Biophys Acta 1402:39–51
Girlanda P, Dattola R, Vita G, Oteri G, Presti FL, Messina C (1982) Effect of electrotherapy on denervated muscles in rabbits: an electrophysiological and morphological study. Exp Neurol 77:483–491
Golde TE, Estus S, Usiak M, Younkin LH, Younkin SG (1990) Expression of ß amyloid protein precursor mRNAs: recognition of a novel alternatively spliced form and quantitation in Alzheimer’s disease using PCR. Neuron 4:253–267
Gorza L, Gundersen K, Lomo T, Schiaffino S, Westgaard RH (1988) Slow-to-fast transformation of denervated soleus muscles by chronic high-frequency stimulation in the rat. J Physiol (Lond) 402:627–649
Gulati AK (1988) Long-term retention of regenerative capability after denervation of skeletal muscle, and dependency of late differentiation on innervation. Anat Rec 220:429–434
Hannon K, Smith II CK, Bales KR, Santerre RF (1992) Temporal and quantitative analysis of myogenic regulatory and growth factor gene expression in the developing mouse embryo. Dev Biol 151:137–144
Hannon K, Kudla AJ, McAvoy MJ, Clase KL, Olwin BB (1996) Differentially expressed fibroblast growth factors regulate skeletal muscle development through autocrine and paracrine mechanisms. J Cell Biol 132:1151–1159
Kästner S, Rivera AJ, Yablonka-Reuveni Z (2000) Gene expression patterns of the fibroblast growth factors and their receptors during myogenesis of rat satellite cells. J Histochem Cytochem 48:1079–1096
Kuschel R, Yablonka-Reuveni Z, Bornemann A (1999) Satellite cells on isolated myofibers from normal and denervated adult rat muscle. J Histochem Cytochem 47:1375–1383
Li SG, Anderson R, Reginelli AD, Muneoka K (1996) FGF-2 influences cell movements and gene expression during limb development. J Exp Zool 274:234–247
Lu D-X, Huang S-K, Carlson BM (1997) Electron microscopic study of long-term denervated rat skeletal muscle. Anat Rec 248:355–365
McGeachie JK (1989) Sustained cell proliferation in denervated skeletal muscle of mice. Cell Tiss Res 257:455–457
McGeachie J, Allbrook D (1978) Cell proliferation in skeletal muscle following denervation or tenotomy. Cell Tiss Res 193:259–267
Mitchell P, Steenstrup T, Hannon K (1999) Expression of fibroblast growth factor family during postnatal skeletal muscle hypertrophy. J Appl Physiol 86:313–319
Murray MA, Robbins N (1982) Cell proliferation in denervated muscle: time course, distribution and relation to disuse. Neuroscience 7:1817–1822
Niederle B, Mayr R (1978) Course of denervation atrophy in type I and type II fibres of rat extensor digitorum longus muscle. Anat Embryol 153:9–21
Pachter BR, Eberstein A, Goodgold J (1982) Electrical stimulation effect on denervated skeletal myofibers in rats: a light and electron microscopic study. Arch Phys Med Rehabil 63:427–430
Phelan JN, Gonyea WJ (1997) Effect of radiation on satellite cell activity and protein expression in overloaded mammalian skeletal muscle. Anat Rec 247:179–188
Rando TA, Blau HM (1994) Primary mouse myoblast purification, characterization, and transplantation for cell-mediated gene therapy. J Cell Biol 125:1275–1287
Rodrigues ADC, Schmalbruch H (1995) Satellite cells and myonuclei in long-term denervated rat muscles. Anat Rec 243:430–437
Rosenblatt JD, Parry DJ (1992) Gamma irradiation prevents compensatory hypertrophy of overloaded mouse extensor digitorum longus muscle. J Appl Physiol 73:2538–2543
Saitoh A, Okumoto T, Nakano H, Wada M, Katsuta S (1999) Age effect on expression of myosin heavy and light chain isoforms in suspended rat soleus muscle. J Appl Physiol 86:1483–1489
Sakuma K, Watanabe K, Totsuka T, Uramoto I, Sano M, Sakamoto K (1998) Differential adaptations of insulin-like growth factor-I, basic fibroblast growth factor, and leukemia inhibitory factor in the plantaris muscle of rats by mechanical overloading: an immunohistochemical study. Acta Neuropathol 95:123–130
Sakuma K, Watanabe K, Sano M, Uramoto I, Totsuka T (2000) Differential adaptation of growth and differentiation factor 8/myostatin, fibroblast growth factor 6 and leukemia inhibitory factor in overloaded, regenerating and denervated rat muscles. Biochim Biophys Acta 1497:77–88
Schmalbruch H, Lewis DM (2000) Dynamics of nuclei of muscle fibers and connective tissue cells in normal and denervated rat muscles. Muscle Nerve 23:617–626
Schmalbruch H, Al-Amood WS, Lewis DM (1991) Morphology of long-term denervated rat soleus muscle and the effect of chronic electrical stimulation. J Physiol (Lond) 441:233–241
Schultz E, McCormick KM (1994) Skeletal muscle satellite cells. Rev Physiol Biochem Pharmacol 123:213–257
Snow MH (1990) Satellite cell response in rat soleus muscle undergoing hypertrophy due to surgical ablation of synergists. Anat Rec 227:437–446
Spector SA (1985) Trophic effects on the contractile and histochemical properties of rat soleus muscle. J Neurosci 5:2189–2196
Szebenyi G, Fallon JF (1999) Fibroblast growth factors as multifunctional signaling factors. Int Rev Cytol 185:45–106
Tatsumi R, Sheehan SM, Iwasaki H, Hattori A, Allen RE (2001) Mechanical stretch induces activation of skeletal muscle satellite cells in vitro. Exp Cell Res 267:107–114
Timson BF (1990) Evaluation of animal models for the study of exercise-induced muscle enlargement. J Appl Physiol 69:1935–1945
Vandenburgh HH, Hatfaludy S, Karlisch P, Shansky J (1989) Skeletal muscle growth is stimulated by intermittent stretch-relaxation in tissue culture. Am J Physiol 256:C674–C382
Viguie CA, Lu D-X, Huang S-K, Rengen H, Carlson RM (1997) Quantitative study of the effects of long-term denervation on the extensor digitorum longus muscle of the rat. Anat Rec 248:346–354
Webb SE, Lee KKH, Tang MK, Ede DA (1997) Fibroblast growth factors 2 and 4 stimulate migration of mouse embryonic limb myogenic cells. Dev Dyn 209:206–216
Winchester PK, Davis ME, Alway SE, Gonyea WJ (1991) Satellite cell activation in the stretch-enlarged anterior latissimus dorsi muscle of the adult quail. Am J Physiol 260:C206–C212
Yamaguchi A, Ikeda Y, Hirai T, Fujikawa T, Morita I (2003) Local changes of IGF-I mRNA, GH receptor mRNA and fiber size in rat plantaris muscle following compensatory overload. Jpn J Physiol 53:53–60
Yang S, Alnaqeeb M, Simpson H, Goldspink G (1997) Changes in muscle fibre type, muscle mass and IGF-I gene expression in rabbit skeletal muscle subjected to stretch. J Anat 190:613–622
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Yamaguchi, A., Ishii, H., Morita, I. et al. mRNA expression of fibroblast growth factors and hepatocyte growth factor in rat plantaris muscle following denervation and compensatory overload. Pflugers Arch - Eur J Physiol 448, 539–546 (2004). https://doi.org/10.1007/s00424-004-1282-5
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00424-004-1282-5