
Abstract
The aim of the present study was to clarify the effect of carbohydrate (CHO) supplementation on moderate and high-intensity endurance exercise in the heat. Eight endurance-trained men [maximal oxygen uptake (VO2max) 59.5±1.6 ml kg−1 bw−1, mean±SE] cycled to exhaustion twice at 60% VO2max and twice at 73% VO2max at an ambient temperature of 35°C. Subjects ingested either a 6.4% maltodextrin solution (CHO) or an artificially flavoured and coloured placebo (PLA). Time to fatigue was significantly greater with CHO in both the 60% and 73% VO2max trials (14.5% and 13.5% improvement, respectively). Heart rate and oxygen uptake (VO2) did not differ at any point between PLA and CHO. Hypoglycaemia was not seen in any condition but plasma glucose concentrations tended to be higher at both intensities when CHO was fed. CHO oxidation rates were similar at 60% VO2max between CHO and PLA. There were no differences between PLA and CHO in the rate of rise of rectal temperatures (T rec) at either intensity but there was a trend for subjects to fatigue at a high temperature when taking CHO. Ratings of perceived exertion (RPE) tended to be lower throughout both CHO trials; this was significant at 80 min and at fatigue at 60% VO2max. It is concluded that supplementation with CHO improves exercise performance in the heat at both moderate and high endurance intensities. In the absence of a clear metabolic explanation, a central effect involving an increased tolerance of rising deep body temperature merits further investigation.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Fatigue occurring during prolonged exercise in cool or temperate conditions is associated with depletion of carbohydrate (CHO) in the liver and muscles [7, 10]. In these circumstances it is not surprising that CHO supplementation has been shown to delay the onset of fatigue and maintain a high rate of CHO oxidation during the latter stages of the exercise [7, 10, 11]. With exercise at 60–70% VO2max, undertaken in the heat, a limiting factor has been suggested to be the rise in body temperature so that exercise stops after about one hour when body temperature approaches 40°C [17, 32]. In these circumstances changes in blood flow or oxygen extraction by working muscles has been shown not to limit exercise [20]. Furthermore, CHO reserves are not seriously depleted at the point of fatigue during exercise in the heat [31, 35]. This could signify the importance of central, as opposed to peripheral, fatigue. In these circumstances, therefore, it would not be expected that CHO supplementation will have any effect on performance. However, surprisingly, there have been a number of reports that CHO does improve performance in the heat [3, 13, 20, 26, 37] as well as a number reporting no effect of CHO [12, 17, 25].
One of the difficulties in evaluating the published work is that the studies have used different protocols, temperatures and work intensities. One suggestion is that there are compensable and uncompensable exercise heat stress situations [16]. During compensable situations the athlete's thermoregulatory capacity is thought to be sufficient to slow the rise in deep body temperature so that CHO reserves are depleted before a critical body temperature is reached. In contrast, during uncompensable situations the athlete is unable to dissipate heat sufficiently so that body temperature rises to reach a level where motor activity is inhibited by central fatigue before CHO reserves are seriously depleted. There have been no studies in which this idea has been directly tested, using the same subjects in two comparable exercise situations, differing only in the work rate.
The purpose of the work described here was, therefore, to test the idea that with exercise in the heat, subjects working at lower rates would benefit from CHO supplementation but those at higher intensities would be limited by body temperature and therefore not benefit from CHO. Preliminary experiments in which different work loads were investigated showed that with ambient conditions of 35°C and 30% relative humidity (RH), subjects exercised for approximately 2 h at 55% peak power output (PPO). This duration in the heat has been suggested to mark the point at which CHO supplementation would be effective [17]. Exercise at 70% PPO, however, was limited to approximately 1 h. The factors that limit performance at this intensity are believed not to be CHO depletion [3, 23].
It is recognised that dehydration and fluid replacement are important factors determining performance during exercise in the heat [20, 21]. However, in the present study every effort was made to ensure that subjects in both situations were adequately hydrated.
Materials and methods
Subjects
Eight endurance-trained, non-acclimatised, male volunteers gave their written informed consent to participate in the study that was approved by the local Ethics Committee. Their mean age, weight, height and VO2max were 22.6±1.1 (21–24 years), 75.0±7.7 (65.7–86.3 kg), 179.6±8.6 (165–194 cm) and 59.5±4.6 (53–65 ml kg−1 min−1), respectively [mean±SD (range)]. All subjects had previously been involved in studies involving cycle ergometry at this intensity and in similar conditions and were fully familiar with the experimental procedures.
Experimental design
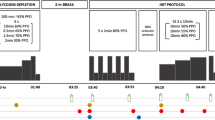
All exercise tests were carried out on an electrically braked cycle ergometer (Lode Excalibur, Groningen, NL) set in the pedal rate independent mode. The protocol consisted of six visits. Visit 1 was an incremental exercise test to determine maximum O2 uptake (VO2max), peak power output (PPO) and the workload to elicit 55% PPO and 70% PPO. These workloads corresponded to 60% and 73% VO2max, respectively, and are referred to as such for the remainder of the manuscript. Visits 2–6 involved exercising to volitional fatigue in a heat chamber maintained at 35°C and an RH of 30%. Visit 2 was an habituation ride, during which subjects exercised at 65% VO2max and consumed water. For the remaining four visits subjects exercise twice at 60% VO2max and twice at 73% VO2max, consuming either a 6.4% maltodextrin solution (CHO) or an artificially flavoured and coloured placebo (PLA). The placebo was identical in taste, content and appearance to the CHO drink, the sole difference being the maltodextrin content (Table 1). The study was carried out in a counterbalanced, double-blind randomised fashion, with each visit separated by an average of 6 days. The order of tests was administered using a Latin-square design.
Visit 1: VO2max
Subjects performed an incremental exercise test to volitional fatigue at a self-selected cadence on a cycle ergometer. The appropriate seat position, handlebar height and orientation were used during testing and replicated in each subsequent visit. The initial workload was 95 W and was increased by 35 W every 3 min until fatigue. Expired gas was recorded continuously throughout (Oxycon Alpha, Jaeger, Germany), as was HR (Polar Accurex Plus, Polar Electro Oy, Finland). PPO was determined according to the method of Kuipers and colleagues [24].
Visits 2–6
Subjects arrived at the laboratory in the morning following an overnight fast, having abstained from exercise, alcohol, caffeine and tobacco for the previous 24 h. To minimise differences in starting muscle glycogen concentrations between visits, subjects were asked to record their diet in the 24-h period before the second visit. This record was copied and returned to the subject with the instruction to follow the same diet before each subsequent visit. Subjects also consumed a 500-ml bolus of water on waking to ensure adequate hydration. On arrival subjects inserted a rectal thermistor 10 cm beyond the anal sphincter before being weighed naked. The subject then rested in a supine position for 30 minutes, during which time a cannula (20-G, Venflon) was inserted into their superficial forearm vein and was kept patent with saline (Baxter) during the test. Two baseline blood samples were taken following the 30-min rest period. Skin thermistors were attached to the subject and the subject drank a bolus (8 ml kg−1, CHO intake 0.51 g kg−1) of either water (visit 2, habituation), CHO (6.4% maltodextrin) or placebo. The subject was then transferred to the heat chamber and cycled to exhaustion at the prescribed workload. The time interval separating bolus consumption and exercise initiation was 5 min. Subjects ingested fluid at a rate of 3 ml kg−1 every 15 min during the ride.
Following the habituation visit the four trials were as follows:
-
1.
Low CHO 60% VO2max with CHO supplementation.
-
2.
Low PLA 60% VO2max with placebo supplementation.
-
3.
High CHO 73% VO2max with CHO supplementation.
-
4.
High PLA 73% VO2max with placebo supplementation.
Subjects wore Lycra shorts and 'clip-in' shoes. In the event of a subject needing to urinate during the test, the subject stopped pedalling and passed urine into a container whilst remaining on the ergometer. A fan was used to circulate the air in the chamber, it was set at the same speed and position between trials. Exercise was stopped either at the subject's volition, their cadence dropping below 20 rpm of their average during the habituation ride or if their rectal temperature (T rec) exceeded 40°C. Following exercise the subject was weighed naked to allow measurement of sweat loss, this was corrected for urinary loss (collected throughout the ride), respiratory loss [38], metabolic loss [28] and fluid intake.
Blood collection
Venous blood samples (8 ml) were collected into EDTA tubes at rest, at 10-min intervals throughout the ride and at the time of fatigue. During blood collection the subjects arm was relaxed and positioned flat across the handlebars. One millilitre of blood was separated and analysed for haemoglobin and haematocrit. The remaining whole blood was centrifuged within 10 min of collection and plasma separated for determination of lactate and glucose concentrations. Samples were stored at 15°C and all assays were completed, in duplicate, within 6 weeks of the exercise.
Blood analysis
Plasma lactate concentrations were determined by spectrophotometry using a quantitative enzymatic (lactate oxidase and peroxidase) method (Sigma Kits, UK). Plasma glucose concentrations were determined by spectrophotometry using a quantitative enzymatic (hexokinase) method (Sigma Kits, UK). Whole blood was used for the quantitative, colorimetric determination of haemoglobin (Sigma Kits, UK) and micro centrifugation was used to determine haematocrit. Percentage changes in plasma volume were calculated from haemoglobin and haematocrit values [14].
Gas analysis
Expired air was analysed for 5 min every 15 min using an automated on-line system (Oxycon Alpha, Jaeger, Germany). CHO oxidation rates for the 60% VO2max trials were calculated from VO2 and VCO2 measures obtained by indirect calorimetry [19]. Indirect calorimetry assumes a respiratory exchange ratio of ≤1.0, therefore calculation of CHO oxidation rates was not possible for the 73% VO2max trial.
Temperature measurement
Ambient temperature during each ride was measured using a wet and dry bulb mercury thermometer (Brannan, UK) and RH calculated from the wet and dry bulb thermometer differential. Thermistors were placed on four skin locations from which mean skin temperature (T msk) was calculated [33]. The rectal and skin thermistors were connected to a Squirrel data logger (Grant Instruments, UK) and values were recorded every 2.5 min and at fatigue.
Perceived exertion, HR and thermal comfort
Rating of perceived exertion (RPE) was recorded every 10 minutes using the 6- to 20-point Borg scale [4] and thermal comfort was recorded simultaneously using the Frank scale [18]. Heart rate was measured continuously by telemetry (Polar Accurex Plus, Polar Electro Oy, Finland).
Data and statistical analysis
Data are reported as mean and standard error (mean±SEM), unless otherwise stated. Differences in time to fatigue were not normally distributed and consequently significance was assessed using the Wilcoxon two-related-samples test. All remaining effect data for both intensities were analysed using a repeated measures ANOVA, with specific differences determined using a paired Student's t test as post hoc.
Data are only reported up to 80 min and fatigue and up to 30 min and fatigue for the 60% VO2max and 73% VO2max trials, respectively. Data points beyond these times did not include all subjects due to variable fatigue. Effect size data (η2) are included together with P values, where appropriate.
Results
60% VO2max trial
An injury (not related to the study) reduced the number of subjects successfully completing the 60% VO2max trial with PLA and CHO to seven. Volitional fatigue was the reason for termination of exercise for all seven subjects during the PLA trial. However, two subjects reached the cut-off temperature (40°C) during the CHO trial.
Exercise time
Six out of the seven subjects were able to exercise for longer with CHO than PLA. PLA time to fatigue was 123.1±13.4 min compared to 145.6±15.1 min for the CHO ride (P=0.028), an average improvement of 14.5±4.9% (Fig. 1).

Individual and average time to fatigue during 60% (above) and 73% (below) maximal oxygen uptake (VO2max) performance trials. The dark line indicates mean change between conditions. Subjects exercised significantly longer on both carbohydrate (CHO) trials
RPE, thermal comfort and cadence
RPE increased throughout the ride with both conditions (Fig. 2a). RPE showed a trend to be lower throughout exercise with CHO, and this was significant at 80 min (P=0.032, η2=0.561) and at the point of fatigue (P=0.012, η2=0.677). Rating of thermal comfort rose steadily during exercise with no difference between PLA and CHO. Cadence was maintained at a steady rate up to 70 min in both conditions (83±2.6 rpm and 86±2.3 rpm for PLA and CHO, respectively) before declining to values at fatigue of 65±2.8 rpm and 71±2.9 rpm for PLA and CHO, respectively. No significant differences were observed between PLA and CHO for cadence at any point (P=0.085, η2=0.414).

Rating of perceived exertion (RPE, Borg scale), plasma glucose and plasma lactate concentrations during the 60% VO2max trials. Filled squares placebo (PLA); open circles CHO. An asterisk indicates significant difference from PLA. Data are mean±SEM, n=7. b RPE, plasma glucose and plasma lactate concentrations during the 73% VO2max trials. Filled squares PLA; open circles CHO. An asterisk indicates significant difference from PLA. Data are mean±SEM, n=8
Physiological measurements
CHO supplementation had no effect on HR response to exercise (Fig. 3a) and sweat rates were not different between PLA and CHO (1261±64 ml h−1 and 1221±68 ml h−1, respectively). Fluid ingested was significantly greater during the CHO trial compared to the PLA (2784±271 ml and 2382±237 ml, respectively; P=0.027, Table 2). Body weight loss (Table 2) was similar between PLA and CHO (471.4±153.9 g and 642.8±235.9 g, respectively), as was urine production (145.0±70.5 ml and 209.3±108.3 ml, respectively). VO2 remained steady during exercise and was similar between PLA and CHO (Table 2). Plasma volume decreased in both PLA and CHO trials (Fig. 4), but this fall was not significantly different (P=0.112, η2=0.367). Plasma glucose concentrations were the same before exercise in the PLA and CHO trials, but were higher at 10 min (P=0.014, η2=0.660), 60 min (P=0.026, η2=0.591) and 70 min (P=0.002, η2=0.811) with CHO compared to PLA (Fig. 2a). Plasma lactate was similar at rest between PLA and CHO and although the concentration increased at the start of exercise in both trials, the response was not significantly different between PLA and CHO (Fig. 2a). Mean CHO oxidation rates were 2.97±0.12 g min−1 and 3.02±0.12 g min−1 for PLA and CHO, respectively. CHO oxidation rates did not change with time and the rates were similar between PLA and CHO (P=0.397, η2=0.122).

% HRmax data during the 60% VO2max trials. Filled squares PLA; open circles CHO. Data are reported as mean+SEM, n=7. b Percentage HRmax data during the 73% VO2max trials. Filled squares PLA; open circles CHO. Data are reported as mean+SEM, n=8

Plasma volume change during all trials. Filled squares PLA; open circles CHO. Data are mean−SEM (n=7 and n=8 for 60% and 73% VO2max, respectively)
Temperature measurements
T rec and T msk responses were similar between PLA and CHO at all time points. T rec increased throughout the exercise until fatigue (Fig. 5a), which occurred at 38.8±0.4°C with PLA and 39.1±0.2°C with CHO (P=0.07, η2=0.477).

Rectal (T rec)and mean skin temperature (T msk) during the 60% VO2max trials. Filled squares PLA; open circles CHO. Data are reported as mean±SEM, n=7. b T rec and Tmsk during the 73% VO2max trials. Filled squares PLA; open circles CHO. Data are reported as mean±SEM, n=8
73% VO2max trial
All eight subjects successfully completed the 73% VO2max trial with PLA and CHO. Volitional fatigue was the reason for termination of exercise for all eight subjects during the PLA trial. However, one subject reached the cut-off temperature (40°C) during the CHO trial.
Exercise time
Six out of the eight subjects were able to exercise for longer with CHO than PLA (Fig. 1). PLA time to fatigue was 50.8±7.5 min compared to 60.6±11.1 min for the CHO ride (P=0.04,). On average CHO resulted in a 13.5±5.5% improvement in exercise time to fatigue compared to PLA.
RPE, thermal comfort and cadence
PE (Fig. 2b) and rating of thermal comfort increased throughout the exercise, but no difference was reported between PLA and CHO trials. Cadence was maintained at a steady rate up to 30 min in both conditions (85±1.8 rpm and 86±1.7 rpm for PLA and CHO, respectively) before declining to values at fatigue of 69±2.9 rpm and 69±3.5 rpm for PLA and CHO, respectively. No significant differences were observed between PLA and CHO for cadence at any point (P=0.506, η2=0.077).
Physiological measurements
CHO supplementation had no effect on HR response to exercise (Fig. 3b) and sweat rates did not differ between PLA and CHO, 1588±131 ml h−1 and 1579±73 ml h−1 respectively. There was no difference in fluid ingested between PLA and CHO conditions (1251±141 ml and 1400±205 ml, respectively; p=0.176, Table 2). Body weight loss (Table 2) was similar between PLA and CHO (385.7±159.5 g and 382.9±192.7 g, respectively), as was urine production (53.6±45.1 ml and 70.7±55.9 ml, respectively). VO2 remained constant with exercise time and was similar between PLA and CHO (Table 2). Plasma volume decreased in both trials (Fig. 4), but this fall was similar between PLA and CHO (P=0.528, η2=0.07) Plasma glucose concentrations were the same in PLA and CHO trials (Fig. 2b), except a higher value was seen at 10 min with CHO compared to PLA (P=0.007, η2=0.734). Plasma lactate was not different at rest between PLA and CHO. The plasma lactate concentration increased at the start of exercise in both trials and continued to do so until the point of fatigue; the response was not different between PLA and CHO (Fig. 2b).
Temperature measurements
T rec and T msk response were similar between PLA and CHO at all time points. T rec increased throughout the exercise until fatigue (Fig. 5b), which occurred at 38.9±0.2°C with PLA and 39.2±0.2°C with CHO (P=0.15, η2=0.312).
Discussion
The objective of the present study was to clarify the effect of exogenous CHO on endurance exercise capacity in the heat. For this purpose two separate exercise intensities were used, one of which was at a workload (60% VO2max) such that subjects could continue for about 2 h and where depletion of CHO stores was thought to be a limiting factor. The other protocol was at a higher intensity (73% VO2max) where subjects were able to continue for approximately 1 h and the limiting factor was most likely raised body temperature rather than depletion of CHO stores. In these circumstances it would be predicted that CHO supplementation would benefit exercise at 60% VO2max but not at 73% VO2max. Our findings were unexpected, with CHO supplementation improving endurance capacity in both instances. This suggests that the mechanism whereby CHO improves performance is not simply by providing an additional energy supply to working muscles.
Exercise at 60% VO2max
Our observation that CHO supplementation improved endurance by 14.5% (Fig. 1) is in agreement with previous reports [3, 13, 20, 26, 37] where the exercise in the heat was of a similar duration (>2 h). It should be noted that in the CHO trial one subject was stopped at 145 min and another at 147 min because T rec reached 40°C. Had they been allowed to continue to volitional fatigue the average increase in performance with CHO supplementation would have been slightly higher than the 14.5% reported here.
It has been demonstrated previously that exogenous CHO helps maintain blood glucose and high rates of CHO oxidation in the later stages of prolonged exercise when muscle glycogen stores are exhausted [7, 10, 11]. The data from our present study at 60% VO2max, however, provide only limited support for this sequence of events as an explanation for the improved performance. Although significantly higher plasma glucose levels were seen with CHO supplementation in the latter part of the exercise (Fig. 2a), plasma glucose never fell below 4.3 mmol l−1 in the PLA condition. Furthermore, no differences were observed in CHO oxidation between CHO and PLA. CHO oxidation rates were reported for the first 80 min since this was the maximum time for which data on all subjects was available. However, analysis of the three subjects who continued for the longest time, and for whom respiratory data were available between 90 and 120 min, also showed no difference in CHO oxidation rates between CHO and PLA. Although unexpected, our results are similar to those of a previous study which reported no difference in RER values over 90 min of exercise at 70% VO2peak in the heat between a 7% CHO supplementation and PLA [17].
The ingestion of fluid during exercise in the heat is well known to improve performance compared to no fluid intake [3, 20, 41]. It could be argued that some of the improvement seen in the present study was due to differences in fluid intake or absorption. Indeed, there was a trend for plasma volume to decrease with CHO supplementation (non-significant) and this could indicate reduced gastric emptying. It has been shown that gastric emptying is slowed by a relatively high CHO content of a drink [9, 17, 39]. However, if fluid delivery was important in the present study the performance effect would have been in favour of the PLA condition. In addition, there were no differences in thermoregulation, T rec and T msk being very similar at all times between PLA and CHO conditions (Fig. 5a). The only difference, although this did not reach significance, was that T rec at fatigue was 0.3°C higher following CHO ingestion compared to PLA. The higher temperature at fatigue in the CHO conditions can be explained by the longer time spent exercising to reach this point.
Exercise at 73% VO2max
The results obtained with CHO supplementation while exercising at 73% VO2max did not support the prediction of Febbraio et al. [17] that supplementation would only be effective with exercise lasting more than 2 h. Our observation that time to fatigue was extended by approximately 10 min or 13.5% with CHO compared to PLA (Fig. 1), was very similar to the improvement seen with CHO in the low-intensity trials. The improvement could have been slightly greater if one of the subjects had not been withdrawn from the study after 55 min during the CHO trial when his T rec reached 40°C.
There have been previous reports of CHO supplementation improving performance in the heat during exercise lasting up to 1 h. Below et al. [3] demonstrated that a 10-min performance ride following 50 min of exercise at 80% VO2max in the heat was completed fastest with subjects taking CHO and fluid compared to either fluid or CHO alone. In another study, the time taken to run 15 km in the heat (75–78% VO2max) was found to be significantly shorter with ingestion of 6% and 8% CHO drinks compared to water [27]. Similar improvements with CHO have also been reported in high-intensity exercise in thermo-neutral conditions where it was thought unlikely that CHO supplementation would be beneficial [1, 2, 15, 23, 30].
In the present study, plasma glucose concentrations were marginally higher in the CHO condition. However, in both cases there was a gradual rise during the exercise with highest values being obtained at the time of fatigue. Hypoglycaemia, as with the 60% VO2max trial, was not a cause of fatigue in the PLA condition and, likewise, CHO supplementation did not exert its effect by preventing hypoglycaemia.
Despite the fact that a number of studies have now found CHO supplementation to be of benefit in conditions where CHO stores would not be considered limiting, the mechanism responsible remains to be established. A higher rate of CHO oxidation has been suggested with CHO ingestion [30]. However, in that study no differences were reported for CHO oxidation rate for the first 45 min of exercise at 77% VO2max. This is supported by previous reports of RER measurements indicating no difference in CHO oxidation between CHO and PLA conditions during exercise at comparable intensities, both in the heat and in thermo-neutral environments [3, 15, 27]. This is consistent with the view that endogenous CHO is not limiting for exercise lasting up to 1 h. It has been suggested that at 60–70% VO2max only 10–20% of ingested CHO is oxidised in the 1st hour of exercise [23, 40].
During exercise at 73% VO2max, which was close to the lactate threshold (steady blood lactates of 4–5 mmol l−1, Fig. 2b), it was not possible to measure CHO oxidation by indirect calorimetry because RER values were greater than 1.0. Although we have no measure of CHO oxidation the fact that plasma glucose levels were maintained, and even elevated, indicates that there was no limitation to CHO oxidation in terms of supply of blood borne glucose in the PLA condition. Therefore, improvements in performance in the CHO condition were unlikely to have been due to the greater availability of blood glucose sustaining high levels of CHO oxidation. The increase of plasma glucose above baseline towards the end of exercise has been noted by others [3, 8, 27].
Causes of fatigue during submaximal exercise
The reason for fatigue in both the moderate and high-intensity trials is not immediately apparent. As has been discussed above there is no evidence that CHO oxidation was ever limited by substrate depletion or that CHO supplementation helped maintain high oxidation rates. There were no conventional indications of peripheral fatigue as a consequence of either workload. Hypoglycaemia was never apparent in any of the conditions. Plasma lactate concentrations during exercise at 60% VO2max were low and constant throughout the exercise with no rapid increase towards fatigue which might have indicated a reduction of peripheral oxygen supply. Although plasma lactate was higher during exercise at 73% VO2max, the values were relatively constant during the exercise and, again, there was no precipitate rise towards the point of fatigue. Increased cardiovascular strain is recognised as an important factor in hyperthermic exercise but there was no evidence that it was either the cause of fatigue or differed between PLA and CHO conditions. Heart rate, ventilation and VO2, never reached limiting values in any condition (Fig. 3a, b, Table 2) nor was there any indication of significant dehydration as assessed by change in plasma volume (Fig. 4).
Since there is no evidence of peripheral fatigue in the present study some central mechanism must be considered. Nielsen and colleagues have suggested that there is a critical core temperature that limits exercise in the heat [32]. These authors found that during exercise at 60% VO2max at an ambient temperature of 40°C, fatigue occurred at the same oesophageal temperature (39.7±0.15°C) independent of acclimation status and duration of exercise. Body temperature has also been reported by several other authors as a major factor limiting exercise in the heat at workloads between 60% and 70% VO2max, [17, 22, 35, 36]. The implication of this work is that a high body temperature results in an inhibition of motor activity [6, 32, 34], presumably with the effect of reducing heat production and thus preventing temperature reaching dangerously high levels. The mechanism by which body temperature might inhibit motor output is not known but may involve the hypothalamus. Hypothalamic activity, as monitored by the release of prolactin, increases towards the point of fatigue and is closely associated with increases in body temperature [5].
In our present study, T rec at the point of fatigue in the PLA condition was 38.8±0.6 in the 60% VO2max trial and 38.9±0.5 in the 73% VO2max trial, values that were not statistically different (P=0.455). This is consistent with the notion that a high body temperature is an important factor limiting performance. Furthermore, it suggests that at both workloads in the present study the limiting factor was a rise in body temperature rather than any metabolic limitation. Exercise continued for longer at both intensities in the CHO condition and although it did not achieve statistical significance, T rec was, on average, 0.3°C higher than at the point of fatigue in the PLA condition. In terms of the critical temperature theory of fatigue, we would suggest that in the present study CHO had the effect of increasing the subjects' tolerance of raised body temperature. It is a matter of pure speculation as to how CHO supplementation might affect heat tolerance and central fatigue but there are two main possibilities. The first is that the increased blood glucose levels in the CHO condition, while not affecting muscle metabolism, did improve the metabolic state of the central nervous system and higher brain centres concerned with motivation and motor output. Alternatively, afferent nerve endings in the mouth or GI tract might detect the CHO and influence the function of the hypothalamus or other centres [23]. One way of distinguishing between these alternatives would be to infuse CHO to avoid stimulating receptors in the gastrointestinal tract.
In conclusion, we have found that supplementation with CHO delays fatigue during exercise at 60% VO2max in the heat. It had been anticipated that at this workload subjects would be limited by CHO availability but despite the fact that exercise continued for about 2 h, the evidence suggests that fatigue was most likely due to the attainment of a high core temperature. In the circumstances it is therefore surprising that CHO supplementation improved performance. At 73% VO2max it was expected that temperature would be the limiting factor, and that CHO supplementation would not affect performance. However, as with exercise at the lower intensity, CHO did improve performance and therefore the improvement with CHO was intensity independent. It is concluded that in both situations the additional CHO probably affected performance through a central rather than a peripheral mechanism. The possible central actions of CHO improving performance during endurance exercise warrant further investigation.
References
Anantaraman R, Carmines AA, Gaesser GA, Weltman A (1995) Effects of carbohydrate supplementation on performance during 1 hour of high intensity exercise. Int J Sports Med 16:461–465
Ball TC, Headley SA, Vanderburgh PM, Smith JC (1995) Periodic carbohydrate replacement during 50 min of high-intensity cycling improves subsequent sprint performance. Int J Sport Nutr 5:151–158
Below PR, Mora-Rodriguez R, Gonzalez-Alonso J, Coyle E (1995) Fluid and carbohydrate ingestion independently improve performance during 1 h of intense exercise. Med Sci Sports Exerc 27:200–210
Borg GAV (1982) Psychophysical bases of perceived exertion. Med Sci Sports Exerc 14:377–381
Brisson GR, Peronnet F, Perrault H, Boisvert P, Massicotte D, Gareau R (1991) Prolactinotrophic effect of endogenous and exogenous heat loads in human male adults. J Appl Physiol 70:1351–1355
Bruck K, Olschewski H (1987) Body temperature related factors diminishing the drive to exercise. Can J Physiol Pharmacol 65:1274–1280
Coggan AR, Coyle EF (1987) Reversal of fatigue during prolonged exercise by carbohydrate infusion or ingestion. J Appl Physiol 63:2388–2395
Coggan AR, Coyle EF (1988) Effect of carbohydrate feedings during high-intensity exercise. J Appl Physiol 65:1703–1709
Costill DL, Saltin B (1974) Factors limiting gastric emptying during rest and exercise. J Appl Physiol 37:679–683
Coyle EF, Hagberg JM, Hurley BF, Martin WH, Ehsani AA, Holloszy JO (1983) Carbohydrate feeding during prolonged strenuous exercise can delay fatigue. J Appl Physiol 55:230–235
Coyle EF, Coggan AR, Hemmert MK, Ivy JL (1986) Muscle glycogen utilisation during prolonged strenuous exercise when fed carbohydrate. J Appl Physiol 61:165–172
Davis JM, Burgess WA, Slentz CA, Bartoli WP, Pate RR (1988) Effects of ingesting 6% and 12% glucose/electrolyte beverages during prolonged intermittent cycling in the heat. Eur J Appl Physiol 57:563–569
Davis JM, Lamb DR, Pate RR, Slentz CA, Burgess WA, Bartoli WP (1988) Carbohydrate-electrolyte drinks: effects on endurance cycling in the heat. Am J Clin Nutr 48:1023–1030
Dill DB, Costill DL (1974) Calculation of percentage changes in volumes of blood, plasma, and red cells in dehydration. J Appl Physiol 37:247–248
El-Sayed MS, Balmer J, Rattu AJM (1997) Carbohydrate ingestion improves endurance performance during a 1-h simulated cycling time trial. J Sports Sci 15:223–230
Febbraio MA (1999) Temperature, muscle metabolism and performance. In: Lamb DR, Murray R (eds) Perspectives in exercise science and sports medicine: the metabolic basis of performance in exercise and sport. Cooper, Carmel, pp 315–353
Febbraio MA, Murton P, Selig SE, Clark SA, Lambert DL, Angus DJ, Carey MF (1996) Effect of CHO ingestion on exercise metabolism and performance in different ambient temperatures. Med Sci Sports Exerc 28:1380–1387
Frank SM, Raja SR, Bulcao CF, Goldstein DS (1999) Relative contribution of core and cutaneous temperatures to thermal comfort and autonomic responses in humans. J Appl Physiol 86:1588–1593
Frayn KN (1983) Calculation of substrate oxidation rates in vivo from gaseous exchange. J Appl Physiol 55:628–634
Galloway SD, Maughan RJ (2000) The effects of substrate and fluid provision on thermoregulatory provision on thermoregulatory and metabolic responses to prolonged exercise in a hot environment. J Sports Sci 18:339–351
Gonzalez-Alonso J, Calbet JAL, Nielsen B (1998) Muscle blood flow is reduced with dehydration during prolonged exercise in humans. J Physiol (Lond) 513:895–905
Gonzalez-Alonso J, Teller C, Andersen SL, Jensen FB, Hyldig T, Nielsen B (1999) Influence of body temperature on the development of fatigue during prolonged exercise in the heat. J Appl Physiol 86:1032–1039
Jeukendrup A, Brouns F, Wagenmakers AJM, Saris WHM (1997) Carbohydrate-electrolyte feedings improve 1-h time trial cycling performance. Int J Sports Med 18:125–129
Kuipers H, Verstappen FTJ, Keizer HA, Geurten P, Van Kranenburg G (1985) Variability of aerobic performance in the laboratory and its physiologic correlates. Int J Sports Med 6:197–201
Millard-Stafford M, Starling PB, Rosskopf LB, Hinson BT, Dicarlo LJ (1990) Carbohydrate-electrolyte replacement during a simulated triathlon in the heat. Med Sci Sports Exerc 22:621–628
Millard-Stafford ML, Starling PB, Rosskopf LB, Dicarlo LJ (1992) Carbohydrate-electrolyte replacement improves distance running performance in the heat. Med Sci Sports Exerc 24:934–940
Millard-Stafford M, Rosskopf LB, Snow TK, Hinson BT (1997) Water versus carbohydrate-electrolyte ingestion before and during a 15-km run in the heat. Int J Sport Nutr 7:26–38
Mitchell JW, Nadel ER, Stolwijk JAJ (1972) Respiratory weight losses during exercise. J Appl Physiol 32:474–476
Murray R, Eddy DE, Murray TW, Seifert JG, Paul GL, Halaby GA (1987) The effect of fluid and carbohydrate feedings during intermittent cycling exercise. Med Sci Sports Exerc 19:597–604
Neuffer PD, Costill DL, Flynn MG, Kirwan JP, Mitchell JB, Houmard J (1987) Improvements in exercise performance: effects of carbohydrate feedings and diet. J Appl Physiol 62:983–988
Nielsen B, Savard G, Richter EA, Hargreaves M, Saltin B (1990) Muscle blood flow and muscle metabolism during exercise and heat stress. J Appl Physiol 69:1040–1046
Nielsen B, Hales JRS, Strange S, Juel Christensen N, Warberg J, Saltin B (1993) Human circulatory and thermoregulatory adaptations with heat acclimation and exercise in a hot, dry environment. J Physiol (Lond) 460:467–485
Nielsen R, Nielsen B (1984) Measurement of mean skin temperature of clothed persons in cool environments. Eur J Appl Physiol 53:231–236
Nybo L, Nielsen B (2001) Perceived exertion is associated with an altered brain activity during exercise with progressive hyperthermia. J Appl Physiol 91:2017–2023
Parkin JM, Carey MF, Zhao S, Febbraio MA (1999) Effect of ambient temperature on human skeletal muscle metabolism during fatiguing submaximal exercise. J Appl Physiol 86:902–908
Pitsiladis YP, Maughan RJ (1999) The effects of exercise and diet manipulation on the capacity to perform prolonged exercise in the heat and in the cold in trained humans. J Physiol (Lond) 517:919–930
Pitts GC, Johnson RE, Consolazio FC (1944) Work in the heat as affected by intake of water, salt and glucose. Am J Physiol 142:253–259
Snellen JW (1966) Mean body temperature and the control of thermal sweating. Acta Physiol Pharmacol Neerl 14:99–174
Vist GE, Maughan RJ (1995) The effect of osmolality and carbohydrate content on the rate of gastric emptying of liquids in man. J Physiol (Lond) 486:523–531
Wagenmakers AJM, Brouns F, Saris WHM, Halliday D (1993) Oxidation rates of orally ingested carbohydrates during prolonged exercise in men. J Appl Physiol 75:2774–2780
Walsh RM, Noakes TD, Hawley JA, Dennis SC (1994) Impaired high-intensity cycling performance time at low levels of dehydration. Int J Sports Med 15:392–398
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Carter, J., Jeukendrup, A.E., Mundel, T. et al. Carbohydrate supplementation improves moderate and high-intensity exercise in the heat. Pflugers Arch - Eur J Physiol 446, 211–219 (2003). https://doi.org/10.1007/s00424-003-1020-4
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00424-003-1020-4