
Abstract
The aim of this paper is to integrate different bodies of research including brain traveling waves, brain neuromodulation, neural field modeling and post-stroke language disorders in order to explore the opportunity of implementing model-guided, cortical neuromodulation for the treatment of post-stroke aphasia. Worldwide according to WHO, strokes are the second leading cause of death and the third leading cause of disability. In ischemic stroke, there is not enough blood supply to provide enough oxygen and nutrients to parts of the brain, while in hemorrhagic stroke, there is bleeding within the enclosed cranial cavity. The present paper focuses on ischemic stroke. We first review accumulating observations of traveling waves occurring spontaneously or triggered by external stimuli in healthy subjects as well as in patients with brain disorders. We examine the putative functions of these waves and focus on post-stroke aphasia observed when brain language networks become fragmented and/or partly silent, thus perturbing the progression of traveling waves across perilesional areas. Secondly, we focus on a simplified model based on the current literature in the field and describe cortical traveling wave dynamics and their modulation. This model uses a biophysically realistic integro-differential equation describing spatially distributed and synaptically coupled neural networks producing traveling wave solutions. The model is used to calculate wave parameters (speed, amplitude and/or frequency) and to guide the reconstruction of the perturbed wave. A stimulation term is included in the model to restore wave propagation to a reasonably good level. Thirdly, we examine various issues related to the implementation model-guided neuromodulation in the treatment of post-stroke aphasia given that closed-loop invasive brain stimulation studies have recently produced encouraging results. Finally, we suggest that modulating traveling waves by acting selectively and dynamically across space and time to facilitate wave propagation is a promising therapeutic strategy especially at a time when a new generation of closed-loop cortical stimulation systems is about to arrive on the market.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
1 Introduction
Over the last few years, evidence for the presence of traveling waves (TWs) occurring both naturally and pathologically in the human brain has been rapidly accumulating (Rubino et al. 2006; Wu et al. 2008; Deco et al. 2008; Takahashi et al. 2011; Sato et al. 2012; Heitmann et al. 2013; Breakspear 2017; Salimpour and Anderson 2019; Roberts et al. 2019). A periodic TW is an oscillatory solution moving with constant shape and speed, which is periodic as a function of time and space and propagates like a stadium wave. Propagating waves have been observed during most types of cortical processing examined such as vision, olfaction and sleep and using various recording modalities. Overall, there is a large consensus in the scientific literature indicating that TWs are ubiquitous in the brain and appear to coordinate distant functional brain regions involved in cognitive functions including executive functions, attention, memory, language, visuospatial abilities, orientation, etc. Cortical TWs reflect functional information originating from both hemispheres, traveling short and long distances, associated with various time delays. TWs cause subthreshold depolarization of the neuronal membrane of about 5–10 mV from resting potential and increase the spiking probability. These spikes in turn “depolarize more postsynaptic neurons in the neighboring area to sustain wave propagation” (Page 2) (Wu et al. 2008).
TWs are observed at the cortical surface as well as in deeper, subcortical structures (e.g., basal ganglia and thalamus) in healthy subjects (Canolty and Knight 2010) and in subjects with neurological disorders such as hallucinations (Brasic 1998), migraine (Dahlem and Chronicle 2004), epilepsy (Lopes Da Silva et al. 2003), tinnitus (Chrostowski et al. 2011) and cortical spreading depression following stroke (Whalen et al. 2018). According to Muller et al. (2018), these waves “travel over spatial scales that range from the mesoscopic (single cortical areas and millimeters of cortex) to the macroscopic (global patterns of activity over several centimeters) and extend over temporal scales from tens to hundreds of milliseconds” (Page 2).These authors noted that cortical TW can occur spontaneously or may be evoked by external stimuli. The propagation of cortical mesoscopic waves depends on axonal conduction delay of horizontal U-fibers located in superficial cortical layers, and their amplitude decays over distance (Muller et al. 2018). Clearly, time delays play an important role in the unfolding of these waves (Wu et al. 2008), but see also Alexander et al. (2016). However, the transition between mesoscopic and macroscopic scales must be considered with caution in the sense that networks are distributed (hodotopic) and the key element here may not be expressed in terms of distance but rather in terms of spatial frequency of the oscillations.
TWs have been observed across multiple regions, directions and frequencies in individual subjects at the single-trial level. Recently, Zhang et al. (2018) examined direct brain electrocorticographic recordings (ECoG) from neurosurgical patients performing a working memory task. Most patients had clusters of electrodes that showed the same frequency of oscillations across space (Fig. 1). They found a total of 208 oscillation clusters in the 77 patients within 2–15 Hz (alpha and theta). Clusters at given electrodes within one patient did not necessarily have the same frequency as the (spatially) identical clusters in another patient, suggesting that neuronal oscillations vary from individual to individual and are difficult to capture with typical group-average analyses (Zhang et al. 2018). TWs are subject dependent and “clusters of ECoG electrodes with narrowband oscillations at the same frequency reflected robust within-subject spatial frequency clustering” (Page 3, Zhang et al. 2018). These oscillatory clusters displayed spatial phase gradients, indicating that they formed TW that propagated at about 0.25–0.75 m/s. In this study, TWs correlate with the subject’s performance, propagate in specific directions and synchronize distributed cortical networks that are communicating. TWs guide the spatial propagation of neural activity with their direction possibly induced by an intrinsic frequency gradient (Zhang et al. 2018).

Adapted from Zhang et al. (2018) (with permission)
TW in the human neocortex recorded in patient #1. The authors analyzed electrocorticographic (ECoG) brain recordings from 77 neurosurgical patients. Patient #1 is a 17-year-old right-handed male. The grid of electrodes was positioned over the temporal and occipital cortex of the left hemisphere. The authors analyzed the data with a new technique that identifies TW at the single trial level across frequencies and electrodes configurations. TWs were observed in 84% of the patients examined (65 out of 77). In patient#1, TWs were present across a wide frequency range (2–15 Hz). Zhang et al. show that the observed theta and alpha bands were relevant behaviorally, as their propagation correlated with subject performance and events in a memory task they had to perform. In patient #1, clusters of significant TW were observed in the left temporal cortex in α (8–15 Hz). Top: raw signals for 4 s of one trial from three selected electrodes. The selected electrodes are ordered from anterior (top) to posterior (bottom). Middle: a 500-ms zoomed version of the signals from the top panel. Bottom: signals filtered at 6–10 Hz. The frequency is around 8.3 Hz. The color of each electrode indicates the relative phase.
TWs vary across subjects, but they tend to have a consistent propagation direction across multiple trials and have been observed across all lobes of the telencephalon. However, directions of wave propagation also depend on the methodology used to record the data as explained by Muller et al. (2018). They are also correlated with performance on the working memory task. In the study by Zhang et al. (2018), the direction of TW was most consistent in the frontal and temporal lobes, and most waves demonstrated a posterior-to-anterior directionality in these regions but direction varied more in the parietal and occipital lobes.
These authors found that directional consistency was higher in the frontal and temporal lobes after the retrieval cue onset (where subjects were required to recall previously shown stimuli) in the working memory task. As mentioned above, directional consistency was also positively correlated with performance, suggesting that waves traveling in a consistent direction were associated with better working memory efficiency (Zhang et al. 2018). In Heitmann et al. (2013), the authors hypothesize that the cortex utilizes the direction and wavelength of TW to encode information which may be decoded “by the spatial arrangement of receptors within the dendritic receptor field” (Page 1) and they show “how the density distributions of excitatory and inhibitory receptors can combine to act as a spatial filter of wave patterns” (Page 1).
Surprisingly, TWs have been artificially induced using transcranial electric stimulation with alternating currents (tACS). Alekseichuk et al. (2016) used tACS protocols with multiple electrodes and phase-shifted stimulation currents to alter the functional connectivity between distant brain regions. The experiment was performed in two anesthetized nonhuman primates. The authors observed an increase in coordination between two remote regions during an in-phase stimulation and conversely an increase in disorganization during an anti-phase stimulation. These authors show that “the location of the electric field maximum changes over the stimulation cycle” (Page 1 Alekseichuk et al. 2018). These results obtained in animals are not necessarily predictive of outcomes in human patients and need to be confirmed, but they open up the possibility for developing noninvasive stimulation protocols with high definition tACS (see Turi et al. 2018 on working memory).
The experimental findings reported above raise several questions including: What are the functional roles of TW in brain information processing? What is the dynamics of TW supporting the production of smooth and efficient human behaviors? How are TWs affected by brain disorders and especially in post-stroke aphasia? Thus, the aim of the paper is to capture recent updates on cortical TW and neural field modeling with the long-term objective of implementing closed-loop, model-guided, cortical neuromodulation as a new therapeutic solution for post-stroke aphasia. Therefore, we first connect experimental, clinical and theoretical observations and we voluntarily stand at the interface between different fields including neurosurgery, neurology, electrophysiology, neuroimagery, mathematics and physics. This prospect paper is organized as follows: We examine TW in general and specifically in aphasia research, we present a neural field model inspired from the current literature as a way to describe the distribution of the electric potential in the cortex, and finally, we discuss some of the issues that need to be addressed before closed-loop model-guided neuromodulation becomes a reality.
2 Functional role and dynamics of cortical TW
As indicated above, TWs, i.e., the propagation of electrical activity at the surface of the cortex, are observed during most types of cortical processing. TWs are thought to serve important functional roles in computation and communication between brain subnetworks. It has been proposed that TW associated with neuronal oscillations can organize spatial phase distributions in populations of neurons. Indeed, far from being an epiphenomenon or a local phenomenon, TWs coordinate neural processing across large contiguous areas of human cortex to allow rapid and efficient communication (Bahramisharif et al. 2013; Zhang et al. 2018). Wu et al. (2008) suggest that “propagating waves reflect the organization of neuronal activity, within which millions of neurons work together to carry out a function” (Page 487). Patten et al. (2012) indicate that “propagating waves of brain oscillations might be involved in mediating long-range interactions between widely distributed parts of the human cortex” (Page 1). Rubino et al. (2006) confirmed an observation, made earlier by Ermentrout and Kleinfeld (2001), suggesting that propagating waves mediate information transfer and increase the network gain for incoming stimulation in the cortex. For Muller et al. (2018), TWs link high precision information with information from a broader contextual content and propagate over multiple functional regions, in the direction of maximum information transfer (some kind of gain adjustment). In the words of Turi et al. (2018), “network oscillations, the aggregate oscillatory activity of a large number of neural assemblies, provide the substrate for information processing and facilitate the local-to-global communication crucial for cognition” (Page 1, Turi et al. 2018). Overall, TWs coordinate activity between brain regions and allow constant updating of cortical activities. In their recent paper, Roberts et al. (2019) show that the human connectome “supports the spontaneous emergence of complex metastable brain waves” (Page 13). These TWs are “robust to changes in model parameters, the choice of model, and the thresholding of the underlying connectome. They reproduce across connectome datasets and yield qualitative matches with empirical resting-state data. The widespread empirical observation of TW phenomena reproduced suggests a range of functional and potentially pathological roles in cognition and states of awareness” (Page 13). For a recent discussion of TW functional role, see also Alamia and van Rullen (2019).
Alexander et al. (2016) studied TW using whole head noninvasive magnetoencephalographic recordings (MEG) during a task in which 20 healthy subjects were engaged in an audio–visual perceptual task. They measured unbiased estimates of TW velocities and observed that TWs in MEG have velocities closely tied to temporal frequency: “The higher the temporal frequency, the higher the velocity” (Page 20). The authors concluded that TW may index processes involved in global coordination of cortical activity. An important factor to consider when measuring propagation speed of TW is that it varies depending on the methodology used. When macroscopic waves are recorded from EEG or from ECoG, which have low spatial and high temporal resolutions, the propagation speed varies between 1 and 10 m/s. As indicated by Muller et al. (2018), these results are compatible with the range of axonal conduction speeds of myelinated white matter tracts (WMTs) in the cortex. However, when measuring mesoscopic wave propagation speed using local field potential from multielectrode arrays or from optical imaging signals recorded with voltage-sensitive dyes having high spatial and temporal resolution, the propagation speed varies from 0.1 to 0.8 m/s (Muller et al. 2018). This result is “consistent with the axonal conduction speed of the unmyelinated long-range horizontal fibers within the superficial layers of the cortex” (Page 3, Muller et al. 2018). These observations will have significant repercussions when calculating time delays.
It is important to note that, while the potential causal role of TW is not entirely elucidated yet, results from recent studies are leading to three remarks.
A first remark is that TWs related to cortical functional connectivity and signal propagation across large brain networks represent a dynamic phenomenon. Thus, linking the connectome and the “dynome”, as suggested by Kopell et al. (2014), is a necessity. Dynamics (the evolution in time) of activity of populations of neurons is crucial for understanding cortical function. Thus, including the dynamic characteristics of TW occurring between nodes or hubs in new therapeutic strategies will help develop more selective and more precise interventions to influence brain rhythms in human subjects. Indeed, in a review paper on dynamic models of large-scale brain activity, Breakspear (2017) presents evidence supporting the view that “collective, nonlinear dynamics are central to adaptive cortical activity” (Page 340) and suggests that “aberrant dynamic processes appear to underlie a number of brain disorders” (Page 340).
A second remark is that TWs have been described and modeled for years. Indeed, modeling TW in precise mathematical terms is not new (Amari 1977; Jirsa and Haken 1996; Robinson et al. 1997; Deco et al. 2008; Coombes et al. 2014; Senk et al. 2018). There is a large body of evidence showing that integro-differential reaction–diffusion equations can be used to model the propagation of waves across the cortex (Wilson and Cowan 1973; Ermentrout and McLeod 1993; Atasoy et al. 2016). Mean field modeling describes the behavior of the collective action of neuron populations. They are used for example to model epilepsy, sleep, anesthesia, migraine, degenerative brain disorders and stroke. Connections are mediated by cortical WMT running just below the gray matter when relating nearby cortical areas or deeper when relating more distant cortical areas (Muller et al. 2018). Connecting nearby or more distant cortical areas will affect axonal conduction delays. This modeling approach is illustrated in Sect. 4.
A third remark is that TWs appear to be related to amplitude and phase modulation. Cross-frequency coupling (CFC), the association of multiple frequency neural oscillations, coordinates slower and faster neural rhythms across spatial and temporal scales (Canolty and Knight 2010; Florin and Baillet 2015). CFC is relevant to understand brain function under healthy or diseased conditions. However, it is a complex concept studied using a variety of methodologies and has fundamental caveats and confounds (Canolty and Knight 2010; Aru et al. 2015; Hyafil 2015). Therefore, it must be used with caution. For Canolty and Knight (2010), “CFC may thus serve as a mechanism to transfer information from large-scale brain networks operating at behavioral timescales to the fast, local cortical processing required for effective computation and synaptic modification, thus integrating functional systems across multiple spatiotemporal scales” (Page 506). With their model, Zhang et al. (2018) were able to predict the direction and speed of the TW, determined both by the strength of local phase coupling and by the spatial gradients of intrinsic oscillation frequencies. In CFC, the local oscillatory phase is coupled to the simultaneous amplitude of neuronal spiking and to high-frequency oscillations. If individual regions simultaneously exhibit both TW as well as CFC, it indicates that bands of neuronal spiking activity propagate across the cortex during behavior. Turi et al. (2018) indicate that “The best-known type of such coupling is the phase-amplitude cross-frequency coupling, in which the phase of lower frequency oscillations (e.g., the theta rhythm) correlates with the amplitude of higher frequencies (e.g., the gamma rhythm)” (Page 1). Through such mechanisms, Turi et al. (2018) showed that it is possible not only to observe but also to modulate brain rhythms in humans via brain stimulation techniques such as tACS. For a general approach to coupling principles, see Klimesch (2018).
The relationship between TW and language/aphasia is not entirely elucidated yet. A change in high gamma bursts activity/propagation appears to be associated with language symptoms. Nishida et al. (2017) have shown a high gamma activity (70–110 Hz) in language cortical sites (including the superior-temporal gyrus and precentral and inferior-frontal gyri) of [nonaphasic] epileptic patients listening to a tone or reproducing this tone by humming. Subsequent high-frequency electrical stimulation of these high gamma sites produced “speech arrest, inability to control vocalization and forced vocalization” (Page 2). High-frequency electrical stimulation is known to act as a virtual and transient functional lesion, with a high spatial and temporal resolution. The symptoms reported by the authors were language perturbations similar to those encountered in expression aphasia. Indeed, the interruption of vocalization is reminiscent of speech arrest, the inability to control vocalization is reminiscent of dysarthria, and in both cases, it affects motor planning (i.e., a disorder of the articulatory loop). In another study by Tamura et al. (2016) similar results were observed, i.e., a correspondence between language cortical sites identified by gamma hyperactivity and speech disorders evoked by cortical electrical stimulation. In other words, functional inhibition induced by high-frequency electrical stimulation of high activity gamma sites produces aphasia symptoms.
Following a stroke electrical activity is perturbed in perilesional tissue. Rabiller et al. (2015) write “the EEG changes observed after ischemia are caused by an electrical impairment of the neurons due to the changes of the membrane potential induced by energy deprivation. This energy deprivation results from the reduction of the CBF [cerebral blood flow] and leads to irreversible neuronal damages if the CBF is not restored in time. However, the neuronal origin of the increase of slow or δ oscillations and the decrease of high frequency oscillations after stroke is still under debate.”(Page 25618). For Rabiller et al. (2015), “Findings in global EEG changes after stroke coalesce to an increase of slower frequency oscillations and a decrease of faster ones.” (Page 25625) and these authors conclude: “high band frequencies, such as the α, β, and γ oscillations, are associated with awake states or cognitive task engagement, and their presence frequently reduces after stroke.” (Page 25627). Despite these observations, perilesional tissue activation after a stroke is associated “with improved outcome even though this tissue may be hypo-perfused” (Page 4) (Kiran et al. 2012). A recent discussion on the role of high and low frequency bands in short- and long-distance brain communication via top-down and bottom-up processes can be found in Khambhati et al. (2019).
Other pertinent references include Fellinger et al. (2011) who showed that traveling alpha waves can predict visual-semantic (picture) categorization in normal subjects; Alexander et al. (2013), Klimesch et al. (2007) and Burkitt et al. (2000).
As indicated above, the results presented do not constitute complete experimental evidence since they are based on correlations and correlation is not causation. These results reflect current comprehension of these complex phenomena, and we surmise that TW may play a role in human behavior in general and human language in particular.
3 Network approach in post-stroke aphasia
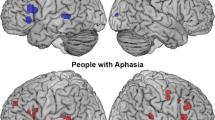
Aphasia is an acquired communication disorder due to brain damage. In a recent review paper, Kiran and Thompson (2019) noted that language recovery after stroke is a nonlinear process which depends on the delay after stroke occurrence. “While it is well documented that the greatest changes in the neural architecture for language occur in early stages of recovery, neuroplasticity occurs even in chronic aphasia, when neurophysiological repair processes have largely been completed” (Page 2, Kiran and Thompson 2019; Smania et al. 2010). According to Kiran and Thompson (2019) “stroke affects 15 million people worldwide” (Page 1), and according to Berthier (2005), “Aphasia is present in 21–38% of acute stroke patients and is associated with high short- and long-term morbidity, mortality and expenditure” (Page 163). Clearly, aphasia has a negative impact on stroke survivors’ quality of life (Koleck et al. 2017; Flowers et al. 2016) and it increases their vulnerability to other diseases (e.g., cancer, Alzheimer’s and Parkinson’s disease) (Lam and Wodchis 2010). Brain networks supporting language are described in the literature as complex, spatially distributed, segregated interconnected, parallel, redundant, dynamic and bidirectional. However, following a stroke, these networks become disorganized, fragmented and partly silent (Fig. 2). In ischemic stroke, a blood vessel carrying blood and oxygen to the brain is occluded leading to a core of necrotic cell death. This area is surrounded by a perilesional area which remains more or less functional and is the potential target for brain neuromodulation.

Adapted from Del Gaizo et al. (2017) (with permission) (colour figure online)
This diagram illustrates the features evaluated by structural and functional connectivity measures. Both types of connectivity information are used to identify redundantly connected cortical areas as well as additional alternative white matter connections. a Considering a network of cortical structures. b If one of the cortical regions is lesioned after a stroke, its connections are also affected, shaded gray node or epicenter and gray lines. If all connections had similar importance regarding behavior (aphasia), CLSM would not distinguish between them. c However, CLSM can identify which connections are more important (c blue line). As such, CLSM is a subset of cortical lesion mapping. d CDLSM, in turn, provides information about the direct and indirect connections that may be crucial for behavior (d blue lines). According to the authors, CDLSM is unique because it can identify crucial corticocortical interactions that may occur indirectly even if their anatomic link is damaged.
Following a stroke, language recovery appears to be driven more by lesion location and load (Mohr 2017; Kiran and Thompson 2019) than by lesion size (Mattioli et al. 2014). Today language processing is described in terms of distributed networks (hodotopic view) rather than language centers (localizationist view) (Thiel and Zumbansen 2016). For information, Rolston and Chang (2017) have identified an average of 7.7 ± 5 crucial language epicenters. Recently, Klingbeil et al. (2019) observed that “aphasia results both from direct effects of focal damage to eloquent cortical areas as well as dysfunction of interconnected remote areas within the language network” (Page22). Del Gaizo et al. (2017) applied machine learning to ninety-two left hemisphere chronic stroke survivors. In this study, connectome lesion symptom mapping (CLSM) analysis is based on connection weight and provides information about the structure of the network, while connectome dynamics lesion symptom mapping (CDLSM) analysis is focused on extracting dynamic features from the individual connectome. Combined with lesion analysis, CLSM and CDLSM have allowed the authors to identify the crucial role played by the temporal parietal region. Their results indicate that “post-stroke deficits result from gray matter damage as well as secondary white matter loss. Conventional methods do not measure loss of connectivity beyond the stroke lesion” (Page 1, Del Gaizo et al. 2017) (Fig. 2). Yet, reentrant white-matter tracts link distant cortical areas and represent a key mechanism for integration of brain function as illustrated by Edelman and Gally (2013).
Marebwa et al. (2017) used graph theory methods and showed that even when cortical structures were relatively spared, “fragmentation to the brain neuronal network community structure,…were directly associated with more severe aphasia in the chronic period” (Page 9). Network modularity (reflecting global network organization) confers several advantages such as efficient computational processing and robustness following disruption (Bullmore and Sporns 2012). According to these authors, when the topology of the network is disrupted, the entire organization of the network architecture is affected. Indeed, the loss of connectivity hubs limits the transfer of information and predicts in part the capacity for recovery. These results highlight the importance of a combined action on network structure and dynamics in order to induce repair processes (see Sect. 5).
Yourganov et al. (2016) examined how the functioning of the language network is disrupted following a stroke in a large cohort of 90 persons with various types of aphasia or without aphasia. They observed that stroke can destroy gray matter areas serving as epicenters or crucial nodes in the language network. Stroke can also damage WMT and thus perturb communication between network nodes spared by the lesion (Fig. 3). Finally, stroke can affect communication between distant parts of the network (e.g., contralesional hemisphere) due to Wallerian degeneration. Thus, stroke can lead to language impairment because of cortical damage, because of disconnection by loss of WMT, or a combination of the two phenomena. These lesion-based and connectome-based approaches allowed the authors to differentiate cortical damage from disconnection and to show that both provide mostly similar but qualitatively different information (see Sect. 5).

Adapted from Yourganov et al. (2016) (with permission) (colour figure online)
Colors represent loadings on the features, reflecting their importance in all aphasia scores (The Aphasia Quotient includes: naming, speech repetition, auditory comprehension and fluency). In the left column, features are gray-matter cortical regions; in the right column, features are WMT. For example, the link between the angular gyrus and the posterior superior temporal gyrus is marked across all language functions examined. These parietal and posterior temporal areas identified by predictive multivariate analyses are considered crucial in the preservation of speech functions. This figure illustrates the relative contribution of WMT and gray-matter cortical regions in language. For example, the choice of the location for recording electrophysiological information and the location for applying electrical stimulation depend on observations from both gray and white matter. Information transfer during language occurs both at the cortical level via cortical TW and at the subcortical level via WMT. Thus, far from being an epiphenomenon, TW appears to contribute to information transfer (see Takahashi et al. (2011, 2015) for an example in the motor cortex).
Gross et al. (2013) used MEG to examine in 22 healthy subjects the role played by brain waves in comprehension of speech and showed that “segmentation and coding of speech rely on a nested hierarchy of entrained cortical oscillations” (Page 1). Regarding this study, Weaver (2013) commented that “speech consists of a hierarchy of components that each takes place on a different timescale. Speech cues such as intonation occur on a relatively long timescale, unfolding over hundreds of milliseconds. At the other end of the spectrum is the phoneme—the smallest unit of speech—which lasts only tens of milliseconds”. “Delta oscillations influenced the magnitude of theta oscillations, which in turn affected the amplitude of gamma oscillations” (Page 1, Weaver, 2013). Efficiently understanding speech necessitates simultaneous processing and coordinating of different components occurring on various timescales, with different delays, over the two hemispheres, and is characterized by high intersubject variability.
Mainy et al. (2008) explored the cortical dynamics of word recognition using word-like and nonword-like stimuli and observed a transient and local synchronization in the gamma frequency range (30–80 Hz) in brain areas involved in language functions. “These processes include (a) low and high level visual processing (letter and letter strings identification) and orthographic processing (of visual words and word-like forms), (b) phonological processing (conversion of graphemic representations into corresponding phonological representations and phono-articulatory programs), and (c) semantic processing (access to and activation of lexical representations)” (Page 10, Mainy et al. 2008). In terms of timing, the gamma band response takes around 200 ms for the midfusiform gyrus to differentiate between word-like and nonword-like stimuli, while semantic processing and phonological processing occur around 400 ms “with longer graphophonological conversion for pseudo-words than for words” (Page 1226, Mainy et al. 2008). Summarizing their observations, Sarubbo et al. (2015) write: “The medial part of dorsal stream (arcuate fasciculus) subserves phonological processing; its lateral part [indirect anterior portion of the superior longitudinal fascicle (SLF)] subserves speech planning. The ventral stream subserves language semantics and matches with the inferior fronto-occipital fascicle. Reading deficits match with the inferior longitudinal fascicle. Anomias match with the indirect posterior portion of the SLF. Frontal white matter underpins motor planning and execution” (Page 1, Sarubbo et al. 2015). Overall, these results confirm the distributed and dynamic organization of cortical sites involved in language processes (patterns of neuronal activation) and language representation (neuronal circuits).
Importantly, TWs have been observed during speech by Rapela (2018) using ECoG in neurosurgical patients producing consonant–vowel syllables (CVS). The author showed that TWs do not occur continuously, but tend to appear before the initiation of CVS and tend to disappear before their termination. During silent periods between the productions of CVS, TWs tend to reverse direction. Rapela observed a concentration of phase across trials at the specific frequency of speech production as well as amplitude modulation and phase amplitude coupling (PAC) (i.e., the relationship between the phase of oscillations in a specific frequency band to the amplitude of oscillations in another frequency band—Salimpour and Anderson, 2019). Again, this study showed that TWs induce an organization of PAC so that “spiking occurs at behaviorally relevant times” (Page 1, Rapela 2018) (in other words, TW across the left speech processing brain areas synchronized to the rhythm of speech production).
4 Neural field models
Neural field models treat brain tissue as an active medium. They are widely used to describe various patterns of brain activity including periodic brain waves (Senk et al. 2018; Pinto and Ermentrout 2001a, b; Atay and Hutt 2006; Venkov et al. 2007; Ermentrout et al. 2014; Gerstner et al. 2014; Meijer and Coombes 2014). For more information about neural field models, we refer the reader to the following references (Coombes 2005; Deco et al. 2008) and to the following websites:
https://pdfs.semanticscholar.org/555d/450b6bd80c3d2302c396a92902b4cb3d3dd2.pdf.
http://www.scholarpedia.org/article/Neural_fields.
The model proposed below (1.1)–(1.3) is a particular case of the existing models (see, e.g., Senk et al. 2018). In its simplest form, this model represents a single integro-differential equation for the electric potential in the cortex (Bessonov et al. 2018, 2019; Beuter et al. 2018). For example, the model suggested in Bessonov et al. 2018 differs by the diffusion term describing additional conduction mechanisms (see Buskila et al. 2019 for additional information). This simplest model describing the distribution u of the electric potential in the cortex is represented by the integro-differential equation:
where Wa and Wi are the activating and inhibiting signals, respectively:
(cf. Senk et al. 2018). The first expression describes activating signals coming to the location x from all other locations y. The connectivity function φa(x − y) shows how the strength of connection between these points depends on the distance between them. It is proportional to the density of connecting axons, and it decreases exponentially with distance. The response function Sa(u) characterizes signal production by neurons depending on the value of the electric potential. It is a sigmoid function with a transition from low to high values at some threshold level. Hence, neurons located at point y produce a signal depending on the value of u there. This signal arrives to the point x with time delay |x − y|/qa, where qa is the speed of its propagation along the axons. Finally, neuron response to the activation can occur with some additional time delay τa. The sum of all these signals coming to the point x is given by the integral in the expression Wa. Expression Wi for inhibiting signals has a similar structure. In the absence of activating signals, the value of the potential returns to its basic level (u = 0 in the model) provided by the last term in the right-hand side of Eq. (1.1) with the rate determined by a positive constant σ. Let us also note that more complete models can be considered, containing diffusion term, or a system of two equations instead of the single equation (Bessonov et al. 2018, 2019, Beuter et al. 2018). Numerical simulations are carried out in a bounded interval with periodic boundary conditions.
We consider in this work a single equation with an asymmetric kernel where the wave propagates only in one direction. There are experimental and modeling studies, suggesting that the kernel can be asymmetric (Pinotsis et al. 2013; Stephan et al. 2001). Propagation of periodic waves can also be observed in the case of symmetric kernels but for more complex models, including time delay or two equations.
The model we use describes various patterns of the potential distribution. Among them, periodic waves that can be approximated by the function cos(ξx + βt), where ξ is the spatial frequency, β is the time frequency, and c = − β/ξ is the wave speed (Fig. 4). As it is discussed above, these waves propagate between different parts of the cortex in order to synchronize their activity. Modulated waves (Fig. 5) contain oscillations with two different frequencies, and according to recent data by Turi et al. (2018), they transfer information about individual brain tasks in order to coordinate neural processing across the human cortex.

Reprinted from Bessonov et al. (2018) with permission
Snapshots of the periodic wave in numerical simulations with a neural field model and asymmetric connectivity function. It is a periodic in space solution moving with a constant speed. The function in the right figure can be obtained from the function in the left figure by a shift of half-period.

Reprinted from Bessonov et al. (2018), with permission
Snapshots of periodic waves with a periodically modulated amplitude (left) and with aperiodic amplitude modulation (right). The wave of amplitude modulation moves in the direction opposite to the direction of the periodic wave.
If a part of cortex tissue is damaged due to a stroke, then wave propagation is perturbed in the perilesional area. Then, the question is whether external stimulation can restore wave propagation, including its frequency, amplitude and speed of propagation. In order to study the effect of the stimulation, we introduce an additional term I(x; t) in the equation (see “Appendix”).
This stimulation is provided by an array of electrodes whose location depends precisely on the location of the perilesional area and on the location of epicenters to be modulated. The value of the stimulating signals at each electrode depends on the registered signals, providing the closed-loop stimulation technique.
An example of open-loop stimulation is shown in Fig. 6. The connectivity function vanishes in some part of the simulation domain, completely destroying wave propagation. An empirically chosen stimulation restores the wave propagation to a reasonably good level. The parameters of the stimulation are chosen in numerical experiments on the basis of the obtained results. Closed-loop stimulation and modern techniques of machine learning can be expected to improve the stimulation result and to produce them in real time in a patient-dependent manner.

Snapshots of the distribution of electric potential in numerical simulations with a neural field model with time delay in the response function and with a symmetric connectivity function. Left: a periodic TW in the normal tissue. Middle: if a part of the tissue is damaged (green interval), then, instead of TW, a time periodic solution is observed oscillating between the curves (a) and (b). Right: external stimulation allows the restoration of the periodic TW. The parameters values are presented in the “Appendix” (colour figure online)
Wave propagation is characterized by its speed, frequency and amplitude which is specific for each subject and for each particular brain function including speech. Assuming that the wave is initiated by epicenter A and propagates toward epicenter B, the distance between the epicenters and its integration in the brain network also affect the properties of the TW. If the wave propagates from A to B (see Rapela 2018 above), it can also propagate from B to A at, or just before, the end of the behavior.
5 Discussion
This prospect paper examines the opportunity of model-guided neuromodulation for the treatment of post-stroke aphasia using neuromodulation of TW. After providing some background about TW, examining the functional role and dynamics of TW, focusing on a network approach in post-stroke aphasia and presenting one neural field model, we argue that a new therapeutic strategy is possible. This neuromodulation approach is proposed to alleviate aphasia symptoms as a first indication; however, the approach is generic and could be extended to alleviate other post-stroke symptoms (e.g., motor) as well as other conditions.
We argue that a brain lesion (e.g., stroke) can interfere with the normal propagation of TW in the perilesional area, and we relate this abnormal propagation to changes in brain rhythmicity such as high gamma bursts activity.
Cerebrovascular stroke is a common cause of cognitive, language and motor impairments due to ischemic damage (Kielar et al., 2016). Among approximately 33 million of persons affected by stroke worldwide (Feigin et al., 2014), roughly 6–7 million have chronic aphasia, and there are no effective medical treatments to improve recovery significantly beyond speech-language therapies (Turkeltaub et al. 2016) during the chronic phase. Turkeltaub et al. (2011) examined language networks after a stroke and concluded that “the network of brain areas aphasic patients recruit for language functions is largely consistent across studies. Several recruitment mechanisms occur, including persistent function in spared nodes, compensatory recruitment of alternate nodes, and recruitment of areas that may hinder recovery” (Page 1726). However, exploiting these mechanisms to enhance or restore a function by electrical stimulation raises several questions as indicated by Sreekumar et al. (2017): Are there reliable neural patterns or networks corresponding to the targeted function? Yes, accumulating evidence of TW is now well documented. Can stimulation induce these patterns? Yes, studies using tACS, for example, show that this is the case. Finally, would inducing such patterns modulate the function in the appropriate way? Reconstructing TW during speech is expected to increase connectivity between epicenters and facilitate the migration of TW. However, before implementing model-guided closed-loop neuromodulation in the treatment of post-stroke aphasia, additional issues need to be considered:
5.1 First issue: Can brain networks be repaired by neuromodulation following stroke?
Several studies using invasive and noninvasive stimulation techniques for the treatment of post-stroke aphasia have shown conflicting results (Balossier et al. 2016). One of the main limits of these studies is that brain stimulation was applied with the same arbitrary parameters for every patient. The same findings were reported in a review of the literature on the effects of direct brain stimulation for modulating human memory and cognition (Sreekumar et al. 2017). Applying stimulation in such a way that it respects intrinsic states and brain dynamics of the patient may be one of the key factors for therapeutic success. For this reason, “mapping brain networks in individuals may provide new diagnostic tools or measures, and may even become important for developing new network-based interventions and therapies that capitalize on charting the network architecture of an individual’s brain” (Page 7, Sporns 2015). New approaches “may involve targeted manipulations of nodes or edges, e.g., with tools of brain stimulation” (Page 7, Sporns 2015; Luft et al. 2014; To et al. 2018), in order to restore network function. Of particular importance in this goal are new developments in computational neuroscience combining connectome data with realistic biophysical models that can reproduce and predict the dynamic activity of human brain networks. In a virtual study, Alstott et al. (2009) showed that a very small lesion (5%) could have network-wide bihemispheric effects when placed unilaterally within cortical midline hub regions. Certainly, the reverse question remains: Could the appropriate stimulation of a crucial and limited part of the network contribute to restoring network-wide functioning after a stroke? Furthermore, Murphy (2017) wrote: “Traveling oscillations may also be able to account for certain language disorders and aphasias, since in the literature there are often cases documenting damage to brain areas not typically seen as ‘language areas’ and yet which negatively impact language processing. It is possible that these damaged regions serve to ‘block’ the migration of traveling oscillations” (Page 27 Murphy 2017). Thus, the idea proposed in this paper is to facilitate the migration of TW between preidentified crucial nodes or epicenters by acting on wave parameters (i.e., speed, amplitude and frequency). Indeed, several studies have identified the potential of this type of neuromodulation.
Luft et al. (2014) suggested developing cortical stimulation protocols that target connectivity rather than individual areas. For LoPresti et al. (2015), the field of neuromodulation is a growing one and has great potential in improving functional recovery after stroke. Furthermore, noninvasive or minimally invasive interventions provide proven benefits to patients “by altering the post-stroke excitable brain architecture to remodel circuitry affected” by stroke (Page 4). LoPresti et al. (2015) conclude that “neuromodulation holds the potential to impact motor function, cognition, and pain syndromes after stroke, with the possibility of greatly impacting and improving quality of life and functional outcomes” (Page 4, LoPresti et al. 2015). For Azad et al. (2016), “Neurorestoration must remain a critical objective for stroke research; as cell-based therapies and neuromodulation are the two fields that have demonstrated the greatest promise in promoting recovery” (Page 9, Azad et al. 2016). Guggenmos et al. (2013) demonstrated that “recovery after brain injury can be facilitated by a neural prosthesis serving as a communication link between distant locations in the cerebral cortex” (Page 21177). For Assenza et al. (2017), neurophysiological markers of oscillatory dysfunctionalities are now available, so are the protocols interacting with these markers. Yet, there are still few brain stimulation clinical applications based on precise and selective neuromodulation of network activity.
In a recent review paper entitled “Flexible Redistribution in Cognitive Networks” Harwigsen (2018) examined “the potential for adaptive plasticity within and between large-scale neural networks” (Page 687) and specifically network compensation following a focal lesion. For the author, large networks including language networks present a resilience or robustness and manifest flexible compensation. In addition, inside large networks, specific nodes or hubs can increase their contribution to compensate for the disrupted function. Furthermore, “alternative pathways might be recruited to support task processing after perturbation” (Page 689, Harwigsen 2018) (see also Fig. 2 this paper). Finally, the author introduces a final form of flexible redistribution involving domain-specific networks. In the case of perturbation, recruitment of domain-general networks may also provide compensatory flexibility across networks (Page 689, Hartwigsen, 2018). However, the reverse does not hold (i.e., recruitment of domain-specific functions cannot compensate for disruptions of domain general functions). Language processes belong to domain-specific networks involving highly specialized brain regions, while domain-general networks are engaged in a wide variety of cognitive behaviors.
Concretely, Zhang et al. (2018) indicate that once reliable TW patterns are identified in a subject, a number of quantifiable properties can be used to provide key insights into instantaneous speed, direction, coherence, frequency, spatial extent and shape. “Measuring TW’s instantaneous properties may provide a new tool for neural interfacing for tracking a subject’s attention or cognitive state for timing stimulus presentation or modulating neuromodulation” (Page 11, Zhang et al. 2018). See also Ezzyat et al. (2017) using direct brain stimulation to modulate memory performance. Indeed, “the consideration of closed-loop approaches to repair cortical disconnection syndromes may open treatment options for a variety of conditions in which neural communication is disrupted, whether due to disease, injury, or idiopathic causes” (Page 21181, Guggenmos et al. 2013). Clearly today, the idea is to personalize and individualize neuromodulation to the specific needs of each patient.
5.2 Second issue: Where in the cortex, should neuromodulation be introduced in order to repair the damaged language network?
Yourganov et al. (2016) examined the spatial overlap of cortical networks using structural connectome-based and lesion-based analyses. Although current investigation methods and analyses have their intrinsic limitations, these two approaches help distinguish deficits caused by cortical necrosis from deficits caused by cortical disconnection (loss of WMT) and both the lesion-based and the connectome-based analyses agreed on the importance of the posterior part of the superior temporal gyrus (PSTG) and its link to the angular gyrus (AG) in predicting all language modality disturbances (i.e., naming, speech repetition, auditory comprehension and fluency) (Fig. 3). However, although both methods were predictive of post-stroke language impairments, they also captured different information. The connectome-based approach identified regions that were missed by the lesion-based approach. For example, they noted that frontal regions predictive of naming disturbances identified by lesion-based analyses were not identified by connectome-based analyses, and inversely, parietal regions predictive of auditory comprehension disturbances were identified by connectome-based analyses but not by lesion-based analyses. What does this mean?
First, the connectivity between PSTG and AG may be an interesting target for neuromodulation because of its importance and of the proximity to the temporoparietal junction. Indeed, the temporoparietal junction is identified as a multimodal area crucial for language tasks. One possibility is to stimulate in the vicinity of these two crucial nodes. Second, some frontal regions predictive of naming disturbances are affected by ischemic damage while having intact WMT, thus leading to another possible target. Certainly modulating residual connectivity of cortical tissue can also be considered. Third, some spared nodes are likely to be missed in the lesion-based analysis, according to Yourganov et al. (2016) and these spared epicenters are a possible target to reshape the network via secondary nodes (Fig. 1).
Of particular interest is the cooperation between cortical TW and WMT in the production of smooth language. Under healthy conditions, information carried by TW migrates at the surface of the cortex to crucial epicenters/hubs which show increased connectivity. In addition, information carried through deeper WMTs also converge toward epicenters. Both sources of information (superficial and deep) must be carefully synchronized for behavior to unfold smoothly. For example, anatomofunctional relations while naming an object presented visually requires (1) visual processing (occipital cortex and fusiform gyrus), then (2) semantic processing (temporal pole), and finally (3) phonological and articulatory processing (parietal and prefrontal regions) (Grappe et al. 2018). We surmise that TWs arrive “just in time” for activation of task-relevant brain regions and stop “just in time” at the end of the behavior. Language smoothness depends on coordination of multiple types of information. Mandonnet et al. (2018) examined diffusion weighted magnetic resonance imaging and tractography to reconstruct very precisely the trajectories of WMT. These fibers are spread out and the possibility exists that even when this pathway is damaged by stroke, a limited percentage of spared fibers might still remain functional and could be solicited by neuromodulation (Shi and Toga 2017). Furthermore, anatomically intact but functionally silent fibers represent an additional potential target for the neuromodulation proposed here. The solution might be to record over a specific perilesional area and to stimulate over corresponding crucial epicenters, the idea being to activate centers exactly as the brain would do it naturally, i.e., with precision and selectivity. Interestingly, López-Barroso and de Diego-Balaguer (2017) examined “the division of labor of the ventral and dorsal streams” (Page 1) for language and in particular regions showing greater likelihood to be redundantly connected through the ventral and dorsal pathways including posterior superior temporal gyrus and inferior frontal gyrus (Figure 1 in Lopez-Barroso 2017). These redundancies allow for compensation mechanisms after brain damage soliciting alternative pathways which can be exploited by closed-loop, model-guided neuromodulation (Fig. 1).
5.3 Third issue: How will electrical stimulation be integrated into cortical TW?
Model-based cortical neuromodulation offers the possibility to extrapolate conduction speed and strength in the perilesional area. In addition, it offers an opportunity to calculate in quasi-real time the stimulation signal to be injected. Finally, it facilitates the computation of the stimulation parameters in a closed-loop design (Beuter et al. 2014, 2018; Bessonov et al. 2018, 2019). Regarding electrical stimulation, Voigt et al. (2018) show that activity elicited by low-current intracortical micro-stimulation “modulates induced cortical responses to a sensory stimulus in the primary auditory cortex” (Page 1). This experiment was not performed in human subjects but in anesthetized guinea pigs of both sexes. Intracortical Microstimulation (ICMS) is commonly used to activate a specific subset of cortical neurons, without taking into account the ongoing activity at the time of stimulation. Here, we found that a low-current ICMS pulse modulated the way the auditory cortex processed a peripheral stimulus, by supra-additively combining the response to the ICMS with the cortical processing of the peripheral stimulus. This artificial modulation mimicked natural modulations of response magnitude such as attention or expectation. In contrast to what was implied in earlier studies, this shows that the response to electrical stimulation is not substituting ongoing cortical activity but is integrated into the natural processes” (Page 7774, Voigt et al. 2018). These results confirm those of Jackson et al. (2006) showing with a different technique that induced functional reorganization in vivo using physiologically derived stimulus trains produce artificial long-term motor cortex plasticity. For an example of TW control in the cortex via electrical stimulation, see Richardson et al. (2005).
5.4 Fourth issue: Given the encouraging results of invasive and noninvasive cortical stimulation, why are we proposing invasive cortical neuromodulation?
A large number of invasive and noninvasive stimulation studies based on electrical epidural stimulation, transcranial magnetic stimulation and various types of transcranial electrical stimulation have produced promising results (Töpper et al. 1998; Kim et al. 2008; Flöel et al. 2008; Naeser et al. 2005; Baker et al. 2010; Cherney et al. 2012; Balossier et al. 2012; Cherney 2016; Duncan and Small 2016; Hamilton et al. 2011; Hamilton 2016; Turkeltaub et al. 2016; Wortman-Jutt and Edwards 2017; Harwigsen and Saur 2017). These approaches have limitations which have been discussed (Beuter et al. 2018). In brief, “Effect sizes in the order of 8–30% of functional improvement have been reported, but a publication bias toward presenting “promising” but not negative results is likely” (Page 370, Hummel et al. 2008). These approaches function in open loop, and the stimulation parameters are not guided by a model or by smart algorithms. However, they produce encouraging results when combined with rehabilitation programs.
What we propose is qualitatively different. Our aim is to reconstruct cortical TW migrating at the surface of the cortex and to integrate this information in the computation of the stimulation to be delivered. The future work will decide whether this reconstruction increases the residual connectivity and lead to a form of network repair (i.e., neuroregeneration) or if reconstruction increases the neuroplasticity (i.e., synaptic long-term potentiation in Balossier et al. 2016). Today, adaptive, responsive and closed-loop stimulation systems are the focus of intense discussion (Zanos 2018; Hazan and Ziv 2017; Hoang et al. 2017; Levi et al. 2018; Parastarfeizabadi and Kouzani 2017; Sun and Morrell 2014; Wright et al. 2016). Closed-loop stimulation systems record specific signals, and adjustments of stimulation settings are performed in real time based on the ongoing neurophysiological variations. These adjustments lead to a specific pattern of stimulation for each patient and period of time. Closed-loop stimulation is used in deep brain stimulation for Parkinson’s disease (Herron et al. 2017; Kern et al. 2016; Little et al. 2013; Meidahl et al. 2017; Priori et al. 2013; Rosin et al. 2011; Swann et al. 2018), for pain (Shirvalkar et al. 2018) and for epilepsy (Edwards et al. 2017). There are also promising results with episodic memory (Ezzyat et al. 2018; Hanslmayr and Roux 2017). All these studies have shown that not only closed-loop stimulation methods reached the same efficacy as open-loop stimulation, but it also reduced side effects and battery consumption (Parastarfeizabadi and Kouzani 2017). A new generation of closed-loop cortical stimulation systems is about to arrive on the market (Gkogkidis et al. 2017; Kohler et al. 2017).
6 Conclusion
Stroke is the leading cause of acquired long-term disability and globally produces immense health, social and economic burdens. Following a large cortical stroke, there is a network-wide reduction in neural activity (Fink 2017) and an “increasing number of stroke survivors are left with persistent neurological deficits” including motor, cognitive or neuropsychological (Page 1002, Fink 2017). However, the neuroplasticity of the language networks is still present even at the chronic stage. Thus, more selective and precise neuromodulation approaches to rehabilitation are urgently needed (Fink 2017). The proposed strategy is based on the integration of different bodies of research (i.e., brain stimulation, neural field modeling, experimental and clinical observation of TW, neuroimagery and electrophysiology). We conclude that closed-loop, model-guided, cortical neuromodulation acting precisely and selectively on crucial epicenters to modulate cortical TW should be explored. This neuromodulation technique is likely to influence brain activity not only in the vicinity of the stimulation site but also in remote areas involved in language (Fink 2017). The technical aspects supporting this precision neuromodulation (e.g., sensors, acquisition system, processing unit, output device, power source, wireless transmission and implementation of closed-loop control) still need to be addressed.
References
Alamia A, VanRullen R (2019) Alpha oscillations and traveling waves: signatures of predictive coding? PLoS Biol 17(10):e3000487. https://doi.org/10.1371/journal.pbio.3000487
Alekseichuk I, Falchier AY, Linn G, Xu T, Milham MP, Schroeder CE, Opitz A (2016) Electric field dynamics in the brain during multi-electrode transcranial electric stimulation. bioRxiv preprint https://doi.org/10.1101/340224
Alexander DM, Jurica P, Trengove C, Nikolaev AR, Gepshtein S, Zvyagintsev M et al (2013) Traveling waves and trial averaging: the nature of single-trial and averaged brain responses in large-scale cortical signals. NeuroImage 73:95–112. https://doi.org/10.1016/j.neuroimage.2013.01.016
Alexander DM, Nikolaev AR, Jurica P, Zvyagintsev M, Mathiak K, van Leeuwen C (2016) Global neuromagnetic cortical fields have non-zero velocity. PLoS ONE 11(3):e0148413. https://doi.org/10.1371/journal.pone.0148413
Alstott J, Breakspear M, Hagmann P, Cammoun L, Sporns O (2009) Modeling the impact of lesions in the human brain. PLoS Comput Biol 5(6):e1000408. https://doi.org/10.1371/journal.pcbi.1000408
Amari S (1977) Dynamics of pattern formation in lateral-inhibition type neural fields. Biol Cybern 27:77–87
Aru J, Aru J, Priesemann V, Wibral M, Lana L, Pipa G et al (2015) Untangling cross-frequency coupling in neuroscience. Curr Opin Neurobiol 31:51–61. https://doi.org/10.1016/j.conb.2014.08.002
Assenza G, Capone F, di Biase L, Ferreri F, Florio L, Guerra A, Marano M, Paolucci M, Ranieri F, Salomone G, Tombini M, Thut G, Di Lazzaro V (2017) Oscillatory activities in neurological disorders of elderly: biomarkers to target for neuromodulation. Front Aging Neurosci 9:189. https://doi.org/10.3389/fnagi.2017.00189
Atasoy S, Donnelly I, Pearson J (2016) Human brain networks function in connectome specific harmonic waves. Nat Commun 7:10340. https://doi.org/10.1038/ncomms10340
Atay FM, Hutt A (2006) Neural fields with distributed transmission speeds and long-range feedback delays. SIAM J Appl Dyn Syst 5(4):670–698
Azad TD, Veeravagu A, Steinberg GK (2016) Neurorestoration after stroke. Neurosurg Focus 40(5):E2. https://doi.org/10.3171/2016.2.FOCUS15637
Bahramisharif A, van Gerven MAJ, Aarnoutse EJ, Mercier MR, Schwartz TH et al (2013) Propagating neocortical gamma bursts are coordinated by traveling alpha waves. J Neurosci 33:18849–18854
Baker JM, Rorden C, Fridriksson J (2010) Using transcranial direct-current stimulation to treat stroke patients with aphasia. Stroke 41:1229–1236. https://doi.org/10.1161/strokeaha.109.576785
Balossier A, Etard O, Descat C, Vivien D, Emery E (2012) Epidural electrical stimulation to improve chronic poststroke aphasia: a 5-year follow-up. Brain Stimul 5(3):364–368. https://doi.org/10.1016/j.brs.2011.04.003
Balossier A, Etard O, Descat C, Vivien D, Emery E (2016) Epidural cortical stimulation as a treatment for poststroke aphasia: a systematic review of the literature and underlying neurophysiological mechanisms. Neurorehabil Neural Repair 30(2):120–130. https://doi.org/10.1177/1545968315606989
Berthier ML (2005) Poststroke aphasia. Drugs Aging 22:163–182. https://doi.org/10.2165/00002512-200522020-00006
Bessonov N, Beuter A, Trofimchuk S, Volpert V (2018) Cortical waves and post-stroke brain stimulation. Math Methods Appl Sci. https://doi.org/10.1002/mma.5620
Bessonov N, Beuter A, Trofimchuk S, Volpert V (2019) Estimate of the travelling wave speed for an integro-differential equation. Appl Math Lett 88:103–110. https://doi.org/10.1016/j.aml.2018.07.037
Beuter A, Lefaucheur JP, Modolo J (2014) Closed-loop cortical neuromodulation in Parkinson’s disease: an alternative to deep brain stimulation? Clin Neurophysiol 125:874–885
Beuter A, Balossier A, Trofimchuk S, Volpert V (2018) Modelling of post-stroke stimulation of the cortical tissue. Math Biosci. https://doi.org/10.1016/j.mbs.2018.08.014
Brasic JR (1998) Hallucinations. Percept Motor Skills 86:851–877
Breakspear M (2017) Dynamic models of large-scale brain activity. Nat Neurosci 20(3):340–352. https://doi.org/10.1038/nn.4497
Bullmore E, Sporns O (2012) The economy of brain network organization. Nat Rev Neurosci 13:336–349. https://doi.org/10.1038/nrn3214
Burkitt GR, Silberstein RB, Cadusch PJ, Wood AW (2000) Steady-state visual evoked potentials and travelling waves. Clin Neurophysiol 111(2):246–258. https://doi.org/10.1016/S1388-2457(99)00194-7
Buskila Y, Bellot-Saez A, Morley JW (2019) Generating brain waves, the power of astrocytes. Front Neurosci 13:1125. https://doi.org/10.3389/fnins.2019.01125
Canolty RT, Knight RT (2010) The functional role of cross-frequency coupling. Trends Cogn Sci 14(11):506–515. https://doi.org/10.1016/j.tics.2010.09.001
Cherney LR (2016) Epidural cortical stimulation as adjunctive treatment for nonfluent aphasia: phase 1 clinical trial follow-up findings. Neurorehab Neural Repair 30(2):131–142. https://doi.org/10.1177/1545968315622574
Cherney LR, Harvey RL, Babbitt EM, Hurwitz R, Kaye RC, Lee JB et al (2012) Epidural cortical stimulation and aphasia therapy. Aphasiology 26(9):1192–1217. https://doi.org/10.1080/02687038.2011.603719
Chrostowski M, Yang L, Wilson HR, Bruce IC, Becker S (2011) Can homeostatic plasticity in deafferented primary auditory cortex lead to travelling waves of excitation? J Comput Neurosci 30(2):279–299
Coombes S (2005) Waves, bumps, and patterns in neural field theories. Biol Cybern 93(2):91–108
Coombes S, beim Graben P, Potthast R, Wright J (Eds) (2014) Neural fields theory and applications. Springer, Berlin. https://doi.org/10.1007/978-3-642-54593-1
Dahlem MA, Chronicle EP (2004) A computational perspective on migraine aura. Prog Neurobiol 74:351–361
Deco G, Jirsa VK, Robinson PA, Breakspear M, Friston K (2008) The dynamic brain: from spiking neurons to neural masses and cortical fields. PLoS Comput Biol 4(8):e1000092. https://doi.org/10.1371/journal.pcbi.1000092
Del Gaizo J, Fridriksson J, Yourganov G, Hillis AE, Hickok G, Misic B et al (2017) Mapping language networks using the structural and dynamic brain connectomes. eNeuro. https://doi.org/10.1523/ENEURO.0204-17.2017
Duncan ES, Small SL (2016) Increased modularity of resting state networks supports improved narrative production in aphasia recovery. Brain Connect 6(7):524–529. https://doi.org/10.1089/brain.2016.0437
Edelman GM, Gally JA (2013) Reentry: a key mechanism for integration of brain function. Front Integr Neurosci 7:63. https://doi.org/10.3389/fnint.2013.00063
Edwards CA, Kouzani A, Lee KH, Ross EK (2017) Neurostimulation devices for the treatment of neurologic disorders. Mayo Clin Proc 92(9):1427–1444. https://doi.org/10.1016/j.mayocp.2017.05.005
Ermentrout GB, Kleinfeld D (2001) Traveling electrical waves in cortex: insights from phase dynamics and speculation on a computational role. Neuron 29:33–44
Ermentrout B, McLeod B (1993) Existence and uniqueness of travelling waves for a neural network. Proc R Soc Edinb Sect A 123(3):461–478
Ermentrout GB, Folias SE, Kilpatrick ZP (2014) Spatiotemporal pattern formation in neural fields with linear adaptation. In: Coombes S et al (eds) Neural fields. Springer, Berlin, pp 119–151
Ezzyat Y, Kragel JE, Burke JF, Levy DF, Lyalenko A, Wanda P (2017) Direct brain stimulation modulates encoding states and memory performance in humans. Curr Biol 27:1–8. https://doi.org/10.1016/j.cub.2017.03.028
Ezzyat Y, Wanda PA, Levy DF, Kadel A, Aka A, Pedisich I et al (2018) Closed-loop stimulation of temporal cortex rescues functional networks and improves memory. Nat Commun 9:365. https://doi.org/10.1038/s41467-017-02753-0
Feigin VL et al (2014) Global and regional burden of stroke during 1990–2010: findings from the Global Burden of Disease Study 2010. Lancet 383(9913):245–254. https://doi.org/10.1016/S0140-6736(13)61953-4
Fellinger R, Gruber W, Zauner A, Freunberger W, Klimesch W (2011) Evoked traveling alpha waves predict visual-semantic categorization-speed. NeuroImage 59:3379–3388. https://doi.org/10.1016/j.neuroimage.2011.11.010
Fink GR (2017) New hope for ameliorating stroke-induced deficits? Brain 139:1002–1004. https://doi.org/10.1093/brain/aww034
Flöel A, Rösser N, Michka O, Knecht S, Breitenstein C (2008) Noninvasive brain stimulation improves language learning. J Cogn Neurosci 20(8):1415–1422
Florin E, Baillet S (2015) The brain’s resting-state activity is shaped by synchronized cross-frequency coupling of neural oscillations. Neuroimage 111:26–35. https://doi.org/10.1016/j.neuroimage.2015.01.054
Flowers HL, Skoretz SA, Silver FL, Rochon E, Fang J, Flamand-Roze C et al (2016) Poststroke aphasia frequency, recovery, and outcomes: a systematic review and meta-analysis. Arch Phys Med Rehabil 97(2188):201.e8. https://doi.org/10.1016/j.apmr.2016.03.006
Gerstner W, Kistler WM, Naud R, Paninski L (2014) Neuronal dynamics. From single neurons to networks and models of cognition. Cambridge University Press, Cambridge
Gkogkidis C, Xi Wang A, Schubert T, Gierthmühlen M, Kohler F, Schulze-Bonhage A et al (2017) Closed-loop interaction with the cerebral cortex using a novel micro-ECoG-based implant: the impact of beta vs. gamma stimulation frequencies on cortico-cortical spectral responses. Brain Comput Interfaces 4(4):214–224. https://doi.org/10.1080/2326263X.2017.1381829
Grappe A, Sarma SV, Sacré P, Gonzàlez-Martınez J, Liégeois-Chauvel C, Alario FX (2018) An intracerebral exploration of functional connectivity during word production. J Comput Neurosci. https://doi.org/10.1007/s10827-018-0699-3
Gross J, Hoogenboom N, Thut G, Schyns P, Panzeri S, Belin P et al (2013) Speech rhythms and multiplexed oscillatory sensory coding in the human brain. PLoS Biol 11(12):e1001752. https://doi.org/10.1371/journal.pbio.1001752
Guggenmos DJ, Azin M, Barbay S, Mahnken JD, Dunham C, Mohseni P et al (2013) Restoration of function after brain damage using a neural prosthesis. PNAS 110(52):21177–21182. https://doi.org/10.1073/pnas.1316885110
Hamilton RH (2016) Neuroplasticity in the language system: reorganization in post-stroke aphasia and in neuromodulation interventions. Restorative Neurol Neurosci 34:467–471. https://doi.org/10.3233/RNN-169002
Hamilton RH, Chrysikou EG, Coslett B (2011) Mechanisms of aphasia recovery after stroke and the role of noninvasive brain stimulation. Brain Lang 118(1–2):40–50. https://doi.org/10.1016/j.bandl.2011.02.005
Hanslmayr S, Roux F (2017) Human memory: brain-state- dependent effects of stimulation. Curr Biol 27:R378–R397
Harwigsen G (2018) Flexible redistribution in cognitive networks. Trends Cogn Sci. https://doi.org/10.1016/j.tics.2018.05.008
Harwigsen G, Saur D (2017) Neuroimaging of stroke recovery from aphasia—insights into plasticity of the human language network. NeuroImage. https://doi.org/10.1016/j.neuroimage.2017.11.056
Hazan H, Ziv NE (2017) Closed Loop Experiment Manager (CLEM)—an open and inexpensive solution for multichannel electrophysiological recordings and closed loop experiments. Front Neurosci 11:579. https://doi.org/10.3389/fnins.2017.00579
Heitmann S, Boonstra T, Breakspear M (2013) A dendritic mechanism for decoding traveling waves: principles and applications to motor cortex. PLoS Comput Biol 9(10):e1003260. https://doi.org/10.1371/journal.pcbi.1003260
Herron JA, Thompson MC, Brown T, Chizeck HJ, Ojemann JG, Ko AL (2017) Cortical brain computer interface for closed-loop deep brain stimulation. IEEE Trans Neural Syst Rehabil Eng. https://doi.org/10.1109/TNSRE.2017.2705661
Hoang KB, Cassar IR, Grill WM, Turner DA (2017) Biomarkers and stimulation algorithms for adaptive brain stimulation. Front Neurosci 11:564. https://doi.org/10.3389/fnins.2017.00564
Hummel FC, Celnik P, Pascual-Leone A, Fregni F, Byblow WD, Buetefisch CM (2008) Controversy: noninvasive and invasive cortical stimulation show efficacy in treating stroke patients. Brain Stimul 1(4):370–382. https://doi.org/10.1016/j.brs.2008.09.003
Hyafil A (2015) Misidentifications of specific forms of cross-frequency coupling: three warnings. Improved narrative production in aphasia recovery. Brain Connect. https://doi.org/10.1089/brain.2016.0437
Jackson A, Mavoori J, Fetz EE (2006) Long-term motor cortex plasticity induced by an electronic neural implant. Nature 444:2. https://doi.org/10.1038/nature05226
Jirsa VK, Haken H (1996) Field theory of electromagnetic brain activity. Phys Rev Lett 77:960–963
Kern K, Naros G, Braun C, Weiss D, Gharabaghi A (2016) Detecting a cortical fingerprint of Parkinson’s disease for closed-loop neuromodulation. Front Neurosci 10:110. https://doi.org/10.3389/fnins.2016.00110
Khambhati AN, Kahn AE, Costantini J, Ezzyat Y, Solomon EA, Gross RE, Bassett DS (2019) Functional control of electrophysiological network architecture using direct neurostimulation in humans. Netw Neurosci 3(3):848–877. https://doi.org/10.1162/netn_a_00089
Kielar A, Deschamps T, Chu RKO, Jokel R, Khatamian YB, Chen JJ, Meltzer JA (2016) Identifying dysfunctional cortex: dissociable effects of stroke and aging on resting state dynamics in MEG and fMRI. Front Aging Neurosci 8:40. https://doi.org/10.3389/fnagi.2016.00040
Kim HI, Shin YI, Moon SK, Chung GH, Lee MC, Kim HG (2008) Unipolar and continuous cortical stimulation to enhance motor and language deficit in patients with chronic stroke: report of 2 cases. Surg Neurol 69:77–80
Kiran S (2012) What Is the Nature of Poststroke Language Recovery and Reorganization? Int Sch Res Netw ISRN Neurol 2012:786872. https://doi.org/10.5402/2012/786872
Kiran S, Thompson CK (2019) Neuroplasticity of language networks in aphasia: advances, updates, and future challenges. Front Neurol 10:295. https://doi.org/10.3389/fneur.2019.00295
Klimesch W (2018) The frequency architecture of brain and brain body oscillations: an analysis. Eur J Neurosci 48:2431–2453. https://doi.org/10.1111/ejn.14192
Klimesch W, Hanslmayr S, Sauseng P, Gruber WR, Doppelmayr M (2007) P1 and traveling alpha waves: evidence for evoked oscillations. J Neurophysiol 97:1311–1318. https://doi.org/10.1152/jn.00876.2006
Klingbeil J, Wawrzyniak M, Stockert A, Saur D (2019) Resting-state functional connectivity: an emerging method for the study of language networks in post-stroke aphasia. Brain Cogn 131:22–33. https://doi.org/10.1016/j.bandc.2017.08.005
Kohler F, Gkogkidi CA, Bentler C, Wang X, Gierthmuehlen M, Fischer J et al (2017) Closed-loop interaction with the cerebral cortex: a review of wireless implant technology. Brain Comput Interfaces 4(3):146–154. https://doi.org/10.1080/2326263X.2017.1338011
Koleck M, Gana K, Lucot C, Darrigrand B, Mazaux JM, Glize B (2017) Quality of life in aphasic patients 1 year after a first stroke. Qual Life Res 26:45–54. https://doi.org/10.1007/s11136-016-1361-z
Kopell N, Gritton HJ, Whittington MA, Kramer MA (2014) Beyond the connectome: the dynome. Neuron 83(6):1319–1328. https://doi.org/10.1016/j.neuron.2014.08.016
Lam JMC, Wodchis WP (2010) The relationship of 60 disease diagnoses and 15 conditions to preference-based health-related quality of life in Ontario hospital-based long-term care residents. Med Care 48:380–387. https://doi.org/10.1097/MLR.0b013e3181ca2647
Levi T, Bonifazi P, Massobrio P, Chiappalone M (2018) Editorial: closed-loop systems for next-generation neuroprostheses. Front Neurosci 12:26. https://doi.org/10.3389/fnins.2018.00026
Little S, Pogosyan A, Neal S, Zavala B, Zrinzo L, Hariz M et al (2013) Adaptive deep brain stimulation in advanced Parkinson’s disease. Ann Neurol 74:449–457. https://doi.org/10.1002/ana.23951
Lopes da Silva FH, Blanes W, Kalitzin SN, Parra J, Suczynski P, Velis DN (2003) Dynamical diseases of brain systems: different routes to epileptic seizures. IEEE Trans Biomed Eng 50(5):540–548
López-Barroso D, de Diego-Balaguer R (2017) Language learning variability within the dorsal and ventral streams as a cue for compensatory mechanisms in aphasia recovery. Front Hum Neurosci 11:476. https://doi.org/10.3389/fnhum.2017.00476
LoPresti MA, Camacho E, Appelboom G, Connolly ES Jr (2015) The promising role of neuromodulation in improving ischemic stroke recovery. J Neurol Neurosurg 1(2):112. https://doi.org/10.19104/jnn.2015.112
Luft CDB, Pereda E, Baniss MJ, Bhattacharya J (2014) Best of both worlds: promise of combining brain stimulation and brain connectome. Front Syst Neurosci 8:132. https://doi.org/10.3389/fnsys.2014.00132
Mainy N, Jung J, Baciu M, Kahane P, Schoendorff B, Minotti L et al (2008) Cortical dynamics of word recognition. Hum Brain Mapp 29:1215–1230
Mandonnet E, Sarubbo S, Petit L (2018) The nomenclature of human white matter association pathways: proposal for a systematic taxonomic anatomical classification. Front Neuroanat 12:94. https://doi.org/10.3389/fnana.2018.00094
Marebwa BK, Fridriksson J, Yourganov G, Feenaughty L, Rorden C, Bonilha L (2017) Chronic post-stroke aphasia severity is determined by fragmentation of residual white matter networks. Sci Rep 7:8188. https://doi.org/10.1038/s41598-017-07607-9
Mattioli F, Ambrosi C, Mascaro L, Scarpazza C, Pasquali P, Frugoni M, Magoni M, Biagi L, Gasparotti R (2014) Early aphasia rehabilitation is associated with functional reactivation of the left inferior frontal gyrus: a pilot study. Stroke 45:545–552. https://doi.org/10.1161/STROKEAHA.113.003192
Meidahl AC, Tinkhauser G, Herz DM, Cagnan H, Debarros J, Brown P (2017) Adaptive deep brain stimulation for movement disorders: the long road to clinical therapy. Mov Disord 32(6):2017. https://doi.org/10.1002/mds.27022
Meijer HGE, Coombes S (2014) Travelling waves in a neural field model with refractoriness. J Math Biol 68:1249–1268
Mohr B (2017) Neuroplasticity and functional recovery after intensive language therapy in chronic post stroke aphasia: which factors are relevant? Front Hum Neurosci. https://doi.org/10.3389/fnhum.2017.00332
Muller L, Chavane F, Reynolds J, Sejnowski TJ (2018) Cortical travelling waves: mechanisms and computational principles. Nat Rev Neurosci 19(5):255–268. https://doi.org/10.1038/nrn.2018.20
Murphy E (2017) Implications of travelling weakly coupled oscillators for the cortical language circuit. In: UCLWPL 2017
Naeser MA, Martin PI, Nicholas M, Baker EH, Seekins H, Kobayashi M et al (2005) Improved picture naming in chronic aphasia after TMS to part of right Broca’s area: an open-protocol study. Brain Lang 93(1):95–105. https://doi.org/10.1016/j.bandl.2004.08.004
Nishida M, Korzeniewska A, Crone NE, Toyoda G, Nakai Y, Ofen N et al (2017) Brain network dynamics in the human articulatory loop. Clin Neurophysiol 128(8):1473–1487. https://doi.org/10.1016/j.clinph.2017.05.002
Parastarfeizabadi M, Kouzani AZ (2017) Advances in closed-loop deep brain stimulation devices. J NeuroEng Rehabil 14:79. https://doi.org/10.1186/s12984-017-0295-1
Patten TM, Rennie CJ, Robinson PA, Gong P (2012) Human cortical traveling waves: dynamical properties and correlations with responses. PLoS ONE 7(6):e38392. https://doi.org/10.1371/journal.pone.0038392
Pinotsis DA, Hansen E, Friston KJ, Jirsa VK (2013) Anatomical connectivity and the resting state activity of large cortical networks. Neuroimage 65:127–138
Pinto DJ, Ermentrout GB (2001a) Spatially structured activity in synaptically coupled neuronal networks: I. Travelling fronts and pulses. SIAM J Appl Math 62(1):206–225
Pinto DJ, Ermentrout GB (2001b) Spatially structured activity in synaptically coupled neuronal networks: II. Lateral inhibition and standing pulses. SIAM J Appl Math 62(1):226–243
Priori A, Foffani G, Rossi L, Marceglia S (2013) Adaptive deep brain stimulation (aDBS) controlled by local field potential oscillations. Exp Neurol 245:77–86. https://doi.org/10.1016/j.expneurol.2012.09.013
Rabiller G, He JW, Nishijima Y, Wong A, Liu J (2015) Perturbation of brain oscillations after ischemic stroke: a potential biomarker for post-stroke function and therapy. Int J Mol Sci 16:25605–25640. https://doi.org/10.3390/ijms161025605
Rapela J (2018) Traveling waves appear and disappear in unison with produced speech. arXiv:1806.09559v1 [q-bio.NC]
Richardson KA, Schiff SJ, Gluckman BJ (2005) Control of traveling waves in the mammalian cortex. Phys Rev Lett 94(2):028103. https://doi.org/10.1103/PhysRevLett.94.028103
Roberts JA, Gollo LL, Abeysuriya RG, Roberts G, Mitchell PB, Woolrich MW, Breakspear M (2019) Metastable brain waves. Nat Commun 10:1056. https://doi.org/10.1038/s41467-019-08999-0
Robinson PA, Rennie CJ, Wright JJ (1997) Propagation and stability of waves of electrical activity in the cerebral cortex. Phys Rev E 56:826–840
Rolston JD, Chang EF (2017) Critical language areas show increased functional connectivity in human cortex. Cereb Cortex. https://doi.org/10.1093/cercor/bhx271
Rosin B, Slovik M, Mitelman R, Rivlin-Etzion M, Haber SN, Israel Z et al (2011) Closed-Loop deep brain stimulation Is superior in ameliorating parkinsonism. Neuron 72:370–384. https://doi.org/10.1016/j.neuron.2011.08.023
Rubino D, Robbins KA, Hatsopoulos NG (2006) Propagating waves mediate information transfer in the motor cortex. Nat Neurosci 9(12):1549–1557
Salimpour Y, Anderson WS (2019) Cross-frequency coupling based neuromodulation for treating neurological disorders. Front Neurosci 13:125. https://doi.org/10.3389/fnins.2019.00125
Sarubbo S, De Benedictis A, Merler S, Mandonnet E, Balbi S, Granieri E, Duffau H (2015) Towards a functional atlas of human white matter. Hum Brain Mapp 36:3117–3136. https://doi.org/10.1002/hbm.22832
Sato TK, Nauhaus I, Carandini M (2012) Traveling waves in visual cortex. Neuron 75(2):218–229. https://doi.org/10.1016/j.neuron.2012.06.029
Senk J, Korvasová K, Schuecker J, Hagen E, Tetzlaff T, Diesmann M et al (2018) Conditions for traveling waves in spiking neural networks. arXiv:1801.06046v1 [q-bio.NC]
Shi Y, Toga AW (2017) Connectome imaging for mapping human brain pathways. Mol Psychiatry 22:1230–1240. https://doi.org/10.1038/mp.2017.92
Shirvalkar P, Veuthey TL, Dawes HE, Chang EF (2018) Closed-loop deep brain stimulation for refractory chronic pain. Front Comput Neurosci 12:18. https://doi.org/10.3389/fncom.2018.00018
Smania N, Gandolfi M, Aglioti MA, Girardi P, Fiaschi A, Girardi F (2010) How long is the recovery of global aphasia? Twenty-five years of follow-up in a patient with left hemisphere stroke. Neurorehab Neural Repair 24(9):871–875. https://doi.org/10.1177/1545968310368962
Sporns O (2015) Cerebral cartography and connectomics. Philos Trans R Soc B 370:20140173. https://doi.org/10.1098/rstb.2014.0173
Sreekumar V, Wittig JH Jr, Sheehan TC, Zaghloul KA (2017) Principled approaches to direct brain stimulation for cognitive enhancement. Front Neurosci 11:650. https://doi.org/10.3389/fnins.2017.00650
Stephan KE, Kamper L, Bozkurt A, Burns GAP, Young MP, Kötter R (2001) Advanced database methodology for the collation of connectivity data on the macaque brain (CoCoMac). Philos Trans R Soc London Ser B Biol Sci 356:1159–1186
Sun FT, Morrell MJ (2014) Closed-loop neurostimulation: the clinical experience. Neurotherapeutics 11:553–563. https://doi.org/10.1007/s13311-014-0280-3
Swann NC, de Hemptinne C, Thompson MC, Miocinovic S, Miller AM, Gilron R et al (2018) Adaptive deep brain stimulation for Parkinson’s disease using motor cortex sensing. J Neural Eng 15:046006. https://doi.org/10.1088/1741-2552/aabc9b
Takahashi K, Saleh M, Penn RD, Hatsopoulos NG (2011) Propagating waves in human motor cortex. Front Hum Neurosci 5:40. https://doi.org/10.3389/fnhum.2011.00040
Takahashi K, Sanggyun K, Coleman TP, Brown KA et al (2015) Large-scale spatiotemporal spike patterning consistent with wave propagation in motor cortex. Nat Commun 6:7169. https://doi.org/10.1038/ncomms8169
Tamura Y, Ogawa H, Kapeller C, Prueckl R, Takeuchi F, Anei R et al (2016) Passive language mapping combining real-time oscillation analysis with cortico-cortical evoked potentials for awake craniotomy. J Neurosurg. https://doi.org/10.3171/2015.4.JNS15193
Thiel A, Zumbansen A (2016) The pathophysiology of post-stroke aphasia: a network approach. Restorative Neurol Neurosci 34(4):507–518. https://doi.org/10.3233/RNN-150632
To WT, De Ridder D, Hart J Jr, Vanneste S (2018) Changing brain networks through non-invasive neuromodulation. Front Hum Neurosci 12:128. https://doi.org/10.3389/fnhum.2018.00128
Töpper R, Mottaghy FM, Brugmann M, Noth J, Huber W (1998) Facilitation of picture naming by focal transcranial magnetic stimulation of Wernicke’s area. Exp Brain Res 121:371–378. https://doi.org/10.1007/s002210050471
Turi Z, Alekseichuk I, Paulus W (2018) On ways to overcome the magical capacity limit of working memory. PLoS Biol. https://doi.org/10.1371/journal.pbio.2005867
Turkeltaub PE, Messing S, Norise C, Hamilton RH (2011) Are networks for residual language function and recovery consistent across aphasic patients? Neurology 76:1726–1734. https://doi.org/10.1212/WNL.0b013e31821a44c1
Turkeltaub PE, Swears MK, D’Mello AM, Stoodley CJ (2016) Cerebellar tDCS as a novel treatment for aphasia? Evidence from behavioral and resting-state functional connectivity data in healthy adults. Restorative Neurol Neurosci 34(4):491–505. https://doi.org/10.3233/RNN-150633
Venkov NA, Coombes S, Matthews PC (2007) Dynamic instabilities in scalar neural field equations with space-dependent delays. Physica D 232:1–15
Voigt MB, Yussuf PA, Kral A (2018) Intracortical microstimulation modulates cortical induced responses. J Neurosci 38(36):7774–7786. https://doi.org/10.1523/JNEUROSCI.0928-18.2018
Weaver J (2013) How brain waves help us make sense of speech. PLoS Biol 11(12):e1001753
Whalen AJ, Kadji YXH, Dahlem MA, Gluckman BJ, Schiff SJ (2018) Control of spreading depression with electrical fields. Sci Rep 8:8769. https://doi.org/10.1038/s41598-018-26986-1
Wilson HR, Cowan JD (1973) A mathematical theory of the functional dynamics of cortical and thalamic nervous tissue. Kybernetik 13:55–80
Wortman-Jutt S, Edwards DJ (2017) Transcranial direct current stimulation in poststroke aphasia recovery. Stroke. https://doi.org/10.1161/STROKEAHA.116.015626
Wright J, Macefield VG, vanSchaik A, Tapson JC (2016) A review of control strategies in closed-loop neuroprosthetic systems. Front Neurosci 10:312. https://doi.org/10.3389/fnins.2016.00312
Wu JY, Huang X, Zhang C (2008) Propagating waves of activity in the neocortex: what they are, what they do. Neuroscientist 14(5):487–502. https://doi.org/10.1177/1073858408317066
Yourganov G, Fridriksson J, Rorden C, Gleichgerrcht E, Bonilha L (2016) Multivariate connectome-based symptom mapping in post-stroke patients: networks supporting language and speech. J Neurosci 36(25):6668–6679. https://doi.org/10.1523/JNEUROSCI.4396-15.2016
Zanos S (2018) Closed-loop neuromodulation in physiological and translational research. Cold Spring Harb Perspect Med. https://doi.org/10.1101/cshperspect.a034314
Zhang H, Watrous AJ, Patel A, Jacobs J (2018) Theta and alpha oscillations are traveling waves in the human neocortex. Neuron. https://doi.org/10.1016/j.neuron.2018.05.019
Author information
Authors and Affiliations
Contributions
Abe and VV drafted the manuscript. Aba, SH and FV provided critical comments, references and edits. VV was supported by the “RUDN University Program 5-100”.
Corresponding author
Ethics declarations
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Additional information
Communicated by Benjamin Lindner.
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Appendix: Model and parameters for numerical simulations
Appendix: Model and parameters for numerical simulations
Consider the equation
where \( W_{a} \) and \( W_{i} \) are given by formulas (1.2), (1.3), respectively, in the approximation of large conductivity limit (\( q_{a} = q_{i} = \infty \)),
and
The stimulation function is considered in the form
I0(x) = i0 for x1 ≤ x ≤ x2, and I0(x) = i1 outside the interval [x1, x2].
Numerical simulations are carried out in a bounded interval with periodic boundary conditions.
Figure 6 shows snapshots of wave propagation in the normal tissue (left), in the damaged tissue (middle), and in the damaged tissue with stimulation. The values of parameters are as follows:
σ = 0.01, Sa(u) = Si(u) = arctan(hu). h = 20, L = 2, p = 6, q = 1, i0 = 0.6, i1 = 0.1. The damaged interval (green line) is [0.5, 1.07], w0 = 0.
Rights and permissions
About this article

Cite this article
Beuter, A., Balossier, A., Vassal, F. et al. Cortical stimulation in aphasia following ischemic stroke: toward model-guided electrical neuromodulation. Biol Cybern 114, 5–21 (2020). https://doi.org/10.1007/s00422-020-00818-w
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00422-020-00818-w