
Abstract
The energy expended to transport the body over a given distance (C, the energy cost) increases with speed both on land and in water. At any given speed, C is lower on land (e.g., running or cycling) than in water (e.g., swimming or kayaking) and this difference can be easily understood when one considers that energy should be expended (among the others) to overcome resistive forces since these, at any given speed, are far larger in water (hydrodynamic resistance, drag) than on land (aerodynamic resistance). Another reason for the differences in C between water and land locomotion is the lower capability to exert useful forces in water than on land (e.g., a lower propelling efficiency in the former case). These two parameters (drag and efficiency) not only can explain the differences in C between land and water locomotion but can also explain the differences in C within a given form of locomotion (swimming at the surface, which is the topic of this review): e.g., differences between strokes or between swimmers of different age, sex, and technical level. In this review, the determinants of C (drag and efficiency, as well as energy expenditure in its aerobic and anaerobic components) will, thus, be described and discussed. In aquatic locomotion it is difficult to obtain quantitative measures of drag and efficiency and only a comprehensive (biophysical) approach could allow to understand which estimates are “reasonable” and which are not. Examples of these calculations are also reported and discussed.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Maximal swimming speed is the greater the higher the maximal metabolic power of the swimmer and the lower his/her energy cost (C). In turn: (1) metabolic power can be derived from aerobic or anaerobic (lactic and alactic) energy sources and (2) C depends on overall efficiency, propelling efficiency and hydrodynamic resistance. All the factors that are known to influence C (e.g., speed, stroke, age, sex, training …) act through their effects on (active) drag, (propelling) efficiency or both. A conceptual diagram of the key determinants of the energy cost of swimming is reported in Fig. 1. Understanding the complex interplay between all these factors, is the major aim of this review.

Conceptual diagram of the key determinants of the energy cost of swimming (C). C is calculated as the ratio between metabolic power (\(\dot{E}\)) and swimming speed. In turn, \(\dot{E}\) can be derived from aerobic (\(\dot{E}_{\text{Aer}}\)) or anaerobic (lactic and alactic, \(\dot{E}_{\text{Anl}}\) and \(\dot{E}_{\text{AnAl}}\)) energy sources: these could be considered the “metabolic determinants” of C; the “mechanical” determinants of C are (essentially) drag and propelling efficiency. C is indeed larger the larger the (active) drag (Fd) which can be considered as the sum of three components: form, friction, and wave drag. The power to overcome drag (\(\dot{W}_{\text{d}}\)) can be calculated by multiplying Fd by the swimming speed. \(\dot{W}_{\text{d}}\) is only a fraction of the total mechanical power (\(\dot{W}_{\text{tot}}\)) provided by the muscles and propelling efficiency (hP) indeed indicates the efficiency of this transformation. All the factors that are known to influence C (e.g., speed, stroke, age, sex, training …) mainly act through their effects on Fd, ηP or both. See text for details
Energy cost
In swimming, as well as in other cyclic forms of human locomotion (in water and on land), metabolic power (the energy expended in the unit of time, \(\dot{E}\)) increases exponentially as a function of speed (that is indeed a “measure” of exercise intensity). The energy cost (C) is a parameter that takes into account differences in energy expenditure, at a given speed, between subjects/modes of locomotion; it is calculated by “normalizing” \(\dot{E}\) to the locomotion speed (v):
If the v is expressed in m s−1 and \(\dot{E}\) in kJ · s−1, C results in kJ · m−1 (e.g., di Prampero 1986). C can thus be defined as:
-
the metabolic power needed to move at a given speed,
-
the metabolic energy needed to cover a given distance.
In more general terms, C represents the energy expended, at a given speed, not as a function of time but as a function of the distance covered. Indeed, C is also defined as the cost of transport and is generally reported normalized by the body mass of the subject (kJ · m−1. kg−1). This normalization is important/meaningful in terrestrial locomotion where body mass influences the amount of mechanical/metabolic energy that has to be expended to oppose the effects of gravity; in aquatic locomotion other anthropometric characteristics play a more important role in determining energy expenditure, as an example stature and arm span (e.g., Chatard et al. 1990c) or “leg sinking” torque (e.g., Pendergast et al. 1977). In swimming studies it is, thus, more common to report C in kJ · m−1 or to normalize it to body surface area (a parameter that influences hydrodynamic resistance, see Sect. “Hydrodynamic resistance (Fd) and power to overcome drag (\(\dot{W}_{\text{d}}\))”, instead of body mass (e.g., Pendergast et al. 1977).
How to measure \(\dot{E}\) will be indicated and discussed in Sect. “Metabolic power”; briefly, we can anticipate that the large majority of aquatic locomotion studies report data of submaximal energy expenditure after steady state conditions are attained, a condition for which is not necessary to estimate the contribution of the anaerobic energy sources, the determination of which is based on several (and sometimes debated) assumptions.
It goes without saying that also measuring the aerobic component of \(\dot{E}\) (e.g., oxygen uptake: \(\dot{V}{\text{O}}_{2}\)) is not an easy task in the aquatic environment; in the specific case of swimming, experiments are generally performed in swimming pools and therefore, for practical reasons, the starting dive should be avoided and the swimmers must perform open turns (and cannot thus perform a proper underwater gliding phase); this means that the energy needed for starts and turns is not taken into account with consequences often ignored/not discussed in the literature. Indeed, “neglecting starts and turns” has important consequences on the estimates of the overall energy expenditure of swimming. The percentage contribution of the time spent in starts and turns, respectively, decreases and increases with race distance and, in long distance events, up to 30% of total race time is spent in turning (e.g., Lyttle and Blanksby 2011). The capability to perform efficient turns is thus expected to have a remarkable effect on the overall energy expenditure (\(\dot{E}\)) of swimming, especially in short course pools, although this is still a relatively unexplored field of study.
Swimming speed in Eq. 1 corresponds to the “clean swimming speed” that can be represented by a single, constant, average value. Indeed, v is generally determined in the central section of the pool where the acceleration caused by the push on the pool edge during the turn is over and the deceleration in view of the next turn is not started yet. The swimmer’s speed, in starts and turns, is far larger than clean swimming speed: typical average velocities for elite male swimmers (in front crawl) in the start phase (first 15 m) are of about 3 m·s−1, while clean swimming speed is in the order of 1.8–2 m·s−1. The faster than average speed during turns is the reason for the faster times in short (25 m) compared to long courses (50 m) races.
Even if the major changes in speed during a race are observed during starts and turns, clean swimming speed cannot be considered “constant” since it shows intra-cyclic variations which differ according to the swimming style and speed (e.g., Craig and Pendergast 1979); indeed in all cyclic forms of locomotion, in land and in water, propulsive forces are not constant but impulsive and this results in intra-cyclic variations in speed. It is however assumed that the average effective force (over one cycle) produces the average (e.g., “constant”) locomotion speed. The effects of the intra-cyclic variations in speed on the energy expenditure of swimming will be briefly discussed in Sect. “Intra-cyclic velocity variations”.
The importance of C in determining performance can be appreciated by rearranging Eq. 1 and applying it to maximal conditions (di Prampero 1986):
Equation 2 indicates that maximal speed (vmax) depends on the ratio of maximal metabolic power (\(\dot{E}_{{\max} }\)) to C; hence, for a given athlete (for a given value of \(\dot{E}_{{\max} }\)), the maximal speed he/she can attain in different forms of land/water locomotion is essentially set by the value of C of that form of locomotion. This formally explains data reported in Fig. 2.

The energy cost of locomotion in water (continuous lines) increases with a steeper rate than that of land locomotion (dashed lines); the use of locomotory tools on land (skates: dark grey dashed line; bicycles: grey dashed line) and in water (gondola: dark grey continuous line; kayak: grey continuous line) allows for a decrease in energy expenditure compared to locomotion without tools (walking and running: black dashed line; front crawl swimming: black continuous line). For a given subject (characterized by a given \(\dot{V}{\text{O}}_{2{\max} }\)) the maximal speed that can be attained in these different modes of locomotion is thus different (lower in water than on land) and this, depends indeed on the energy cost of each specific form of locomotion (larger in water than on land). Land locomotion (as reported by di Prampero 1986): running: C = 270 + 0.72 · v2; speed skating on ice (“dropped posture”): C = 70 + 0.79 · v2; cycling (standard racing bike, “dropped posture”): C = 13 + 0.77 · v2. Water locomotion: front crawl swimming: C = 228 × 100.488 v (Capelli et al. 1998); sculling a venetian gondola: C = 155 · v1.67 (Capelli et al. 1990); kayaking: C = 20 · v2.26 (Zamparo et al. 1999)
Data reported in Fig. 3 represent, schematically, the energy cost during a given form of locomotion (e.g., front crawl swimming) in three subjects with different proficiency level (recreational, good swimmer and elite). In analogy with Fig. 2, this figure shows that differences in energy cost can be observed among subjects in a given form of locomotion: C increases with a steeper rate in less experienced swimmers. The differences in the maximal attained speed, also in this case, depend on maximal metabolic power and on the energy cost of locomotion: elite swimmers have both a larger \(\dot{E}_{{\max} }\) and a lower C, for these reasons they can reach faster speeds than recreational swimmers. An athlete can thus improve his/her performance (can increase his/her vmax) by increasing his/her maximal metabolic power (\(\dot{E}_{{\max} }\), the numerator of Eq. 2) and/or by decreasing his/her energy cost (C, the denominator of Eq. 2).

Energy cost of front crawl swimming for a recreational swimmer (continuous line) a good swimmer (dotted line) and an elite swimmer (dashed line) (adapted from Holmer 1972); C is larger, at a given submaximal speed, and increases with a steeper rate, in less experienced swimmers; the swimmers with better technical skills can thus reach larger values of speed (see text for details)
Maximal metabolic power depends on the availability of aerobic and anaerobic (lactic and alactic) energy sources. The larger these sources (i.e., the capacity, in kJ) and the faster they can be utilized the larger the metabolic power (kJ·s−1 = kW), and hence vmax. As shown by Capelli (1999), an improvement in a subject’s vmax can more easily be obtained by a reduction of C rather than by an (equal) increase in metabolic power (in either of its components, aerobic or anaerobic).
To reduce C it is thus important to know which factors determine/influence this parameter. As it will be formally demonstrated later on (Sect. “Mechanical power and efficiency”), in water locomotion C depends on hydrodynamic resistance (Fd), propelling efficiency (ηP) and overall efficiency (ηO):
This equation indicates that maximal speed in swimming depends on the interplay among all these factors: as an example, if two athletes have the same \(\dot{E}_{{\max} }\), the swimmer with higher propelling efficiency and lower hydrodynamic resistance (and hence with a lower C) will outswim the swimmer with a poor ηP and a large Fd (and hence with a higher C). On the other hand, a swimmer with an elevated \(\dot{E}_{{\max} }\) could outswim a peer with a better C but characterized by a lower maximal aerobic and/or anaerobic power.
Equation 4 is, as an example, useful to understand: (1) why a given athlete (with a given \(\dot{E}_{{\max} }\)) could reach lower speeds in breaststroke than in front crawl: probably because of a larger Fd and a lower ηp in the former compared to the latter; or (2) why adults swim faster than children: probably because of their larger \(\dot{E}_{{\max} }\) and their larger ηP (better technical skills) despite their larger Fd.
Equation 4 is, also, useful to understand why a given athlete (with a given \(\dot{E}_{{\max} }\)) could reach lower speeds in water compared to land locomotion or in swimming compared to boat locomotion: probably because of a larger Fd and a lower ηp in the former compared to the latter.
In the following sections the methods proposed in the literature to calculate/estimate the energy cost of swimming and its determinants will be briefly discussed. After that the studies in the literature reporting data of energy cost in swimming will be reviewed and discussed in the framework of Eq. 3.
Metabolic power
The large majority of the studies in the literature report data of submaximal energy expenditure essentially because, in this case, it is not necessary to estimate the contribution of the anaerobic energy sources, the determination of which is based on several assumptions. This is a remarkable limitation because swimming races are not carried out at submaximal speeds.
Aerobic contribution (\(\dot{E}_{\text{Aer}}\))
Metabolic power (\(\dot{E}\)) at constant, submaximal (aerobic) speeds can be calculated by measuring steady state oxygen consumption by standard open circuit techniques. In these conditions energy expenditure is based on “aerobic energy sources” (\(\dot{E}\) = \(\dot{E}_{\text{Aer}}\)) and the rate of ATP utilization is equal to the rate of ATP re-synthetized by lipid and glucose oxidation in the Kreb’s cycle (e.g.,, di Prampero 1981).
Douglas bags, balloons or portable metabolimeters can be utilized to measure steady state oxygen consumption during exercise but often this is a challenging task in the aquatic environment. As indicated above, when these devices are utilized, diving starts should be avoided and the swimmers should perform open turns. The use of snorkels to collect expired air thus affects turning time as well as some swimming movements (e.g., it virtually eliminates body roll) but the most recent models were shown not to affect hydrodynamic resistance (e.g., Baldari et al. 2013; Ribeiro et al. 2016).
Given the objective difficulties of assessing steady state oxygen consumption in water in some studies an alternative method (the “back extrapolation” technique) is utilized where breath by breath oxygen consumption is recorded at the mouth not during the exercise itself but immediately after it, during the first 30–60 s of recovery (e.g., Zamparo et al. 2000, 2008b; Rodriguez 2000; Keskinen et al. 2003; Costill et al. 1985; Montpetit et al. 1981).
Swimming competitions could be disputed over distances from 50 to 1500 m (in the pool), the corresponding world records ranging from about 20 s to 15 min. The 400 m distance is generally taken as the competition eliciting the \(\dot{V}{\text{O}}_{2{\max} }\) of the swimmer (it lasts about 4 min in the front crawl). Races over longer distances are mainly based on aerobic energy sources and are swum at a fraction of \(\dot{V}{\text{O}}_{2{\max} }\) that is smaller the longer the race duration (see Sect. “Open water swimming and ultra-endurance races”) whereas races over shorter distances (50, 100 and 200 m) are based on a substantial contribution of the anaerobic energy sources, the more so the shorter the race duration (e.g., Toussaint and Hollander 1994; di Prampero et al. 2011; Capelli et al. 1998); in this case the contribution of the anaerobic (lactic and alactic, \(\dot{E}_{\text{Anl}}\) and \(\dot{E}_{\text{AnAl}}\)) energy sources to metabolic power (\(\dot{E}\)) could play an important role and could not be neglected; in this case:
Anaerobic lactic contribution (\(\dot{E}_{\text{Anl}}\))
The lactic contribution to \(\dot{E}\) can be estimated from the net increase of blood lactate concentration (Lab) assessed at the end of exercise: \(\dot{E}_{\text{Anl}}\) = β Lab · t−1 where t is the exercise duration and β is the energy equivalent of lactate (e.g., di Prampero 1981), thus, \(\dot{E}_{\text{Anl}}\) decreases along with the duration of exercise, independently of the absolute peak lactate concentration in blood. As an example, as shown by Capelli et al. (1998) in maximal front crawl trials over a distance of 100 m, \(\dot{E}_{\text{Anl}}\) = 1.29 kW (about 47% of \(\dot{E}\)) whereas over the 200 m distance \(\dot{E}_{\text{Anl}}\) = 0.48 kW (about 25% of \(\dot{E}\)) even if in both cases Lab is similar (e.g., 10–11 mM). For a detailed analysis of the pro and cons of this method the reader is referred to the papers of di Prampero and Ferretti (1999), Capelli et al. (1998) and di Prampero (2003).
Anaerobic alactic contribution (\(\dot{E}_{\text{AnAl}}\))
A direct assessment of the anaerobic alactic energy sources through invasive methods (e.g., muscle extracts) is impracticable so that non-invasive alternatives should be used. The alactic contribution to \(\dot{E}\) can be estimated based on PCr breakdown at the beginning of exercise: \(\dot{E}_{\text{AnAl}}\) = PCr (1 − e−t/τ) · t−1, where PCr is the phosphocreatine concentration at rest (in the contracting muscles), t is the exercise τduration and τ is the time constant of PCr splitting at work onset (about 30 s); the implicit assumption is thus made that the time constant of PCr breakdown at work onset is the mirror increment of the \(\dot{V}{\text{O}}_{2}\)-on response at the muscle level (e.g., Rossiter et al. 2002).
As is the case for \(\dot{E}_{\text{Anl}} ,\;\dot{E}_{\text{AnAl}}\) decreases along with the duration of exercise, independently of the absolute PCr concentration at rest. As an example, as shown by Capelli et al. (1998) in maximal front crawl trials over a distance of 100 m, \(\dot{E}_{\text{AnAl}}\) = 0.53 kW (about 20% of \(\dot{E}\)) whereas over the 200 m distance \(\dot{E}_{\text{AnAl}}\) = 0.27 kW (about 14% of \(\dot{E}\)) (Capelli et al. 1998). For a detailed analysis of the pro and cons of this method the reader is referred to the papers of Capelli et al. (1998) and di Prampero (2003).
Another method to estimate the anaerobic alactic contribution is based on the analysis of the fast component of the \(\dot{V}{\text{O}}_{2}\)-off kinetics (e.g., di Prampero et al. 1970; Beneke et al. 2002) since in the post-exercise period part of the oxygen debt is necessary to rebuild the high energy substrates splitted at the beginning of exercise (e.g., di Prampero and Ferretti 1999). This method was applied by Sousa et al. (2013) in their analysis of the 200 m front crawl race; these authors have shown that, despite the existence of some caveats regarding both methods (PCr breakdown or fast-off component), they yield similar values of \(\dot{E}_{\text{AnAl}}\).
Finally, the anaerobic energy contribution can be calculated by means of the accumulated oxygen deficit method, originally proposed by Medbø et al. (1988) and applied to swimming by several authors (e.g., Ogita et al. 2003; Reis et al. 2010; Peyrebrune et al. 2014). This method estimates values of \(\dot{E}_{\text{AnAl}}\) somewhat lower than in the two previous cases but, as pointed out by Peyrebrune et al. (2014), these differences could be attributed, besides to methodological issues, to the competitive level of the participants, their specialization and, above all, to differences in the exercise duration (see below). The differences in the relative contribution of the energy systems in short events reflects the difficulties in estimating/computing \(\dot{E}_{\text{Anl}}\) and \(\dot{E}_{\text{AnAl}}\) but, compared to less recent literature, these disparities have been greatly reduced (Rodriguez and Mader 2011).
The “Wilkie’s approach”
Another approach reported in the literature to estimate metabolic power (\(\dot{E}\)) at maximal speed was originally proposed by Wilkie (1980) and later applied to land and water locomotion (e.g., di Prampero 1986; Capelli et al. 1998; Zamparo et al. 1999, 2000). With this approach, during all-out test of short duration (less than 2–3 min), \(\dot{E}\) can be calculated as:
where α is the energy equivalent of O2 (e.g., 20.9 kJ · lO −12 , for a respiratory exchange ratio of 0.96), τ is the time constant (s) wherewith \(\dot{V}{\text{O}}_{2{\max} }\) is attained at the onset of exercise at the muscular level (about 30 s, Rossiter et al. 2002), \(E_{\text{AnS}}\) is the amount of energy derived from anaerobic energy utilization, t is the time of performance (s) and \(\dot{V}{\text{O}}_{2{\max} }\) is the net maximal oxygen uptake (above resting values). The first term of Eq. 6 (\(E_{\text{AnS}}\)· t−1) is the sum of \(\dot{E}_{\text{Anl}}\) and \(\dot{E}_{\text{AnAl}}\) and can be calculated as described in Sects. “Anaerobic lactic contribution (\(\dot{E}_{\text{Anl}}\))” and “Anaerobic alactic contribution (\(\dot{E}_{\text{AnAl}}\))”. The third term of Eq. 6. is a measure of the O2 debt incurred at the onset of exercise and, therefore, the aerobic contribution to metabolic power (\(\dot{E}_{\text{Aer}}\)) can be calculated as the sum of the second and third term of Eq. 6: \(\dot{E}_{\text{Aer}}\) increases with the duration of exercise becoming equal to α\(\dot{V}{\text{O}}_{2{\max} }\) in maximal trials lasting at least 2–3 min (e.g., more than 4–5τ). \(\dot{E}_{\rm Aer}\) calculated in this way was shown to be the same as that directly measured by means of a metabolimeter (e.g., Zamparo et al. 1999).
This approach to estimate \(\dot{E}\) (by means of Eq. 6) is discussed in detail by di Prampero (2003) and Capelli et al. (1998) and should be strictly applied only to “square wave” exercises of intensity close or above maximal aerobic power where a true steady state of oxygen uptake cannot be attained and where energy contributions other than the aerobic one cannot be neglected.
The percentage contribution of the aerobic (\(\dot{E}_{\text{Aer}}\)) and anaerobic (\(\dot{E}_{\rm Anl} ,\;\dot{E}_{\text{AnAl}}\)) energy sources to total metabolic power, respectively increases and decreases as a function of exercise duration (e.g., Capelli et al. 1998; Rodriguez and Mader 2011; Toussaint and Hollander 1994). When calculated with the above methods/equations, these contributions are almost independent of sex, skill, and mode of locomotion and depend essentially upon the duration of the exercise (t). As an example, for all-out efforts of the same duration (about 1 min, e.g., over a distance of 100 m in swimming and 250 m in kayaking) the contribution of the aerobic and anaerobic lactic energy sources is of about 40% and that of the anaerobic alactic energy sources is of about 20% of total metabolic power expenditure (e.g., Capelli et al. 1998; Zamparo et al. 1999).
Finally, metabolic power at race pace can also be estimated by extrapolating the \(\dot{E}\)vs. v relationship, as determined at submaximal speeds, to race speeds. Even if such extrapolations should be taken with care, it can be reasonably assumed that if data calculated at maximal speed and measured at submaximal speeds lay on the same \(\dot{E}\)vs. v relationship this is evidence of correct estimation of the supramaximal data (see Sect. ““Reasonable” assumptions”).
To summarize: \(\dot{E}_{\text{Aer}}\), \(\dot{E}_{\text{Anl}}\) and \(\dot{E}_{\text{AnAl}}\) can be considered the “metabolic determinants of C” because, once \(\dot{E}\) is known C can be calculated, at any given speed (see Eq. 1). To understand the differences in C between subjects/among conditions it is, however, important to know which other parameters could influence C. As indicated by Eq. 3, these are: hydrodynamic resistance (Fd), propelling efficiency (ηp), and overall efficiency (ηO). In the following sections these “mechanical determinants of C” will be defined and discussed.
Mechanical power and efficiency
The increase in metabolic power (\(\dot{E}\)) with speed reflects the increase in (total) mechanical power output (\(\dot{W}_{\text{tot}}\)) that the muscles have to provide to sustain that speed. As for land locomotion, the total mechanical power of aquatic locomotion (\(\dot{W}_{\text{tot}}\)) can be considered as the sum of two terms (e.g., Cavagna and Kaneko 1977): the power needed to accelerate and decelerate the limbs with respect to the centre of mass (the internal power, \(\dot{W}_{\text{int}}\)) and the power needed to overcome external forces (the external power, \(\dot{W}_{\text{ext}}\)):
In aquatic locomotion \(\dot{W}_{\text{ext}}\) can be further partitioned into the power to overcome drag (\(\dot{W}_{\text{d}}\)), that contributes to useful thrust, and the power that does not contribute to thrust \((\dot{W}_{\text{k}} )\):
Both \(\dot{W}_{\text{d}}\) and \(\dot{W}_{\text{k}}\) give water kinetic energy but only \(\dot{W}_{\text{d}}\) effectively contributes to propulsion (e.g., Alexander 1977, Daniel 1991). The efficiency with which the external mechanical power produced by the swimmer is transformed into useful mechanical power is termed Froude (theoretical) efficiency (ηF) and is given by:
The efficiency with which the overall mechanical power produced by the swimmer is transformed into useful mechanical power (to overcome hydrodynamic resistance) is termed propelling efficiency (ηP) and is given by:
The efficiency with which the overall mechanical power produced by the swimmer is transformed into external mechanical power is termed hydraulic efficiency (ηH) and is given by:
Froude, propelling, and hydraulic efficiency are of utmost importance in water locomotion since they indicate the capability of the swimmer to transform at best his/her muscular power in power useful for propulsion: they could vary from 0 to 1 (all power is utilized for propulsion). If the internal power (\(\dot{W}_{\text{int}}\)) is nil or negligible: ηH = 1 and ηF = ηP. Propelling, hydraulic and Froude efficiency refer to the mechanical partitioning only but other two efficiencies should be defined in water locomotion and these take into account also metabolic energy expenditure. The efficiency with which the metabolic power input (\(\dot{E}\)) is transformed into useful mechanical power output (\(\dot{W}_{\text{d}}\)) is termed performance (or drag) efficiency (ηd) and is given by:
The efficiency with which the metabolic power input (\(\dot{E}\)) is transformed into mechanical power output (\(\dot{W}_{\text{tot}}\)) is termed overall (or gross or mechanical) efficiency (ηO) and is given by:
It follows that: ηd = ηO · ηP. Combining Eqs. 10 and 13 one obtains:
which, at any given speed, relates the metabolic power input (\(\dot{E}\)) with the power needed to overcome hydrodynamic resistance (\(\dot{W}_{\text{d}}\)) and with propelling (ηP) and overall (ηO) efficiency. Since, for any given speed, C = \(\dot{E}\)/v and \(\dot{W}_{\text{d}}\) = Fd · v, it follows that C = Fd/(ηP · ηO) and this formally explains Eq. (3).
This partitioning of the efficiencies in human swimming (see Fig. 4) is based on the flow diagram of the steps of energy conversion in steady state aquatic locomotion proposed by Daniel (1991): metabolic energy is converted to contractile work, only a fraction of it goes to moving fluid and of this work, only a fraction goes into useful thrust. It can thus be possible, by knowing the partitioning between useful and non-useful work components, at each step of this diagram, to calculate C (at a given swimming speed) starting from measures of useful thrust in water at the same speed. Some examples of these calculations are reported in Sect. ““Reasonable" assumptions”.

Flow diagram of the steps of energy conversion in steady state aquatic locomotion (adapted from Daniel 1991): metabolic power (\(\dot{E}\)) is converted into mechanical power output (\(\dot{W}_{\text{tot}}\)) but only a fraction of it goes into moving fluid (external power, \(\dot{W}_{\text{ext}}\)) and of this, only a fraction goes into useful thrust (e.g., into power to overcome drag, \(\dot{W}_{\text{d}}\)). See text for details
Some clarifications are needed in regard to these equations:
-
Efficiency is a dimensionless parameter and hence numerator and denominator should be expressed in the same units. Therefore, the efficiencies of aquatic locomotion can be calculated (as above) based on values of mechanical (\(\dot{W}\)) and metabolic (\(\dot{E}\)) power, in this case numerator and denominator represent the energy expended in the unit of time (e.g., J . s−1). When metabolic energy is expressed as a function of distance (e.g., to highlight the role of the energy cost, which is expressed in J · m−1) also the mechanical counterpart has to be expressed in the same units (J · m−1, work per unit of distance); this can be obtained by dividing \(\dot{W}\) to the speed (as \(\dot{E}\) is divided by the speed to obtain C).
-
The work, per unit distance, needed to overcome resistive forces (at a given speed) corresponds to the drag force (Fd), at that speed (J · m−1 = N). Indeed when this resistive force (Fd, N) is measured, at a given (constant) speed (v, m s−1), the power to overcome drag (\(\dot{W}_{\text{d}}\), W) is simply calculated as: Fd · v.
-
At constant speed the sum of propulsive forces must equal the sum of resistive forces; indeed, were a swimmer able to produce propulsive forces larger than the resistive ones he/she would accelerate. At constant speed thrust force (Ft) and drag force (Fd) should balance: the power needed to overcome drag (\(\dot{W}_{\text{d}}\)) and the power required to push the swimmer forward (\(\dot{W}_{\text{t}}\) = Ft · v) can be considered two sides of the same coin.
-
Only few studies have investigated so far internal work/power in swimming (e.g., Zamparo et al. 2002, 2005a; Gonjo et al. 2018); therefore data of mechanical power reported in the literature mainly refer to its external component (\(\dot{W}_{\text{tot}}\) = \(\dot{W}_{\text{ext}}\)); in these cases, the so called values of propelling efficiency are, therefore, values of Froude efficiency (and the implicit assumption is made that hydraulic efficiency = 1).
-
Only few studies have investigated so far work and efficiency of the lower limbs (e.g., Zamparo et al. 2002, 2005a); therefore, the values of Froude/propelling efficiency reported in the literature are, mostly, values of Froude efficiency of the arm stroke.
To summarize, the differences in \(\dot{E}\) (or in C) among swimmers (or locomotion modes in water), at any given speed and for a specific ηO, depend on differences in ηP and/or in \(\dot{W}_{\text{d}}\) (or \(\dot{W}_{\text{t}}\)). For a given locomotion mode, an increase in ηP and/or a decrease in \(\dot{W}_{\text{d}}\) (or \(\dot{W}_{\text{t}}\)) will lead to a decrease in \(\dot{E}\) (or in C) (Eq. 3) and this will allow for an increase in the maximal attainable speed (Eq. 4).
Hydrodynamic resistance (Fd) and power to overcome drag (\(\dot{W}_{\text{d}}\))
In aquatic locomotion metabolic energy is mainly expended to overcome hydrodynamic resistance (Fd), the more so the larger the speed. Air resistance is indeed negligible, in comparison to water resistance, and the Archimedes lift allows the swimmer to spend virtually no energy to remain at the water surface (gravitational work is much reduced compared to land locomotion). Fd can be considered as the sum of three components: form, friction, and wave drag.
1. Form (pressure) drag depends on the shape of the body and on its effects on the water flow; this component of drag increases with the square of swimming speed and prevails at low to moderate speeds (it is about 80% of total -passive and surface- drag at 1 m · s−1, e.g., Pendergast et al. 2005):
where ρ is water density, v is the swimming speed, Cd is the drag coefficient and A is the projected frontal area (in the direction of movement). Cd is a coefficient that takes into account the shape of the body in motion; smaller values of Cd are associated with elongated shapes oriented in the direction of flow (streamlined objects, e.g., Vogel 1994).
For partially submerged bodies (as is the case in swimming) A is the surface exposed to the water (e.g., the “wetted” area) and depends on:
-
the dimensions of the swimmer’s body (A is greater in heavier and taller swimmers),
-
the fraction of the body that is submerged (A is greater in more muscular, denser, individuals),
-
the body incline/the angle of attack (A is lowest when the body is oriented in the direction of flow),
-
the position of the body segments during the stroke cycle (A differs among swimming stroke).
The first three points indicate that pressure drag depends on the anthropometric characteristics of the subjects: C was indeed found to increase with body length, mass and surface area and to decrease with buoyancy (e.g., Chatard et al. 1985, 1990c; Kjendlie et al. 2004a, b). When C is scaled for body size, inter-subject differences tend to disappear (e.g., Kjendlie et al. 2004a, b; Ratel and Poujade 2009).
A way to take into account the effects of body size and density is to measure the “leg sinking torque” (T) a parameter related to the (static) position the body assumes when immersed in water: taller, heavier and less buoyant swimmers have larger values of T and are thus expected to require more energy to overcome drag forces. Indeed, men have greater values of T than women (and adults than children) and spend more energy per unit distance: at slow swimming speeds (0.9–1.1 m · s−1), about 70% of the variability of C is explained by the variability of T, regardless the sex, age, and technical level of a swimmer (e.g., Pendergast et al. 1977; Zamparo et al. 2008b).
At faster speeds the static and dynamic position in water are no longer related; indeed, body incline and A decrease with increasing speed (e.g., Zamparo et al. 2009) and at 1.6 m · s−1 (and above) no relationship between T and C could be observed anymore (Zamparo et al. 2000). As discussed by Yanai and Wilson (2008), the forces caused by the hands and legs movements during swimming affect buoyancy (and T) differently than when the body floats motionless in water. As an example, Yanai (2001) showed that, at high swimming speeds (e.g., 1.6 m · s−1), the arm movements generate a leg raising rather than a leg sinking effect in front crawl; thus, Yanai and Wilson (2008) challenged the interpretation that the larger C in male adults compared to children (and females) could be attributed to the kicking actions needed to actively counteract the tendency of the legs to sink, this compensation costing some extra energy (Pendergast et al. 1977; Zamparo et al. 2008b; Kjendlie et al. 2004a) and suggest that the differences in C have to be attributed to differences in body size and composition only.
The fourth point is related to the differences between active and passive drag (see end of this section); during swimming A is larger than in passive conditions (e.g., when the swimmer is passively towed in water in a streamlined position) because of the movements of the trunk and the limbs during the stroke cycle (e.g., Gatta et al. 2015). A “smooth” and efficient stroke (that minimizes A during the swimming motion) can thus be expected to lead to a reduction in (active) drag (and hence of C, as indicated above). Indeed, the differences in C reflect the differences in A among strokes (see Sect. “Speed and stroke”).
2. Friction drag depends on the surface characteristics of the body and on the features of the boundary layer (e.g., a layer of more or less stationary fluid immediately surrounding the body in motion); this component of drag is rather unaffected by swimming speed (e.g., Wilson and Thorp 2003). To reduce friction drag, swimmers shave their legs, as do cyclists; the effects of hair removal on the physiological responses in swimming were studied, as an example by Sharp and Costill (1989) who observed, after shaving, a reduction in the rate of velocity decay during a prone glide after a maximal underwater leg push-off and a decrease in oxygen uptake during a 400 m breaststroke swim at 90% effort. Full body swimsuits were developed to reduce this component of drag (that indeed decreases in proportion to the amount of body surface covered) even if drag reduction seems to be attributable to a reduction in pressure, rather than friction, drag (e.g., Mollendorf et al. 2004; Cortesi et al. 2014). As shown, as an example, by Chatard and Wilson (2008), using full body swimsuits allows to significantly reduce (passive) drag and this is associated with a decrease in performance time and in the energy cost of swimming, as it can be expected based on Eqs. 3 and 4.
3. Wave drag depends on the wave formation at the water surface; this component of drag increases with swimming speed and is prevailing at the highest speeds attained in swimming competitions (about 60% of total -passive and surface-drag at 1.7 m · s−1; Vennell et al. 2006). Wave drag decreases in proportion to the level of immersion of the body: at the surface it can be up to 2.4 times than when fully immersed, as demonstrated by Vennell et al. (2006) by towing a mannequin in a swimming flume and, more recently, by Tor et al. (2015) by towing a swimmer in a pool. To our knowledge no studies have been conducted so far to assess the effects of wave drag on the energy expenditure/energy cost of swimming perhaps because to single out the effects of this component of drag is not an easy task. The effects on energy expenditure could, however, be inferred from Eqs. 3 and 4 since speed largely increases when swimmers move underwater (as an example after starts and turns). Another (indirect) approach to quantify the effects of wave drag on energy expenditure is to investigate the effects of drafting distance between swimmers. As is the case for cycling, a sheltered position (when swimming in the wake of another swimmer) allows for a significant decrease in drag and energy cost depending on the distance from the leading swimmer when drafting (e.g., Basset et al. 1991, Chatard and Wilson 2003; Janssen et al. 2009). Morphology influences wave drag: a taller swimmer generally swims faster because he/she creates less wave resistance at a given speed and has a greater potential for maximal speed (e.g., Kjendlie and Stallman 2011). This is the same for boat locomotion: the longer the scull the faster the maximal attainable speed (see Sect. “Locomotory tools and boat locomotion”). As pointed out by Kjendlie and Stallman (2011) a greater stature is a determinant in sprint swimming but is not so critical for long distance races where speed, and therefore wave resistance, is lower and the leg sinking torque is greater.
Hydrodynamic resistance determined by towing a non-swimming subject through the water is defined as passive drag. Data of passive drag reported in the literature are fairly consistent even in different experimental designs (e.g., Havriluk 2005, 2007; Scurati et al. 2019). In the gliding phases of a race (after starts and turns) it is important to maintain the best hydrodynamic position (to reduce as much as possible passive drag). Since these phases correspond to 10–25% of the race, depending on race distance and pool length (e.g., Lyttle and Blanksby 2011), it is not surprising that passive drag was found to be a good evaluator of swimming performance (e.g., Chatard et al. 1990a, b).
When passive drag is measured, no propulsion is provided by the swimmer whereas, during swimming, the limbs movements create propulsion but also create additional resistance, thus, active drag is expected to be significantly larger than passive drag. Measuring active drag is clearly more interesting/informative that measuring passive drag if one wants to understand the effects of hydrodynamic resistance on energy expenditure but how to measure active drag is a controversial issue in swimming literature (e.g., Wilson and Thorp 2003; Toussaint et al. 2004; Havriluk 2007; Zamparo et al. 2009). The reader is referred to Zamparo et al. (2011) for some historical considerations about this point. The most recent papers (based on paradigms/methods, proposed after 2011) indicate that active drag is from 1.5 to 2.5 times larger than passive drag (Formosa et al. 2011; Gatta et al. 2015; Narita et al. 2017). As is the case for energy expenditure/energy cost, active drag is mainly studied in front crawl swimming and there is paucity of data regarding the other swimming strokes.
Internal and “wasted” power (\(\dot{W}_{\text{int}}\) and \(\dot{W}_{\text{k}}\))
There are not many data in the literature regarding \(\dot{W}_{\text{int}}\) and \(\dot{W}_{\text{k}}\); \(\dot{W}_{\text{k}}\) was (and can be) estimated based on measures of ηF, \(\dot{W}_{\text{d}}\) and \(\dot{W}_{\text{ext}}\) (e.g., Zamparo et al. 2005a). Relatively more attention, in the literature, was given to \(\dot{W}_{\text{int}}\): as for land locomotion it was found to depend on the movement frequency and on the inertial proprieties of the moving limbs. In the front crawl \(\dot{W}_{\text{int}}\) is larger the larger the speed and is larger in the leg kick than in the arm stroke (e.g., Zamparo et al. 2005a) and this provides a quantitative mechanical explanation of the suboptimal hydraulic efficiency in lower compared to upper limbs motion. As for land locomotion, a reduction in movement frequency (at a given speed) is thus expected to lead to a decrease in energy expenditure (e.g., Minetti et al. 1995). Indeed, as shown by Zamparo et al. (2002, 2005a) the decrease in stroke frequency (and hence in \(\dot{W}_{\text{int}}\)) when fins are used allows to sensibly reduce the energy cost/energy expenditure of swimming. This is also confirmed by studies on hand paddles that also induce (at a given speed) both a decrease in stroke frequency and in the energy expenditure of swimming (e.g., Ogita and Tabata 1993; Tsunokawa et al. 2019). The decrease in stroke frequency when fins/hand paddles are used, is also associated to an increase in propelling (or Froude) efficiency (see Sect. “Locomotory tools and boat locomotion”); according to Eq. 3 this formally explains why C decreases when movement frequency decreases (e.g., when Froude efficiency increases).
The internal power (\(\dot{W}_{\text{int}}\)) was also found to increase with stroke frequency in the breaststroke (Lauer et al. 2015); in this case the limbs action is synchronous, and no major differences are observed between upper and lower limbs. Interestingly, these authors found that the swimmers have the ability to moderate the increase in \(\dot{W}_{\text{int}}\) at increasing stroke frequencies (a strategy of energy minimization) by reducing the angular excursion of the heaviest segments (thighs and trunk) in favour of the lighter ones (shanks and forearms).
As pointed out by Lauer et al. (2015) and discussed in many studies regarding these matters in land locomotion (e.g., Kautz and Neptune 2002), internal and external power might be coupled to some extent and cannot be regarded as independent, additive components (as depicted in Fig. 4). Moreover in water, compared to land locomotion, the inertia of the “added mass of water” should be accounted for in the calculations of \(\dot{W}_{\text{int}}\); indeed, an immersed body behaves as it was heavier and additional mechanical work is required against both the inertia of the body and that of the displaced fluid (Lauer et al. 2015). These matters need to be further investigated in swimming studies.
Drag efficiency and overall efficiency (ηd and ηO)
Even if the methods developed so far to determine active and/or passive drag are quite debated in the literature, they consistently indicate that less than 10% of metabolic power input can be transformed into useful mechanical power output (ηd = 0.03–0.09) in swimming (e.g., Zamparo et al. 2011); the lowest values of ηd being reported in the studies where passive drag is assessed instead of active drag. Only few studies investigated so far overall (sometimes defined mechanical or gross) efficiency in swimming (e.g., Toussaint et al. 1990a, Zamparo et al. 2005a) and, also in this case, quite different values were reported in the literature (ηO = 0.10–0.20), due to the different methods proposed to measure/estimate total mechanical power. The reader is referred to Zamparo et al. (2011) for some historical considerations about this point.
The “uncertainty” in the values of ηd and ηO does not depend on the denominator of Eqs. 12 and 13, since metabolic power can, nowadays, be measured with a good accuracy also in the aquatic environment (see Sect. “Metabolic power”); rather, it depends on the estimates of the nominator of these equations (e.g., mechanical power output). Thus, reports of a lower efficiency in aquatic, compared to land, locomotion can be attributable to an underestimation of mechanical power output (or of one of its components) rather than to inefficiency per se.
However, whereas the power to overcome drag should necessarily be measured in water (and this is not an easy task), total mechanical power can also be measured outside it, with the aid of proper ergometers. As recently pointed out by Cortesi et al. (2019), notwithstanding the difficulties of accurate reproduction of the swimming movements outside the water, laboratory based ergometry (swim benches and swimming ergometers) can be used to investigate the mechanical requirements of swimming. Zamparo and Swaine (2012) calculated ηO by using a whole-body swim ergometer and found it to be, in competitive swimmers, of 0.23 ± 0.01 (when considering the external work only). Similar values (0.21 ± 0.02) were obtained when investigating just the action of the arms (arm-ergometer, dry-land) in male master athletes (Zamparo et al. 2014). These values are similar to those found in cycling and in rowing where \(\dot{W}_{\text{ext}}\) is easily measurable with proper ergometers. Values of ηO of about 0.25 should indeed be expected for all cyclic forms of locomotion in which no recoil of elastic energy takes place and for which mechanical power output can be accurately assessed. Thus, the “rather low” values of overall (gross) swimming efficiency reported in less recent studies seem indeed to be attributable to an incomplete computation of all work components/energy losses.
Froude and Propelling efficiency (ηF and ηP)
A swimmer needs to produce more mechanical power than that needed to overcome (active) drag when he/she moves in water: Froude/propelling efficiency indicates the fraction of external/total mechanical power that can be utilized in water to produce useful thrust. Thus, to calculate Froude/propelling efficiency both the work to overcome drag and the external/total work must be computed. Again, this is not an easy task in the aquatic environment and quite different values were reported in the literature (0.20–0.80) due to the different methods utilized to measure/estimate \(\dot{W}_{\text{d}}\), \(\dot{W}_{\text{k}}\) and \(\dot{W}_{\text{int}}\). The reader is referred to Zamparo et al. (2011) for some historical considerations about this point.
In the last decade a different approach to determine Froude efficiency was proposed: it attempts to estimate ηF based on measures of speed (easier to determine in this environment), rather than on measures of power. The rationale is that while a swimmer moves forward at constant speed (v) the upper limbs move forward and backward with a (larger) velocity (u) relative to the body. The ratio v/u is proportional to the theoretical (or Froude) efficiency in animal locomotion (Alexander 1983) as well as in all fluid machines (pumps, turbines, propellers, water wheels, and paddle wheels) (e.g., Fox and McDonald 1992). The limbs velocity (u, the hand speed for the arm stroke) can be directly measured by means of underwater kinematic analysis (e.g., Figueiredo et al. 2011; Gonjo et al. 2018) or estimated, by modelling the upper arms motion as if it was a paddle wheel motion (e.g., Zamparo et al. 2005a). The values of ηF calculated with this method, even if sometimes referred to as values of propelling efficiency, are indeed values of Froude efficiency (of the arm stroke only). In the many studies published so far with this model, ηF was found to be at most of 0.40–0.45 (in male elite front crawl swimmers), as expected on theoretical grounds (Zamparo and Swaine 2012, see below), and not to differ from the values of ηF measured by means of kinematic analysis (e.g., Figueiredo et al. 2011). As shown by Zamparo (2006), when calculated with this model, ηF (front crawl) changes as a function of age (is as low as 0.20 in young swimmers and in elderly masters) and no major differences are observed between males and females at all ages: mechanical power output could be lower in females but they have, for the same technical skill, the same ability to transform it into useful power for propulsion. Froude efficiency decreases with speed as stroke length (SL) does (e.g., Peterson-Silveira et al. 2017) and indeed a significant correlation can be observed between these two parameters: larger values of ηF are associated to larger values of SL (e.g., Zamparo et al. 2005a, 2014; Toussaint and Hollander 1994).
Thus, were a swimmer, at a given speed, able to reduce his/her movement frequency (thus increasing his/her stroke length), his/her internal power would decrease (see Sect. “Internal and “wasted” power (\(\dot{W}_{\text{int}}\) and \(\dot{W}_{\text{k}}\))”), his/her Froude efficiency would increase and his/her energy cost would decrease (see Eq. 3).
A recent study of Gonjo et al. (2018) reports data of v/u ratio (assessed by 3D kinematic analysis) also in the backstroke, with lower values compared to front crawl (0.34 vs. 0.40 at a speed of about 1 m·s−1); as it can be expected in the framework of Eq. 3, these difference in efficiency are associated with differences in energy cost between these strokes. No data of Froude efficiency are reported in the literature for the butterfly and the breaststroke.
A further approach to investigate Froude efficiency was put forward by Zamparo and Swaine (2012): indeed, ηF can be calculated based on values of drag efficiency and overall (gross) efficiency (ηF = ηd /ηo). Based on measured values of ηO of about 0.23 (obtained by using a whole-body swimming ergometer in dry conditions) and since the values of ηd reported in the literature could be at most 0.09, Froude efficiency could be at most 0.39. This value is close to those determined with the methods described above (based on the v/u ratio) thus confirming that humans can transform less than half of total power output into power useful for propulsion when moving in water.
A recent proof in favour of these estimates, comes from a study of Gatta et al. (2018) who recently measured (external) mechanical power by means of a whole-body swimming ergometer in a group of sprinters and found these values larger than those of thrust power in the same swimmers. The ratio of useful power (thrust power, exerted in water) to external mechanical power (exerted outside the water, with the ergometer) is of 0.40, a value close to that obtained by means of the “paddle wheel method” (0.39) on the same swimmers (in both cases these are values of Froude efficiency).
Last but not least, the limbs speed (u in the v/u ratio) is that of the upper limbs only but also the lower limbs contribute to propulsion (to the value of v), thus, either the swimmers should be asked to use a pull buoy (and in this case propulsion is totally provided by the arms) or the effects of leg kick should be accounted for (e.g., Zamparo et al. 2005a). Not taking into account the contribution of the legs implies an overestimation of the arm stroke efficiency (Peterson-Silveira et al. 2017).
Factors influencing the energy cost of swimming
According to Eq. 3 the determinants of the energy cost of swimming are propelling efficiency, hydrodynamic resistance and overall efficiency. Whereas overall efficiency is seldom investigated in swimming studies, many factors are known to influence C through their effects on drag or propelling efficiency, or both (e.g., morphological characteristic could affect both drag and propelling efficiency). To single out the effects of one factor, the other “confounding factors” must be kept constant. As an example: (1) sex differences can be meaningfully evaluated only in swimmers with the same technical level because technical skill has a deep influence on C (e.g., Morris et al. 2016); (2) age differences in C should be evaluated at the very same speed, even if it could be difficult to make a senior master swim at the same speed of a younger one (e.g., Zamparo et al. 2012a); (3) differences among strokes should be evaluated in swimmers with similar anthropometric and physiological characteristics otherwise the eventual differences in C would reveal differences in hydrodynamic resistance and fitness level (e.g., Gonjo et al. 2018). Many of the “discrepancies” reported in the literature about the energy cost of swimming and its determinants could be referred to these (and similar) considerations.
Speed and stroke
The energy cost of swimming increases with speed and differs among strokes. After the seminal work of Holmer (1974), the most complete analysis of the effects of speed and stroke on the energy expenditure of swimming was performed by Capelli et al. (1998) who measured C in young elite male swimmers in the four strokes at submaximal and maximal speed. For this population the relationship between C and v is well described by the following equations: front crawl: C = 0.670v1.614, backstroke: C = 0.799v1.624, breaststroke: C = 1.275v0.878, butterfly: C = 0,784v1.809. Hence, for a given speed, C is lowest for the front crawl followed by the backstroke, the butterfly and the breaststroke being the most demanding strokes (e.g., Holmer 1974; Capelli et al. 1998). Similar conclusions were reported in more recent studies where only one or few speeds were investigated, mainly in the aerobic speed range (e.g., Barbosa et al. 2006; Gonjo et al. 2018) (see Fig. 5).

Energy cost of swimming in different populations of swimmers at a given, submaximal, speed (1 m·s-1). Differences in C, at a given v, can be observed among strokes (in swimmers of the same age, sex and technical skill), between males and females (of the same age and technical skill and for the same stroke) and as a function of age (in swimmers of the same sex and for the same stroke; in this latter case is more difficult to disentangle the effects of skill for those attributable to age per se). See text for details
For a given stroke, the increase in C with speed, is mainly attributable to the concurrent increase in water resistance while, at any given speed, the differences in C among strokes could be attributed to differences both in Fd and ηP (Eq. 3). As an example, as suggested by Gonjo et al. (2018) front crawl is less costly than backstroke (at a speed of about 1 m·s−1) because of differences in ηP (larger in front crawl) rather than in hydrodynamic resistance. Indeed, no major differences in frontal area (and hence in Fd) were observed between these two strokes (Gatta et al. 2015). Unfortunately, values of Fd (in active conditions) and ηP are not available for the other strokes.
Differences in C between strokes could also be attributed to the different role of the lower and upper limbs to propulsion; in the breaststroke the leg push (less efficient) plays a more important role than in the other strokes, where propulsion is essentially generated by the arm pull (more efficient).
Besides swimming stroke also swimming style/technique could influence C, but this topic is less investigated in the literature. As an example, the amount and timing of body roll in front crawl and backstroke can influence drag as well as propelling efficiency (Payton and Sanders 2011) and this is expected to influence C. Another example is the different styles in the breaststroke (e.g., flat or undulating): timing and degree of undulation influence Fd (or at least its determinants, such as frontal area) and probably also ηP; indeed, different breaststroke styles/techniques are characterized by differences in swimming economy (e.g., Vilas-Boas and Santos 1994). The duration of the gliding phase in the breaststroke was also observed to influence C, at controlled speed (e.g., Seifert et al. 2014b); in this case the differences in C were not attributed to differences in frontal area (and hence in Fd) but to differences in the intra-cyclic variations in speed (IVV) and in inter-limb coordination. The effects of these parameters on C are discussed in more detail in Sects. “Intra-cyclic velocity variations” and “Inter-limb coordination”, respectively.
Stroke parameters (length and frequency)
Swimming velocity is determined by stroke length and stroke frequency (v = SL · SF). An increase in swimming speed is generally obtained by an increase in SF while SL remains stable at low-moderate speeds and tends to decrease at fast speeds. The relationship between these three parameters is unique for each stroke (e.g., Craig and Pendergast 1979) and each swimmer selects his/her unique combination of SL and SF to reach and maintain the highest possible speed. Stroke parameters are related to the swimmer’s technical ability: faster swimmers, indeed, swim with a greater SL and with a slower rate (e.g., Craig and Pendergast, 1979; Craig et al. 1985). Several studies have shown that an increase in SF is associated with an increase in C in all strokes (e.g., Wakayoshi et al. 1995; Barbosa et al. 2005; Smith et al. 1988) even if in many of these studies the effects of SF on C were observed during incremental protocols and this actually reflects the effect of v on C. The effect of SF on C was, however, observed also when controlling for speed, in all strokes (e.g., Barbosa et al. 2008). This suggests that if a swimmer were able to reduce SF at a given speed, his/her C at that speed would decrease: the manipulation of stroke mechanics is thus a factor through which C can be altered.
The effect of SF on C could be attributed to two main mechanisms: (1) an increase in SF brings about an increase in \(\dot{W}_{\text{int}}\) (see Sect. “Internal and “wasted” power (\(\dot{W}_{\text{int}}\) and \(\dot{W}_{\text{k}}\))”); this, in turn, would increase \(\dot{W}_{\text{tot}}\) and metabolic power requirements; (2) an increase in SF, at a given speed, is necessarily associated with a decrease in SL (e.g., in propelling efficiency, see Sect. “Froude and Propelling efficiency (ηF and ηP)”) that would also increase energy expenditure (see Eq. 3).
As reviewed by Barbosa et al. (2010) a clear relationship between C and SL is not observed in all studies/strokes, probably because speed was not controlled (otherwise the changes in SF mirror the changes in SL). Moreover, at variance with SF, SL remains stable at slow to moderate speeds and thus its effects on C could be better appreciated at maximal speed. Indeed, in these conditions, the decrease in speed brought about by the development of local fatigue is always associated to a decrease in SL (e.g., Craig and Pendergast 1979; Craig et al. 1985) and a consequent increase in C (e.g., Figueiredo et al. 2011).
The most complete analysis of the effects of stroke mechanics on energy expenditure (in front crawl) was performed by Termin and Pendergast (2000) who investigated the effects of a 4 years “high-speed training” on the v-SF relationship and on energy expenditure (see Figs. 6 and 7). The 8–10% increase in performance over the 100–200 m distances (greater than the 3% improvement generally observed for more conventional, long distance training programs) was attained by a shift of the v-SF relationships up and to the right (+ 10% per year): at submaximal speeds the swimmers were able to sustain each speed with a lower SF (and thus with a larger SL); in addition, they were able to reach a faster vmax (+ 27%) also because they were capable to reach and sustain a higher SFmax (+ 8%). These authors have furthermore shown that these changes in stroke mechanics are “coincident” with a decrease in the energy cost of swimming over the entire range of speeds (− 20%) and to an increase in \(\dot{E}_{{\max} }\) (+ 16%) both in its aerobic (\(\dot{V}{\text{O}}_{2{\max} }\): + 48%) and anaerobic (\(\dot{E}_{\text{AnL}}\): + 35%) components, as it could be expected based on Eq. 2: the increase in vmax is, therefore, attributable to both by an increase in \(\dot{E}_{{\max} }\) and a decrease in C.

Changes in the v vs. SF relationship in front crawl before and after four years of a “high-speed” training program (adapted from Termin and Pendergast 2000): after training the swimmers were able to swim (at a given submaximal speed) with a lower SF (and thus with a larger SL) and to reach a higher vmax because capable to sustain a higher SFmax.

The changes in the stroke parameters reported in Fig. 6 after four years of a “high-speed” training program are associated with changes in energy expenditure (adapted from Termin and Pendergast 2000): after training the swimmers were able to swim (at a given submaximal speed) with a lower energy expenditure (\(\dot{E}\) is the metabolic power) (and thus with a lower energy cost: C = \(\dot{E}\)/v) and to reach a higher vmax because they increased their \(\dot{E}_{{\max} }\) (vmax = \(\dot{E}_{{\max} }\) /C). See text for details
Training, detraining, and skill level
The American College of Sports Medicine guidelines (ACSM 2018) underline that energy expenditure “can vary substantially from individual to individual during swimming as a result of different strokes and skill levels”. Indeed, technical skill in swimming deeply influences C because of its effects on ηP (and, to a lower extent, also on Fd). As indicated by Holmer (1972), for any given speed in the aerobic range, a recreational swimmer can spend as much as twice the energy that a good swimmer uses for proceeding in water at the same speed and the latter consumes about 20–30% more energy than an elite swimmer at comparable speeds (see Fig. 2). Similar results were reported by Chatard et al. (1990c) who investigated swimmers of three performance levels: at a given speed (e.g., 1.1 m·s−1) the more proficient swimmers expend 50% less energy than less proficient ones and 25% less than the intermediate group.
The volume of aerobic or anaerobic training and specialization in long distance or sprint events have an impact on energy expenditure: at submaximal speeds sprinters generally have a higher C than long distance swimmers (e.g., Seifert et al. 2010b; Chatard et al. 1990c). This reveals the importance of the testing protocol: long distance swimmers are expected to be less proficient when asked to perform maximal sprints whereas sprinters are at disadvantage when performing aerobic tasks. This could explain why, in some studies, differences in C between competitive swimmers grouped for swimming speed (high or low) tend to disappear (Ribeiro et al. 2017); in this latter study a maximal 100 m sprint was utilized and this is expected to put “slow” swimmers at disadvantage (raising their C) and to favour high-speed sprinters. Indeed, Ribeiro et al. (2017) reported lower values of propelling efficiency, over this distance, in long distance swimmers compared to sprinters; lower values of ηP, according to Eq. 3, are associated with larger values of C. These considerations highlight the difficulty of singling out the factors that affect swimming performance.
Training is expected to improve technical skills and to lead to a significant decrease in C through an increase in ηP; on the other hand detraining is expected to lead to a decrease in ηP and an increase in C. A significant effect of training on the energy cost of swimming was observed after a 4 years “high-speed training program” (e.g., Termin and Pendergast 2000, see Figs. 6 and 7); this was not observed in other studies (e.g., Costa et al. 2012; Clemente-Suarez et al. 2017) probably because of different training stimuli (intensity and volume) and a much lower training duration.
Accordingly, the effects of detraining are expected to depend on the duration of the training cessation: short (typically the off-season) or long-term detraining (e.g., Mujika and Padilla, 2000). Caution should be taken when investigating the effects of training/detraining in young swimmer since changes in C and ηP could also derive from growth and maturation (see Sect. “Age and sex”): as an example, the “expected” increase in C after detraining could be offset by improved stroke mechanics, such as a higher SL because of an increase in arm span (e.g., Moreira et al. 2014; Zocca et al. 2017) that are generally associated to a decrease in C (see Sect. “Age and sex”).
Differences in C after training/detraining can also derive from differences in Fd (in active conditions). Data of active drag before and after 4 years of a “ high-speed training protocol” (i.e., collected during the study of Termin and Pendergast, 2000) are reported by Pendergast et al. (2005): active drag decreased of 15–35% and, according to Eq. 3, this reduction contributes to the reduction in C observed after the training period.
Intra-cyclic velocity variations
Swimmers do not move at constant speed (see Sect. “Energy cost”) and the speed profile shows intra-cyclic variations (IVV) that differ according to the swimming stroke: larger in breaststroke and butterfly than in freestyle and backstroke (e.g., Craig and Pendergast, 1979). Large velocity fluctuations increase the work needed to proceed at a certain velocity, both for the need to overcome inertia and drag. A “smooth motor pattern”, with reduced IVV, is thus expected to minimize C. Indeed the differences in C among strokes can be (at least partially) attributed to their differences in IVV and the effects of IVV on C are more easily observed in breaststroke and butterfly (e.g., Vilas-Boas, 1996; Barbosa et al. 2005) than in front crawl and backstroke (e.g., Figueiredo et al. 2013; Gonjo et al. 2018) where the IVV are much lower. In some studies on this topic the effects of IVV on C were, however, “overestimated” because the relationship between C and IVV was investigated by pooling data collected over a wide range of speeds (e.g., Barbosa et al. 2005) and thus rather reflects the effects of V on C. The discrepancies in the findings of the studies in this area can be attributed, among the others, to the methods used to measure IVV and their limitations. In a study of Psycharakis et al. (2010) the IVV were accurately measured (in 3D and relative to the body centre of mass) during a 200 max front crawl swim and showed a noteworthy consistency across laps (regardless of the decrease in speed due to fatigue): IVV are thus not correlated to swimming performance (to average speed). Similar findings were observed by Figueiredo et al. (2012, 2013) who, in addition, observed that C increased from the first to the last lap of a 200 m max trial while ηP decreased: thus IVV are not associated to changes in C or ηP either and cannot be considered an index of swimming efficiency/economy (at least in front crawl). Figueiredo et al. (2013) observed that IVV remain stable along the effort because the swimmers adapt their pattern of coordination during the race to cope with fatigue; they indeed found significant correlations between the index of coordination (IdC) and C or ηP (see Sect. “Inter-limb coordination”).
Arms, legs and whole stroke
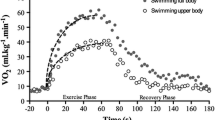
A comprehensive analysis of the energetics of swimming with arms only (AO), legs only (LO), and with the whole stroke (WS) was performed by Holmer (1974) in the four strokes at speeds up to 1.2–1.4 m·s−1 (in a swimming flume). He reported that, at any given speed, energy expenditure (\(\dot{V}{\text{O}}_{2}\)) is lower in AO (with pull buoy) compared to WS and is the largest in LO (with kickboard) in all strokes but the breaststroke (where no major differences in \(\dot{V}{\text{O}}_{2}\) are observed among conditions). Holmer (1974) also reported that, in all strokes, larger values of \(\dot{V}{\text{O}}_{2{\max} }\) (and vmax) could be attained in WS compared to LO and AO; he furthermore investigated the differences in water resistance (during passive towing) when using a pull buoy or a kickboard.
More recent literature confirms and extends these findings focusing, mainly, on front crawl swimming at maximal speed. The leg kick is, per se, very energy demanding (e.g., Holmer 1974; Zamparo et al. 2002) but its contribution to the whole stroke allows to increase vmax in front crawl of about 10% (e.g., Deschodt et al. 1999; Morris et al. 2016; Gourgoulis et al. 2014; Ribeiro et al. 2015); at submaximal swimming speeds this contribution tends to decrease (e.g., Peterson-Silveira et al. 2017). At sprinting speed, energy expenditure is larger in WS than in AO not only because of the larger muscle mass involved but also because of the differences in vmax; both factors are indeed responsible for the observed differences in the energy demands and in the energy system contribution in these two conditions (e.g., Ogita et al. 1996; Ribeiro et al. 2015).
The lower limbs action is relevant for trunk balance, buoyancy, and overall coordination (e.g., Gatta et al. 2012) but the leg kick not only helps to keep the body in a more streamlined position reducing A (and hence Fd) (e.g., Gourgoulis et al. 2014) but also has an influence on the propulsive action of the arms modifying the trajectory of the hand (Gourgoulis et al. 2014; Deschodt et al. 1999) and increasing stroke length (and hence improving ηP) (Zamparo et al. 2005a; Peterson-Silveira et al. 2017; Morris et al. 2016). As underlined several times in this review, speed must be controlled when investigating the effects of the leg kick and arm stroke on the whole stroke, otherwise the observed differences in energy expenditure/energy cost could be (also) attributed to differences in speed. On this line of reasoning Morris et al. (2016, 2017) compared AO, LO, and WS at controlled kick and stroke rates (since these parameters are known to affect C). Interestingly Morris et al. (2016) observed that, at paired and moderate speeds, C in AO was larger than in WS in males than in females probably because they have to increase more their core activation (in the arms only effort) to maintain a hydrodynamic body position (even when using a pull buoy) (see Sect. “Age and sex”).
The effects of the leg kick on the kinematics of the arm stroke indicate that inter-limb coordination (e.g., synchronization of arm and leg movements) should be optimized to decrease C and improve swimming performance (see Sect. “Inter-limb coordination”).
Inter-limb coordination
Swimming propulsion is based on the coordinate action of the upper and lower limbs. When swimming speed increases the swimmers change their arms (and legs) coordination to adapt to the higher water resistance encountered. The index of coordination (IdC) reflects changes in the organization of the stroking phases and measures the time gap between the pull and push phases of the upper arms in front crawl (e.g., Chollet et al. 2000). Other indexes were proposed to measure the time gap between the upper and lower limbs movements in the breaststroke (e.g., Seifert et al. 2014a). The IdC increases (becomes less negative) with speed and, for a given speed, is higher in males than in females of comparable technical skill (Seifert et al. 2004), is higher in more skilled swimmers (e.g., Seifert et al. 2010a) and in sprinters compared to long distance swimmers (e.g., Seifert et al. 2010b) . For these reasons, differences in IdC among swimmers could be expected to correlate with differences in C. However, in several studies on this topic the relationship between IdC and C was investigated by pooling data collected over a wide range of speeds, actually reflecting the effects of v on C (e.g., Komar et al. 2012; Seifert et al. 2010b); according to these studies a high IdC is associated with an elevated energy cost (which is indeed paradoxical). As shown by Fernandes et al. (2010) when “removing the effect of velocity” the relationship between IdC and C becomes, indeed, non-significant.
Seifert et al. (2014a, b) investigated whether the coordination pattern (at a given, submaximal, speed of about 1 m·s−1) can be adapted /constrained (e.g., by asking the swimmers to increase or decrease their gliding phase duration in front crawl and backstroke) and whether these adaptations are associated to changes in the metabolic demands. They observed that, in front crawl, the freely chosen pattern is the most economical one (probably because more trained) and that a high IdC is associated with a low C (which seems reasonable, but contradicts previous results, see above) (e.g., Seifert et al. 2014a). Expert breaststroke swimmers, instead, select the coordination pattern that involves the minimal glide duration, which is characterized by the lower IVV and lower C (Seifert et al. 2014b). This suggests that, also in swimming, movement is adapted with respect to energy cost minimization, as in terrestrial locomotion (e.g., Sparrow and Newell 1998).
At maximal speeds (e.g., a 200 m max front crawl swim) the decrease in speed from lap 1 to 4 is associated to an increase in IdC, a decrease in ηP and an increase in C (with no changes in IVV); in these conditions a higher IdC is thus associated with a high C (e.g., Figuereido et al. 2013). The interplay between all these parameters is therefore complicated by fatigue evolvement and this, again, highlights the difficulty of singling out the factors that affect swimming performance.
Age and sex
Both ηP and Fd depend on the anthropometric characteristics of the swimmer and both change during growth (along with body development and training). Therefore, C is expected to differ between children and adults and between males and females (see Fig. 5). C is about 20–30% lower in females than in males, at submaximal swimming speeds (e.g., Chatard et al. 1991; Morris et al. 2016; Pendergast et al. 1977; Zamparo et al. 2000, 2008b), the higher economy of female swimmers is traditionally attributed to a smaller hydrodynamic resistance (Fd) due to their smaller size, larger percentage of body fat and more horizontal position in water (lower A) compared to male swimmers (e.g., Zamparo et al. 1996; Zamparo et al. 2008b; Pendergast et al. 1977; di Prampero 1986). Since no major differences in the anthropometric characteristics are observed between males and females before puberty, no differences in C are observed between boys and girls (of similar technical skills) in childhood as well (e.g., Zamparo et al. 2008b; Ratel and Poujade 2009).
Only few studies have investigated, so far, the determinants of swimming economy in children and adolescents. These studies indicate that, at comparable speed, C is indeed lower in children than in adults (e.g., Kjendlie et al. 2004a, b; Poujade et al. 2002; Ratel and Poujade 2009), the more so the younger the age (Zamparo et al. 2008b). The higher economy of children is attributed to a smaller hydrodynamic resistance due to their smaller size and more horizontal position in water (e.g., Zamparo et al. 2008b; Kjendlie et al. 2004a). The increase in C with growth could, at least in part, be attributed to an increase in (pressure) drag; however, the increase in stature during growth also results in a better streamline of the body (e.g., length/thickness ratio) and in an increase in Froude number (e.g., a decrease in wave making resistance, see Sect. “Hydrodynamic resistance (Fd) and power to overcome drag (\(\dot{W}_{\text{d}}\) )” (e.g., Toussaint et al. 1990b; Kjendlie and Stallman 2008): the effects of age on drag (and on C) are thus part of a quite complex process that takes place during growth. As reported in Sect. “Hydrodynamic resistance (Fd) and power to overcome drag (\(\dot{W}_{\text{d}}\) )”, when C is scaled for body size (e.g., length or surface area) the age differences tend to disappear (e.g., Ratel and Poujade 2009; Kjendlie et al. 2004a, b;).
When normalized for their anthropometric characteristics, the differences in C between males and females tend also to disappear; this implicitly suggests that no major differences in propelling efficiency are to be expected between male and female swimmers of the same age and technical skill, as indeed found in some studies (e.g., Zamparo 2006; Zamparo et al. 2008b). Sex differences can be, however, observed in SL (an index of propelling efficiency) that is lower, at a given speed, in females compared to males of the same technical level (e.g., Zamparo 2006; Seifert et al. 2007) and age (e.g., Zamparo et al. 2008b). Young (pre-pubertal) swimmers are instead characterized by lower ηP values compared to adults, the more so the younger the age (Zamparo, 2006; Zamparo et al. 2008b) but this, more than to age per se, can be attributed to differences in technical skill. Indeed, in some studies, it was observed that when C is normalized to body dimension (e.g., stature or BSA) children are less economical than adults (e.g., Poujade et al. 2002; Kjendlie et al. 2004b) and this can be attributed to differences in technical skill (and hence in ηP).
Taken together, the values of (absolute) energy cost reported in the literature indicate that: (1) C, at a given speed, is lower the lower the age (see Fig. 5); (2) the increase in C with age is steeper in boys than in girls; so that (3) C is larger in males than in females (see Fig. 5). In both sexes, maximal aerobic and anaerobic power (the determinants of \(\dot{E}_{{\max} }\), see Eq. 2) increase with age and training, contributing to the improvement in performance and possibly masking the effects of changes in ηP and Fd (and therefore of C) on vmax.
Master swimmers are characterized by a lower economy (a higher C) compared to young swimmers, the more so the older the age (Zamparo et al. 2012a). The decrease in maximal metabolic power that occurs with age forces these swimmers to reduce their speed (Donato et al. 2003; Tanaka and Seals 1997; Zamparo et al. 2012b). At very low speeds (< 0.8–0.9 m·s−1) this is associated with an unfavourable alignment of the body (an increase in A that reduces the decrease in Fd that can be expected based on the decrease in speed alone); more importantly, though, with age a deterioration in ηP occurs bringing about an increase in C (of about 0.75% per year after the age of 50) (Zamparo et al. 2012a). The differences in ηP and C observed in the study of Zamparo et al. (2012a) could be partially attributed to differences in v (the eldest swimmers were not able to sustain the target speed of 1 m·s−1) and to differences in technical skill between swimmers of different ages. In a more recent study of Mejias et al. (2014) no differences in C and ηP were observed between young masters (formerly elite swimmers; 30 years of age) and competitive swimmers (20 years old) but the experiments were performed at relative and not absolute speeds (a 200 m maximal test); since the masters were swimming at slower speed but with the same C of the elite swimmers this suggests that they would be less economical if tested at comparable speed. Further studies should thus be performed to understand the complex relationship between age per se, technical skill and detraining in this population. In any case, as indicated by Zamparo et al. (2012b), no major changes in C should be expected before 50 years of age since, up to this age, the decrease in \(\dot{E}_{{\max} }\) fully explains the decrease in swimming performance (see Eq. 2).
When comparing swimmers with different technical level (e.g., children, adults and master swimmers) a given absolute speed could correspond to different relative intensities: children (and old master swimmers) are expected to swim at a higher percentage of their aerobic maximal velocities compared to competitive swimmers so that, in some studies, it was preferred (reasonably) to compare different populations at the same relative, instead of at the same absolute, velocity (e.g Ratel and Poujade; 2009, Zocca et al. 2017). However, since C changes with speed, differences in C at “relative speeds” could reveal the effect of v on C. Again, these considerations highlight the difficulty of singling out the factors that affect swimming performance in different populations of swimmers.
“Reasonable” assumptions
As indicated in the previous sections to measure ηP, ηO and Fd is of paramount importance to understand the source of variability in the energy cost of swimming but, unfortunately, this is not an easy task and indeed the different methods proposed so far in the literature to assess/calculate these parameters yield very different results. The reader is referred to Zamparo et al. (2011) and Zamparo and Swaine (2012) for a detailed discussion on this point.
As reported in the previous sections, the most recent evidence (in front crawl swimming) indicates that:
-
active drag is larger than passive drag (Fd active ~ 1.5 Fd passive),
-
Froude efficiency is of about 0.4,
-
overall efficiency is of about 0.2.
If these values are “reasonable”, it should be possible to estimate C with fairly good accuracy. Indeed, based on data of Fd (active drag) and ηF at a given speed external mechanical power can be calculated (\(\dot{W}_{\text{ext}} = F_{\text{d}} v/\eta_{\text{F}}\)); based on data of \(\dot{W}_{\text{ext}}\) and ηO it is thus possible to calculate \(\dot{E}\) (= \(\dot{W}_{\text{ext}} /\eta_{\text{O}}\)) and hence C (= \(\dot{E}/v\)). These estimates could then be compared to the values of C reported in the literature, at that speed.
The C vs. v relationship in elite (male) collegiate swimmers reported by Capelli et al. (1998) is indicated by the continuous line in Fig. 8 and the white circle indicates the C value measured at maximal speed (1.75 m·s−1) in these swimmers, over the 100 m distance.

Measured and estimated values of C. Continuous line: the C vs. v relationship in elite (male) collegiate swimmers (as reported by Capelli et al. 1998); white circle: C in elite (male) sprinters (covering a 100 m distance at a speed of 1.75 m·s-1; Capelli et al. 1998); black circle: C in elite (male) open water swimmers (covering a 2 km distance at a speed of 1.43 m·s-1; Zamparo et al. 2005a). White and black squares represent estimates of C based on different values of Froude efficiency, overall efficiency and active drag. See text for details
Let’s take a “typical value” of passive drag for a competitive male swimmer (e.g., Fd passive = 25v2) and assume that Fd active ~ 1.5 Fd passive (therefore Fd active = 37.5v2) let’s further assume that ηP = 0.4 and ηO = 0.2 and let’s use Eq. 3 to estimate the energy cost at this speed: C = (37.5 · 1.752) · (0.40 · 0.20) = 1.44 kJ·m−1. This value is not far from that reported by Capelli and coworkers (white circle: 1.73 kJ·m−1) and it is indeed expected to be somewhat lower if only because the internal work is not taken into account (these calculations are based on \(\dot{W}_{\text{ext}}\) instead of \(\dot{W}_{\text{tot}}\)) and because active drag is probably larger than that assumed here (see Gatta et al. 2015). The important point, however, is that the values of ηP, ηO, and Fd utilized in these calculations are indeed quite “reasonable”. Indeed, when these calculations are repeated by considering passive drag instead of active drag, the value of C drops to 0.95 kJ·m−1; if, in addition, Froude efficiency is considered to be as large as 0.70 (as reported in some studies), C further drops to 0.55 kJ·m−1 (see white squares in Fig. 8).To “correct” for these “un-correct” assumptions (to regain a value of C close to that reported by Capelli et al. 1998) overall efficiency should be of about 0.015 and this seems, indeed, “not reasonable”.
Another example can be based on data of C reported by Zamparo et al. (2005b) in elite (male) open water swimmers while covering a 2 km distance at a speed of 1.43 m·s−1. Considering the same assumptions as above: C = (37.5 · 1.432) · (0.40 · 0.20) = 0.96 kJ·m−1. This value is not far from that reported in the study of Zamparo and coworkers (1.19 kJ·m−1) and measured based on aerobic energy sources only (therefore no “assumptions” were needed in that study to calculate/estimate the anaerobic energy sources, which are indeed negligible in these conditions). Again, beyond the consideration that the calculated value is expected to be somewhat lower than the measured one, by considering passive drag instead of active drag, the calculated value of C drops to 0.64 kJ·m−1; if, in addition, Froude efficiency is considered to be as large as 0.70, C further drops to 0.37 kJ·m−1 (see black squares in Fig. 8).
These simple calculations show that our knowledge of the determinants of C in swimming is, nowadays, “quite reasonable”, at least for the front crawl. Of course, more research is needed to refine the estimates of Fd active, ηP and ηO. The example at maximal speed (where also anaerobic energy sources are utilized) indicates, albeit indirectly, that also our estimates of metabolic power input (the other parameter that determines performance in swimming, E′max) are indeed “fairly well known”, nowadays.
Open water swimming and ultra-endurance races
Open water swimming (OWS) is an outdoor endurance discipline that takes place in oceans, seas, rivers and lakes. Conventional races are covered over the 5, 10 and 25 km distances (the 10 km race is an Olympic event since 2008) whereas in ultra-endurance races the swim course is typically longer than 30 km. The three most challenging open water swims worldwide are the ‘English Channel Swim’ (34 km), the ‘Catalina Channel Swim’ (32.2 km) and the ‘Manhattan Island Marathon Swim’ (45.9 km) also called the ‘Triple Crown of Open-Water Swimming’.
For these disciplines Eq. 2 can be written as:
where F is the fraction of \(\dot{V}{\text{O}}_{2{\max} }\) (maximal oxygen uptake) that can be sustained during the race, and vend is the endurance speed (di Prampero 1986).
As reviewed by Baldassarre et al. (2017) the \(\dot{V}{\text{O}}_{2{\max} }\) values of elite middle-distance and open water swimmers are comparable to those reported for athletes specialized in endurance events on land (e.g., marathon runners): about 60 and 70 ml·min−1.kg−1 for female and male swimmers, respectively (e.g., Dwyer 1983). However, rather than a high \(\dot{V}{\text{O}}_{2{\max} }\) per se, the ability to sustain an elevated fraction of \(\dot{V}{\text{O}}_{2{\max} }\) is the factor associated to a good performance in endurance events (e.g., di Prampero 1986). Even if no specific studies on these matters were performed in swimming, in land locomotion (e.g., running) it was observed that, in well trained subjects and for races shorter than 8–10 min, F is close to 1 and that F decreases steadily with race duration (e.g., to 0.77, 0.73, 0.69 and 0.65 after 1, 2, 4 and 8 h, respectively, as reported by Peronnet and Thibault 1989). As shown by Tam et al. (2012) in marathon runners, \({\text{F}}\dot{V}{\text{O}}_{2{\max} }\) corresponds to the ratio of the speed attained at the lactate threshold to vend.
It seems thus logical to hypothesize that being able to swim at a high fraction of \(\dot{V}O_{2\max}\) is important for all open water races as well as in pool events over the longest distances (e.g., 1500 m). Equation 16 indeed indicates that larger values of vend would be attained by swimmers with larger values of \({\text{F}}\dot{V}{\text{O}}_{2{\max} }\) and/or with higher speeds at lactate threshold. Indeed, in elite open water swimmers, VanHeest et al. (2004) reported that lactate threshold occurred at a speed equal to 89% of peak pace for males and 94% for females. Other estimates (e.g., Zamparo et al. 2005b) are close to these values: 91% and 97% for males and females during a 10 km OW competition.
Direct measurements of energy expenditure during open water swimming races are technically difficult and the few studies reported in the literature focused mainly on cardiovascular responses (heart rate monitoring) and stroke mechanics. Stroke frequency (SF) and Stroke length (SL) of a master swimmer while solo crossing the Adriatic sea (78.1 km in 23:44 h:min) were monitored by De Ioannon et al. (2015): after the first 3 h, SL and swimming speed started to decrease and SF to increase; besides the possible effects of environmental conditions (tides, current and waves) on stroke mechanics this suggests a detrimental effect of fatigue on swimming technique. It has indeed been shown that, in fatiguing conditions, SF tends to increase and SL to decrease (e.g., Zamparo et al. 2005b; Figueiredo et al. 2011; Toussaint et al. 2006) and since SL is an index of propelling efficiency (see Sect. “Froude and Propelling efficiency (ηF and ηP)”) a decrease in SL is expected to bring about an increase in C. This was indeed observed by Zamparo et al. (2005b) who measured C in elite endurance swimmers (during 400 m swims in the pool) before and after a “fatiguing” 2 km trial performed at 10 km race pace. The authors noted an increase in C (about 20%) and SR after the fatiguing trial and a decrease in SL, suggesting a decrease in propelling efficiency in fatigued conditions.
Endurance swimmers are characterized by values of C comparable to those of short distance swimmers (matched for sex, competitive level and age) and females have lower values of C compared to males at the same swimming speed (Zamparo et al. 2005b). As indicated in Sect. “Hydrodynamic resistance (Fd) and power to overcome drag (\(\dot{W}_{\text{d}}\))”, the smaller body size and greater fat percentage in females results in less drag and lower energy cost; thus, according to Eq. 16, and considering the larger values of \({\text{F}}\dot{V}{\text{O}}_{2{\max} }\) in females this suggests that they are particularly “suited” for excelling in long distance events.
FINA does not hold official world records for open water swimming competitions since weather, water conditions, temperature and tides can considerably affect race time, irrespective of race distance (see Fig. 9). Contrary to other endurance races (i.e., marathon) where athletes race for best performances, OWS races are tactical events: successful swimmers adopt a “conservative tactic” during the race and increase their speed only in the last lap; the lead group is indeed unified until the last buoy to benefit from drafting (see Sect. “Hydrodynamic resistance (Fd) and power to overcome drag (\(\dot{W}_{\text{d}}\))" (Rodriguez and Veiga 2018, Baldassarre et al. 2017, 2019) (Fig. 10).

Race speed as a function of race distance: the longer the race the lower the speed but for the Manhattan Island race (open circle) where the current of the Hudson river and the tides favour the swimmers. Weather, water conditions, temperature and tides can considerably affect race time in OWS, and thus evaluating performance in terms of swimming speed can be misleading, in these races. Open squares: Olympic distances (5, 10 and 25 km); full circles: Zurich lake, English channel and Catalina channel marathons. Redrawn from Zamparo et al. (2019)

Males/Females ratio in average swimming speed in conventional (Olympic distances of 5, 10 and 25 km) and ultra-endurance OWS events. Redrawn from Zamparo et al. (2019). In ultra-endurance swimming events, women could swim faster than men; they are indeed suited for long distance races because their smaller body size and greater fat percentage results in less drag and lower energy cost compared to males. See text for details
Even if evaluating OWS performance in terms of swimming speed can be misleading, sex differences in performance (for a given course) can indeed be compared and are of interest. The sex difference in swimming speed in elite swimmers competing in conventional 5, 10 and 25 km races is smaller (~ 7%, e.g., Vogt et al. 2013) compared to other ultra-distance disciplines such as running (up to 27%, e.g., Zingg et al. 2014). Sex differences are further reduced in ultra-endurance swimming events, where women could swim faster than men (see Fig. 9). In the 32 km ‘Catalina Channel Swim’ and the 46 km ‘Manhattan Island Marathon Swim’, the fastest women ever were faster than best-performing men (Sandbakk et al. 2018). The fastest ever to complete the Triple Crown was, indeed, a woman performing the 3 races within 36 days in 70:00 h:min (Knechtle et al. 2014).
Locomotory tools and boat locomotion
Changes in the energy cost of swimming can be observed when passive locomotory tools, such as fins and hand paddles, are used for moving in water. In analogy with locomotion on land these tools do not supply any additional mechanical energy to the body but provide effective compensation for limitations in the anatomical design and for inadequacy in muscle performance (Minetti 2004).
Fins, as an example, reduce the energy cost of front crawl swimming, at a given speed; of about 10–15% and this reduction is associated to a reduction in kick (and stroke) frequency and to an increase in ηF and ηP (Zamparo et al. 2005a). Energy expenditure, at a given speed, is also reduced when using hand paddles and, also in this case, this is associated to an increase in ηF (Ogita and Tabata 1993; Ogita et al. 1999; Toussaint et al. 1991). An increase in ηF and a decrease in stroke frequency when using hand paddles was also observed in other studies (e.g., Gourgoulis et al. 2006; Tsunokawa et al. 2019), the entity of these changes being proportional to the paddle surface, as in the case of fins. Thus, when small locomotory tools are used, the gain in speed/economy mainly derives from an increase in efficiency rather than a decrease in resistive forces, the power to overcome drag (when measured) being essentially unaffected by their use (e.g., Zamparo et al. 2005a; Tsunokawa et al. 2019).
Fins and hand paddles improve ηF and ηP essentially because they increase the “propelling surface area”; for a given propulsive force this means giving a larger amount of water a smaller change in velocity, thus reducing the kinetic energy dissipation (the term \(\dot{W}_{\text{k}}\), see Sect. “Internal and “wasted” power (\(\dot{W}_{\text{int}}\) and \(\dot{W}_{\text{k}}\))”). Accordingly, hands and foot size are also expected to affect swimming performance (Kjendlie and Stallman 2011) and energy cost. A small finger spread increases the “functionally equivalent” palm area possibly allowing the swimmers to enhance, albeit to a small extent, swimming propulsion (e.g., Minetti et al. 2009; van Houwelingen et al. 2017).
Far larger improvements in speed/economy can be observed when “larger tools” (e.g., human powered boats) are utilized. In this case, the lower energy requirement compared to swimming at a given speed, is not only attributable to an improvement in efficiency but also to a substantial decrease in hydrodynamic resistance (Pendergast et al. 2003).
As is the case for swimming, the energy cost of boat locomotion increases as a function of speed and depends on the type of boat (and on the skill level of the athlete). For kayaking with a flat-water sprint K1 scull C = 0.020v2.26 (Zamparo et al. 1999) and with a slalom canoe C = 0.098v1.76 (Pendergast et al. 1989), for rowing (two-oared shell with coxswain, for one rower) C = 0.088v1.05 (di Prampero et al. 1971), for sculling a venetian gondola C = 0.155. v1.67 (Capelli et al. 1990), for pedalling with a water bike (a double hulled catamaran) C = 0.063v1.57 (Zamparo et al. 2008a) or with a paddle wheel boat (used on lakes and beaches for recreational purposes) C = 0.179v2.66 (Zamparo et al. 2008a).
The values of C at a speed of 2 m·s−1, are reported in Fig. 11 for all these boats: the more economical mean of transportation is the K1 scull, followed by the catamaran and the rowing boat; the larger values of C being observed in the less hydrodynamic crafts (e.g., gondola and paddle wheel boat).

Energy cost of front crawl swimming and boat locomotion at a speed of 2 m·s-1: the more economical mean of transportation is the K1 scull (kayaking), followed by the catamaran (water bike) and the rowing shell; the less economical crafts are the less hydrodynamic ones (e.g., gondola and paddle wheel boat). It is very likely that data of C in in rowing would be much lower if measured nowadays because of the improvements in boat design since the experiments performed by di Prampero et al. (1971). Swimming data (front crawl, elite male swimmers) are taken from Capelli et al. (1998), boat locomotion data are taken from Zamparo et al. (1999), Pendergast et al. (1989), di Prampero et al. (1971), Capelli et al. (1990) and Zamparo et al. (2008a)
In analogy with swimming studies, in kayaking C was found to be up to 30–50% lower (at paired speeds) in elite than in less experienced paddlers (Pendergast et al. 1989; Buglione et al. 2011). These differences were attributed to differences in propelling efficiency. As is the case for swimming, for the same skill level, male kayakers were found to be less economical than females (Pendergast et al. 1989): a 5–12% difference in C, increasing with increasing speed. In this case the difference was attributed to differences in Wd (a lower body mass in female kayakers and hence a lower wetted area of the boat). Moreover, as previously indicated for swimming (Sect. “Training, detraining and skill level”), training decreases the energy cost of boat locomotion (of about 25–33% in rowing and 6–10% in kayaking) due to both an increase in propelling efficiency and a decrease in drag (e.g., after 4 years of training active drag can be reduced of 18–50% in kayaking; Pendergast et al. 2005).
As reviewed by Pendergast et al. (2003), hydrodynamic resistance, in boat locomotion, is largely reduced compared to swimming and the differences in drag among boats roughly mirror those in C. The most economical boats are long and narrow and are made of new generation of materials (e.g., carbon-fibre reinforced plastic) that decrease ”dead weight” (as is the case for the newest bicycles on land); the shape of the fastest boats is now believed to have reached a nearly optimal design (Greidanus et al. 2012). Compared to swimming, drag is thus reduced because wetted area, pressure drag and wave drag are reduced. At variance with swimming, however, in rowing, sailing or canoeing pressure and wave drag contribute only for 10–20% to total drag and friction drag is the major source of resistance to motion (80–90%) at the typical flow conditions for these sports (Greidanus et al. 2012). Therefore, friction drag reduction is a main issue in boat locomotion and boat wetted surfaces are generally treated to reduce turbulent flow (in a similar way that swimmers shave or wear suits) (see Sect. “Hydrodynamic resistance (Fd) and power to overcome drag (\(\dot{W}_{\text{d}}\)) ”).
Pressure drag is lower in boat locomotion (e.g., rowing and kayaking) compared to human swimming because boats are narrow whereas wave drag is low because long boats create less wave drag. Hull speed is the speed where the wavelength along the hull equals the hull length, above hull speed the cost of propulsion increases sharply. Since hull speeds is positively correlated with the boat length, the fastest boats are the longest ones (from 8 to 17 m in rowing), as the fastest swimmers (front crawl sprinters) are, generally, the tallest ones (e.g., Kjendlie and Stallman 2011) (see Sect. “Hydrodynamic resistance (Fd) and power to overcome drag (\(\dot{W}_{\text{d}}\)”).
In boat locomotion also oars’ length matters, as is the case for the upper limb length in swimmers, another anthropometric characteristic that influences swimming performance (e.g., Kjendlie and Stallman 2011). As indicated by Pendergast et al. (2003), the distance covered by the hull for each cycle (a parameter that corresponds to the “distance covered per stroke”, or stroke length, in swimming) is larger the longer the propulsive tool (oars/blades): about 8 m in rowing and 4 m in kayaking (it is at most 2–2.5 m in swimming) and is proportional to propelling efficiency which, in boat locomotion, ranges from 0.4 (for paddle wheel boats, a value similar to that of front crawl swimming) to 0.70 (in rowing and kayaking) (e.g., Pendergast et al. 2003). In the framework of Eq. 3, therefore, boats allow to decrease the energy cost of locomotion in water, compared to swimming, both because a decrease in hydrodynamic resistance and because an increase in propelling efficiency.
Thus, the knowledge of the parameters that affect performance in boat locomotion could be utilized to understand swimming performance (and viceversa) and human locomotion in water can be described by similar rules in spite of differences in the type of propeller (e.g., legs, arms), in the tools utilized (e.g., fins, paddles, boats) and in the pattern of movement (e.g., synchronous in rowing and butterfly, alternate in kayaking and front crawl).
Summary and conclusions
Propelling efficiency and hydrodynamic resistance are the major determinants of the energy cost of swimming (of aquatic locomotion, in more general terms); these parameters are difficult to measure in the water environment but recent evidence suggests that the estimates of ηP and Fd are nowadays “quite reasonable” thus allowing to estimate C with good accuracy, at least in front crawl. Compared to less recent literature, there is greater consensus also on the methods to estimate/calculate metabolic power (based on which C is calculated) at maximal speed; the number of studies that investigate C and its determinants at race pace has, indeed, increased recently.
Many factors are known to influence C through their effects on ηP, Fd, or both (e.g., sex, age, training …). To single out the effects of one of these factors, the other “confounding factors” must be kept constant but this is not always easy to do or even reckoned; some of the “discrepancies” reported in the literature about the determinants of C can be attributed to this issue. In particular, it is important to control for speed because, when speed increases, Fd is bound to increase, along with energy expenditure.
Swimming research is fascinating, the challenges imposed by the water environment are sometimes frustrating but also stimulate imaginative attempts to develop new methods to derive quantitative (and reasonable) data; compared to land locomotion much remains to be done in this filed. We hope that this review could serve as a general framework to understand the complex interplay between the many factors that are known to influence C, and hence swimming performance.
Abbreviations
- A :
-
Projected frontal area
- AO :
-
Swimming with arms only
- C :
-
Energy cost
- C d :
-
Drag coefficient
- \(\dot{E}\) :
-
Metabolic power
- \(\dot{E}_{{\max} }\) :
-
Maximal metabolic power
- \(\dot{E}_{\text{Aer}}\) :
-
Metabolic power (aerobic component)
- \(\dot{E}_{\text{Anl}}\) :
-
Metabolic power (anaerobic lactic component)
- \(\dot{E}_{\text{AnAl}}\) :
-
Metabolic power (anaerobic alactic component)
- F d :
-
Hydrodynamic resistance (drag force)
- F t :
-
Thrust force
- IVV:
-
Intra-cyclic variations in speed
- IdC:
-
Index of coordination
- LO:
-
Swimming with legs only
- Lab :
-
Blood lactate concentration
- OWS:
-
Open water swimming
- PCr:
-
Phosphocreatine concentration
- SF:
-
Stroke frequency
- SL:
-
Stroke length
- T :
-
“Leg sinking” torque
- t :
-
Exercise duration
- v :
-
Swimming speed
- v max :
-
Maximal swimming speed
- \(\dot{V}{\text{O}}_{2}\) :
-
Oxygen uptake
- \(\dot{V}{\text{O}}_{2{\max} }\) :
-
Maximal oxygen uptake
- \(\dot{W}_{\text{tot}}\) :
-
Total mechanical power
- \(\dot{W}_{\text{ext}}\) :
-
External mechanical power
- \(\dot{W}_{\text{int}}\) :
-
Internal mechanical power
- \(\dot{W}_{\text{d}}\) :
-
Drag power (mechanical power needed to overcome drag)
- \(\dot{W}_{\text{k}}\) :
-
“Wasted” mechanical power
- \(\dot{W}_{\text{t}}\) :
-
Thrust power (mechanical power needed to propel the body forward)
- WS:
-
Swimming the whole stroke
- η O :
-
Overall efficiency
- η P :
-
Propelling efficiency
- η d :
-
Drag efficiency
- η F :
-
Froude efficiency
- η H :
-
Hydraulic efficiency
- ρ :
-
Water density
- τ :
-
Time constant
- 3D:
-
Three-dimensional
References
Alexander RMN (1977) Swimming. In: Alexander RMN, Goldspink G (eds) Mechanics and energetics of animal locomotion. Chapman et Hall, London, pp 222–248
Alexander RMN (1983) Motion in fluids. Animal mechanics. Blackwell, Oxford, pp 183–233
American College of Sport Medicine (2018) Guidelines for exercise testing and prescription, 10th edn. Wolters Kluwer Philadelphia, Philadelphia
Baldari C, Fernandes RJ, Meucci M, Ribeiro J, Vilas-Boas JP, Guidetti L (2013) Is the new aquatrainer snorkel valid for VO2 assessment in swimming? Int J Sports Med 34(4):336–344
Baldassarre R, Bonifazi M, Zamparo P, Piacentini MF (2017) Characteristics and challenges of open water swimming performance: a review. Int J Sport Physiol Perf 12(10):1275–1284
Baldassarre R, Bonifazi M, Piacentini MF (2019) Pacing profile in the main international open water swimming competitions. Eur J Sport Sci 19(4):422–431
Barbosa T, Keskinen KL, Fernandes R, Colaço P, Lima AB, Vilas-Boas JP (2005) Energy cost and intracyclic variation of the velocity of the centre of mass in butterfly stroke. Eur J Appl Physiol 93:519–523
Barbosa T, Fernandes RJ, Keskinen KL, Colaço P, Cardoso C, Silva J, Vilas-Boas JP (2006) Evaluation of the energy expenditure in competitive swimming strokes. Int J Sports Med 27:894–899
Barbosa T, Keskinen KL, Fernandes RJ, Vilas-Boas JP (2008) The influence of stroke mechanics into energy cost of elite swimmers. Eur J Appl Physiol 103:139–149
Barbosa TM, Bragada JA, Reis VM, Marinho DA, Carvalho C, Silva AJ (2010) Energetics and biomechanics as determining factors of swimming performance: updating the state of the art. J Sci Med Sport 13:262–269
Basset DR, Flohr J, Duey WJ, Howley ET, Pein RL (1991) Metabolic responses to drafting during front crawl swimming. Med Sci Sports Exerc 23(6):744–777
Beneke R, Pollmann C, Bleif I, Leithauser RM, Hutler M (2002) How anaerobic is the wingate anaerobic test for humans? Eur J Appl Physiol 87:388–392
Buglione A, Lazzer S, Colli R, Introini E, di Prampero PE (2011) Energetics of best performances in elite kayakers and canoeists. Med Sci Sports Exerc 43:877–884
Capelli C (1999) Physiological determinants of best performances in human locomotion. Eur J Appl Physiol 80:298–307
Capelli C, Donatelli C, Moia C, Valier C, Rosa G, di Prampero PE (1990) Energy cost and efficiency of sculling a Venetian gondola. Eur J Appl Physiol 105:635–661
Capelli C, Termin B, Pendergast DR (1998) Energetics of swimming at maximal speed in humans. Eur J Appl Physiol 78:385–393
Cavagna GA, Kaneko M (1977) Mechanical work and efficiency in level walking and running. J Physiol 268:467–481
Chatard JC, Wilson B (2003) Drafting distance in swimming. Med Sci Sports Exerc 35(7):1176–1181
Chatard JC, Wilson B (2008) Effect of fatskin suits on performance, drag an energy cost of swimming. Med Sci Sports Exerc 40(6):1149–1154
Chatard JC, Padilla S, Cazorla G, Lacour JR (1985) Influence of body height, weight, hydrostatic lift and training on the energy cost of front crawl. NZ J Sports Med 13:82–84
Chatard JC, Lavoie JM, Bourgoin B, Lacour JR (1990a) a) The contribution of passive drag as a determinant of swimming performance. Int J Sports Med 11:367–372
Chatard JC, Bourgoin B, Lacour JR (1990b) Passive drag is still a good evaluator of swimming aptitude. Eur J Appl Physiol 59:399–404
Chatard JC, Lavoie JM, Lacour JR (1990c) Analysis of determinants of swimming economy in front crawl. Eur J Appl Physiol 61:88–92
Chatard JC, Lavoie JM, Lacour JR (1991) Energy cost of front crawl swimming in women. Eur J Appl Physiol 63:12–16
Chollet D, Chalies S, Chatard JC (2000) A new index of coordination for the crawl: description and usefulness. Int J Sports Med 21:54–59
Clemente-Suarez VJ, Dalamitros A, Riberio J, Sousa A, Fernandes RJ, Vilas-Boas JP (2017) Tracking the performance, energetics and biomechanics of international versus national level swimmers during a competitive season. Eur J Appl Physiol 112:811–820
Cortesi M, Fantozzi S, di Michele R, Zamparo P, Gatta G (2014) Passive drag reduction using full-body swimsuits: the role of body position. J Strength Cond Res 28(11):3164–3171
Cortesi M, Gatta G, Swaine I, Zamparo P, Konstantaki M (2019) Laboratory based ergometry for swimmers: a narrative review. J Sport Med Phys Fitness 59(9):1503–1512
Costa MJ, Bragada JA, Mejias JE, Louro H, Marinho DA, Silva AJ, Barbosa TM (2012) Effects of swim training on energetics and performance. Int J Sports Med 34:507–513
Costill DL, Kovalesky J, Porter D, Kirwanm J, Fielding R, King D (1985) Energy expenditure during front crawl swimming: predicting success in middle distance events. Int J Sport Med 6:266–270
Craig AB Jr, Pendergast DR (1979) Relationships of stroke rate, distance per stroke and velocity in competitive swimming. Med Sci Sports 11:278–283
Craig AB Jr, Skehan P, Pawelczyk JA, Boomer WL (1985) Velocity, stroke rate and distance per stroke during elite swimming competitions. Med Sci Sports Exerc 17(6):625–634
Daniel TL (1991) Efficiency in aquatic locomotion: limitations from single cells to animals. In: Blake RW (ed) Efficiency and economy in animal physiology. Cambridge University Press, Cambridge, pp 83–96
De Ioannon G, Cibelli G, Mignardi S, Antonelli A, Capranica L, Piacentini MF (2015) Pacing and mood changes while crossing the Adriatic Sea from Italy to Albania: a case study. Int J Sports Physiol Perform 10(4):520–523
Deschodt VJ, Arsac LM, Rouard AH (1999) Relative contribution of arms and legs in humans to propulsion in 25-m sprint front crawl swimming. Eur J Appl Physiol 80:192–199
di Prampero PE (1981) Energetics of muscular exercise. Rev Physiol Biochem Pharmacol 89:143–222
di Prampero PE (1986) The energy cost of human locomotion on land and in water. Int J Sports Med 7:55–72
di Prampero PE (2003) Factors limiting maximal performance in humans. Eur J Appl Physiol 90:420–429
di Prampero PE, Ferretti G (1999) The energetics of anaerobic muscle metabolism: a reappraisal of old and recent concepts. Resp Physiol 118:103–115
di Prampero PE, Davies CT, Cerretelli P, Margaria R (1970) An analysis of O2 debt contracted in submaximal exercise. J Appl Physiol 29(5):547–551
di Prampero PE, Cortili G, Celentano F, Cerretelli P (1971) Physiological aspects of rowing. J Appl Physiol 31:853–857
di Prampero PE, Pendergast D, Zamparo P (2011) Swimming economy (energy cost) and efficiency. In: Seifert L, Chollet D, Mujika I (eds) Word book of swimming: from science to performance. Nova Science Publisher, New York, pp 297–312
Donato AJ, Tench K, Glueck DH, Seals DR, Eskurza I, Tanaka H (2003) Declines in physiological functional capacity with age: a longitudinal study in peak swimming performance. J Appl Physiol 94:764–769
Dwyer J (1983) Marathon swimmers: physiologic characteristics. J Sports Med Phys Fitness 23(3):263–272
Fernandes RJ, Morais P, Keskinen KL, Seifert L, Chollet D, Vilas-Boas JP (2010) relationship between arm coordination and energy cost in front crawl swimming. In: Kjendlie PL, Stallman RK, Cabri J (eds) Biomechanics and medicine in swimming XI. Norwegian School of Sport Sciences, Oslo, pp 74–76
Figueiredo P, Zamparo P, Sousa A, Vilas-Boas JP, Fernandes RJ (2011) An energy balance of the 200 m front crawl race. Eur J Appl Physiol 111:767–777
Figueiredo P, Barbosa T, Vilas-Boas JP, Fernandes R (2012) Energy cost and body centre of mass’ 3D intracycle velocity variation in swimming. Eur J Appl Physiol 112:3319–3326
Figueiredo P, Toussaint HM, Vilas-Boas JP, Fernandes RJ (2013) Relation between efficiency and energy cost with coordination in aquatic locomotion. Eur J Appl Physiol 113:651–659
Formosa D, Mason B, Burkett B (2011) The force-time profile of elite front crawl swimmers. J Sports Sci 29(8):811–819
Fox RW, McDonald AT (1992) Fluid machines. Introduction to fluid machines. Wiley, New York, pp 544–625
Gatta G, Cortesi M, Di Michele R (2012) power production of the lower limbs in flutter kick swimming. Sports Biomech 11(4):480–491
Gatta G, Cortesi M, Fantozzi S, Zamparo P (2015) Planimetric frontal area in the four swimming strokes: implications for drag, energetics and speed. Human Mov Sci 39:41–54
Gatta G, Cortesi M, Swaine I, Zamparo P (2018) Mechanical power, thrust power and propelling efficiency: relationships with elite swimming performance. J Sport Sci 36(5):506–512
Gonjo T, McCabe C, Sousa A, Ribeiro J, Fernandes RJ, Vilas-Boas JP, Sanders R (2018) Differences in kinematics and energy cost between front crawl and backstroke below the anaerobic threshold. Eur J Appl Physiol 118(6):1107–1118
Gourgoulis V, Aggelousis N, Vezos N, Mavromatis G (2006) Effect of two different sized hand paddles on front crawl stroke kinematics. J Sports Med Phys Fitness 46:232–237
Gourgoulis V, Boli A, Aggeloussis N, Toubekis A, Antoniu P, Kasimatis P, Vezos N, Michalopulou M, Kambas A, Mavromatis G (2014) The effects of leg kick on sprint front crawl swimming. J Sport Sci 32(3):278–289
Greidanus A, Delfos R, Westerwell J (2012) Rowing faster by surface treatment. Procedia Eng 34:361–366
Havriluk R (2005) Performance level differences in swimming: a meta-analysis of passive drag forces. Res Q Exerc Sport 76:112–118
Havriluk R (2007) Variability in measurements of swimming forces: a meta-analysis of passive and active drag. Res Q Exerc Sport 78:32–39
Holmer I (1972) Oxygen uptake during swimming in man. J Appl Physiol 33:502–509
Holmer I (1974) Energy cost of arm stroke, leg kick and the whole stroke in competitive swimming styles. Eur J Appl Physiol 33:105–118
Janssen M, Wilson BD, Toussaint HM (2009) Effects of drafting on hydrodynamic and metabolic responses in front crawl swimming. Med Sci Sports Exerc 41(4):837–843
Kautz SA, Neptune RR (2002) Biomechanical determinants of pedalling energetics: internal and external work are not independent. Exerc Sport Sci Rev 30:159–165
Keskinen KL, Rodriguez FA, Keskinen OP (2003) Respiratory snorkel and valve system for breath-by-breath gas analysis in swimming. Scand J Med Sci Sports 13:322–329
Kjendlie PL, Stallman RK (2008) Drag characteristics of competitive swimming children and adults. J Appl Biomech 24:35–42
Kjendlie PL, Stallman RK (2011) Morphology and swimming performance. In: Seifert L, Chollet D, Mujika I (eds) World book of swimming: from science to performance, pp 203–221
Kjendlie PL, Ingjer F, Stallman RK, Stray-Gundersen J (2004a) Factors affecting swimming economy in children and adults. Eur J Appl Physiol 93:65–74
Kjendlie PL, Ingjer F, Madsen O, Stallman RK, Stray-Gundersen J (2004b) Differences in the energy cost between children and adults during front crawl swimming. Eur J Appl Physiol 91:473–480
Knechtle B, Rosemann T, Lepers R, Rust CA (2014) Women outperform men in ultradistance swimming: the Manhattan Island Marathon Swim from 1983 to 2013. Int J Sport Physiol Perf 9:913–924
Komar J, Lepretre PM, Alberty M, Vantorre J, Fernandes RJ, Hellard P, Chollet D, Seifert L (2012) Effect of increasing energy cost on arm coordination in elite sprint swimmers. Hum Mov Sci 31:620–629
Lauer J, Olstad BH, Minetti AE, Kjendlie PL, Rouard AH (2015) Breastroke swimmers moderate internal work increases toward the highest stroke frequencies. J Biomech 48(12):3012–3016
Lyttle A, Blanksby B (2011) A review of swimming dive starting and turning performance. In: Seifert L, Chollet D, Mujika I (eds) Word book of swimming: from science to performance. Nova Science Publishers, Hauppauge, pp 425–442
Lyttle AD, Blanksby BA, Elliott BC, Lloyd DG (2000) Net force during tethered simulation of underwater streamlined gliding and kicking techniques of freestyle turn. J Sport Sci 18(10):801–807
Medbø J, Mohn AC, Tabata I, Bahr R, Vaage O, Sejersted O (1988) Anaerobic capacity determined by maximal accumulated O2 deficit. J Appl Physiol 64:50–60
Mejias JE, Bragada JA, Costa MJ, Reis VM, Garrido ND, Barbosa TM (2014) “Young” masters vs. elite swimmers: comparison of performance, energetics, kinematics and efficiency. Int Sport Med J 15(2):165–177
Minetti AE (2004) Passive tools for enhancing muscle-driven motion and locomotion. J Exp Biol 207:1265–1272
Minetti AE, Capelli C, Zamparo P, di Prampero PE, Saibene F (1995) Effects of stride frequency on mechanical power and energy expenditure of walking. Med Sci Sports Exer 27(8):1194–1202
Minetti AE, Machtsiras G, Masters J (2009) The optimum finger spacing in human swimming. J Biomech 42:2188–2190
Mollendorf JC, Termin A, Oppenheim E, Pendergast DR (2004) Effect of swim suit design on passive drag. Med Sci Sports Exerc 36:1029–1035
Montpetit R, Leger L, Lavoie JM, Cazorla GA (1981) VO2 peak during free swimming using the backward extrapolation of the recovery curve. Eur J Appl Physiol 47:385–391
Moreira M, Morais JE, Marhino DA, Silva AJ, Babosa TM, Costa MJ (2014) Growth influences biomechanical profile of talented swimmers during the summer break. Sport Biomech 13(1):62–74
Morris KS, Osborne MA, Shephard ME, Skinner TL, Jenkins DG (2016) Velocity, aerobic power and metabolic cost of whole body and arms only front crawl swimming at various stroke rates. Eur J Appl Physiol 116(5):1075–1085
Morris KS, Osborne MA, Shephard ME, Jenkins DG, Skinner TL (2017) Velocity, oxygen uptake and metabolic cost of pull, kick and whole body swimming. Int J Sport Physiol Perf 12:1046–1051
Mujika I, Padilla S (2000) Detraining: loss of training-induced physiological and performance adaptations. Part I. Sport Med 30(2):79–87
Narita K, Nakashima M, Takagi H (2017) Developing a methodology for estimating the drag in front crawl swimming at various velocities. J Biomech 54:123–128
Ogita F, Tabata I (1993) Effect of hand paddle aids on oxygen uptake during arm–stroke–only swimming. Eur J Appl Physiol 66(6):489–493
Ogita F, Hara M, Tabata I (1996) Anaerobic capacity and maximal oxygen uptake during arm stroke, leg kicking and whole body swimming. Acta Physiol Scand 157:435–441
Ogita F, Onodera T, Tabata I (1999) Effect of hand paddles on anaerobic energy release during supramaximal swimming. Med Sci Sports Exerc 31(5):729–735
Ogita F, Onodera T, Tamaki H, Toussaint H, Hollander P (2003) Wakayoshi K (2003) Metabolic profile during exhaustive arm stroke, leg kick and whole body swimming lasting 15 s to 10 min. In: Chatard J-C (ed) Biomechanics and medicine in swimming IX. Publications de l’ Universite de Saint-Etienne, Saint-Etienne, pp 361–366
Payton CJ, Sanders RH (2011) Body-roll in front crawl swimming. In: Seifert L, Chollet D, Mujika I (eds) Word book of Swimming: from science to performance. Nova Science Publishers, Hauppauge, pp 173–190
Pendergast DR, di Prampero PE, Craig AB Jr, Wilson DR, Rennie DW (1977) Quantitative analysis of the front crawl in men and women. J Appl Physiol 43:475–479
Pendergast DR, Bushnell D, Wilson DR, Cerretelli P (1989) Energetics of kayaking. Eur J Appl Physiol 59:342–350
Pendergast DR, Zamparo P, di Prampero PE, Capelli C, Cerretelli P, Termin A, Craig A, Bushnell D, Paschke D, Mollendorf J (2003) Energy balance of human locomotion in water. Eur J Appl Physiol 90:377–386
Pendergast DR, Mollendorf J, Zamparo P, Termin A, Bushnell D, Paschke D (2005) The influence of drag on human locomotion in water. Undersea Hyperb Med 32:45–57
Peronnet F, Thibault G (1989) Mathematical analysis of running performance and word running records. J Appl Physiol 67:453–465
Peterson-Silveira R, de Souza Castro F, Figuereido P, Vilas-Boas JP, Zamparo P (2017) The effects of leg kick on swimming speed and arm-stroke efficiency in the front crawl. Int J Sport Physiol Perform 12(6):728–735
Peyrebrune MC, Toubekis AG, Lakomy HKA, Nevill ME (2014) Estimating the energy contribution during single and repeated sprint swimming. Scand J Med Sci Sports 24:369–376
Poujade B, Hautier CA, Rouard A (2002) Determinants of the energy cost of front crawl swimming in children. Eur J Appl Physiol 87:1–6
Psycharakis SG, Naemi R, Connaboy C, McCabe C, Sanders RH (2010) Three-dymensional analysis of intracycle velocity fluctuations in front crawl swimming. Scand J Med Sci Sports 20(1):128–135
Ratel S, Poujade B (2009) Comparative analysis of the energy cost during front crawl swimming in children and adults. Eur J Appl Physiol 105:543–549
Reis VM, Marinho DA, Policarpo FB, Carneiro AL, Baldari C, Silva AJ (2010) Examining the accumulated oxygen deficit method in front crawl swimming. Int J Sports 31:421–427
Ribeiro J, Figueiredo P, Sousa A, Monteiro J, Pelarigio J, Vilas-Boas JP, Toussaint H, Fernandes RF (2015) VO2 kinetics and metabolic contribution during full and upper body extreme swimming intensities. Eur J Appl Physiol 115:1117–1124
Ribeiro J, Figueiredo P, Guidetti L, Alves F, Toussaint H, Vilas-Boas JP, Baldari C, Fernandes RJ (2016) AquaTrainer snorkel does not increase hydrodynamic drag but influences turning time. Int J Sports Med 37(04):324–328
Ribeiro J, Figueiredo P, Morais S, Alves F, Toussaint H, Vilas-Boas JP, Fernandes RJ (2017) Biomechanics, energetics and coordination during extreme swimming intensity: effect of performance level. J Sport Sci 35(16):1614–1621
Rodríguez FA (2000) Maximal oxygen uptake and cardiorespiratory response to maximal 400-m free swimming, running and cycling tests in competitive swimmers. J Sports Med Phys Fitness 40(2):87–95
Rodriguez FA, Mader A (2011) Energy systems in swimming. In: Seifert L, Chollet D, Mujika I (eds) Word book of swimming: from science to performance. Nova Science Publisher, New York, pp 225–240
Rodriguez L, Veiga S (2018) Effect of the pacing strategies on the open-water 10-km world swimming championships performances. Int J Sport Physiol Perform 13(6):694–700
Rossiter HB, Ward SA, Kowalchuk JM, Howe FA, Griffiths FA, Whipp BJ (2002) Dynamic asymmetry of phosphocreatine concentration and O2 uptake between the on- and off-transients of moderate and high-intensity exercise in humans. J Physiol 541:991–1002
Sandbakk Ø, Solli GS, Holmberg HC (2018) Sex differences in World record performance: the influence of sport discipline and competition duration. Int J Sports Physiol Perform 2:1–7
Scurati R, Gatta G, Michielon G, Cortesi M (2019) Techniques and considerations for monitoring swimmer’s passive drag. J Sports Sci 37(19):1168–1180
Seifert L, Boulesteix L, Chollet D (2004) Effect of gender on the adaptation of arm coordination in front crawl. Int J Sports Med 25:217–223
Seifert L, Chollet D, Chatard JC (2007) Kinematic changes during a 100 m front crawl: effects of performance level and gender. Med Sci Sports Exerc 39:1784–1793
Seifert L, Toussaint H, Alberty M, Schnitzel C, Chollet D (2010a) Arm coordination, power and swim efficiency in national and regional front crawl swimmers. Hum Mov Sci 29:426–439
Seifert L, Komar J, Lepretre PM, Lemaitre F, Chavallard F, Alberty M, Houel N, Hausswirth C, Chollet D, Hellard P (2010b) Swim speciality affects energy cost and motor organization. Int J Sports Med 31:624–630
Seifert L, Komar J, Crettenand F, Dadashi F, Aminian K, Millet GP (2014a) Inter.limb coordination and energy cost in swimming. J Sci Med Sport 17:439–444
Seifert L, Komar J, Crettenand F, Millet G (2014b) Coordination pattern adaptability: energy cost of degenerate behaviors. PLoS One 9(9):e107839. https://doi.org/10.1371/journal.pone.0107839
Sharp RL, Costill DL (1989) Influence of body air removal on physiological responses during breaststroke swimming. Med Sci Sports Exerc 21(5):576–580
Smith HK, Montpetit RR, Perrault H (1988) The aerobic demand of backstroke swimming, and its relation to body size, stroke technique, and performance. Eur J Appl Physiol 58(1–2):182–188
Sousa A, Figueiredo P, Zamparo P, Vilas Boas JP, Fernandes RJ (2013) Anaerobic alactic energy assessment in middle distance swimming. Eur J Appl Physiol 113(8):2153–2158
Sparrow WA, Newell KM (1998) Metabolic energy expenditure and the regulation of movement economy. Psychon Bull Rev 5:173–196
Tam E, Rossi H, Moia C, Berardelli C, Rosa G, Capelli C, Ferretti G (2012) Energetics of running in top-level marathon runners from kenya. Eur J Appl Physiol 112(11):3797–3806
Tanaka H, Seals DR (1997) Age and gender interactions in physiological functional capacity: insight from swimming performance. J Appl Physiol 83:846–851
Termin B, Pendergast DR (2000) Training using the stroke frequency-velocity relationship to combine biomechanical and metabolic paradigms. J Swim Res 14:9–17
Tor E, Pease DL, Ball KA (2015) How does drag affect the underwater phase of a swimming start. J Appl Biomech 31:8–12
Toussaint HM, Hollander AP (1994) Energetics of competitive swimming. Implications for training programmes. Sports Med 18(6):384–405
Toussaint HM, Knops W, De Groot G, Hollander AP (1990a) The mechanical efficiency of front crawl swimming. Med Sci Sports Exerc 22:409–415
Toussaint HM, de Looze M, van Rossem B, Leijdekkers M, Dignum H (1990b) The effect of growth on drag in young swimmers. J Appl Biomech 6:18–28
Toussaint HM, Janssen T, Kluft M (1991) Effect of propelling surface size on the mechanics and energetics of front crawl swimming. J Biomec 24:205–211
Toussaint HM, Roos PE, Kolmogorow S (2004) The determinants of drag in front crawl swimming. J Biomech 37:1655–1663
Toussaint HM, Carol A, Kranenborg H, Truijens MJ (2006) Effect of fatigue on stroking characteristics in arms-only 100 m front crawl race. Med Sci Sports Exerc 38(9):1635–1642
Tsunokawa T, Mankyu J, Takagi H, Ogita F (2019) The effect of using paddles on hand propulsive forces and Froude efficiency in arm stroke only front crawl swimming at various velocities. Hum Mov Sci 64:378–388
Van Handle PA, Katz A, Morrow JR, Troup JP, Daniels JT, Bradleym PW (1988) Aerobic economy and competitive performance of US elite swimmers. In: Ungerechts BE, Wilke K, Reischle K (eds) Swimming science V. Human kinetics, Champaign, pp 219–228
Van Houwelingen J, Willemsen DHJ, Kunnen RPJ, van Heijst GF, Grift EJ, Breugem WP, Delfos E, Westerweel J, Vlercx HJH, van de Water W (2017) The effect of finger spreading on drag of the hand in human swimming. J Biomech 63:67–73
VanHeest JL, Mahoney CE, Herr L (2004) Characteristics of elite open- water swimmers. J Strength Cond Res 18(2):302
Vennell R, Pease D, Wilson B (2006) Wave drag on human swimmers. J Biomech 39:664–671
Vilas-Boas JP (1996) Speed fluctuations and energy cost of different breaststroke techniques. In: Troup JP, Hollander AP, Strasse D, Trappe SW, Cappaert JM, Trappe TA (eds) Biomechanics and medicine in swimming VII. E & FN Spon, London, pp 167–171
Vilas-Boas JP, Santos P (1994) Comparison of swimming economy in three breaststroke techniques. In: Miyashita M, Richardson AB (eds) Medicine and science in aquatic sports. Kargel, Basel, pp 48–54
Vogel S (1994) Life in moving fluid. Princeton University Press, Princeton, pp 132–155
Vogt P, Rüst CA, Rosemann T, Lepers R, Knechtle B (2013) Analysis of 10 km swimming performance of elite male and female open-water swimmers. Springerplus 2:603
Wakayoshi K, D’Acquisto LJ, Cappaert JM, Troup JP (1995) Relationship between oxygen uptake, stroke rate and swimming velocity in competitive swimming. Int J Sports Med 16(1):19–23
Wilkie DR (1980) Equations describing power input by humans as a function of duration of exercise. In: Cerretelli P, Whipp BJ (eds) Exercise bioenergetics and gas exchange. Elsevier, Amsterdam, pp 75–80
Wilson B, Thorp R (2003) Active drag in swimming. In: Chatard JC (ed) Biomechanics and medicine in swimming IX. University Press, Saint Etienne, pp 15–20
Yanai T (2001) Rotational effect of buoyancy in front crawl swimming: does it really cause the legs to sink? J Biomech 34:235–243
Yanai T, Wilson BD (2008) How does buoyancy influence front crawl performance? Exploring the assumptions. Sports Technol 1(2–3):89–99
Zamparo P (2006) Effects of age and gender on the propelling efficiency of the arm stroke. Eur J Appl Physiol 97:52–58
Zamparo P, Swaine I (2012) Mechanical and propelling efficiency in swimming derived from exercise using a laboratory based whole-body swimming ergometer. J Appl Physiol 113:584–594
Zamparo P, Antonutto G, Capelli C, Francescato MP, Girardis M, Sangoi R, Soule RG, Pendergast DR (1996) Effects of body size, body density, gender and growth on underwater torque. Scand J Med Sci Sports 6:273–280
Zamparo P, Capelli C, Guerrini G (1999) Energetics of kayaking at sub-maximal and maximal speeds. Eur J Appl Physiol 80(6):542–548
Zamparo P, Capelli C, Cautero M, Di Nino A (2000) Energy cost of front crawl swimming at supramaximal speeds and underwater torque in young swimmers. Eur J Appl Physiol 83:487–491
Zamparo P, Pendergast D, Termin A, Minetti AE (2002) How fins affect the economy and efficiency of human swimming. J Exp Biol 205:2665–2676
Zamparo P, Pendergast DR, Mollendorf J, Termin A, Minetti AE (2005a) An energy balance of front crawl. Eur J Appl Physiol 94:134–144
Zamparo P, Bonifazi M, Faina M, Milan A, Sardella F, Schena F, Capelli C (2005b) Energy cost of swimming of elite long distance swimmers. Eur J Appl Physiol 94:697–704
Zamparo P, Carignani G, Plaino L, Sgalmuzzo B, Capelli C (2008a) Energetics of locomotion with pedal driven watercrafts. J Sport Sci 26(1):75–81
Zamparo P, Lazzer S, Antoniazzi C, Cedolin S, Avon R, Lesa C (2008b) The interplay between propelling efficiency, hydrodynamic position and energy cost of front crawl in 8 to19-year-old swimmers. Eur J Appl Physiol 104:689–699
Zamparo P, Gatta G, Capelli C, Pendergast DR (2009) Active and passive drag, the role of trunk incline. Eur J Appl Physiol 106:195–205
Zamparo P, Capelli C, Pendergast DR (2011) Energetics of swimming: a historical perspective. Eur J Appl Physiol 111:367–378
Zamparo P, Dall’Ora A, Toneatto A, Cortesi M, Gatta G (2012a) The determinants of performance in master swimmers: age related changes in propelling efficiency, hydrodynamic position and energy cost of front crawl. Eur J Appl Physiol 112(12):3949–3957
Zamparo P, Gatta G, di Prampero PE (2012b) The determinants of performance in master swimmers: an analysis of master world records. Eur J Appl Physiol 112:3511–3518
Zamparo P, Turri E, Peterson Silveria R, Poli A (2014) The interplay between arms-only propelling efficiency, power output and speed in master swimmers. Eur J Appl Physiol 144(6):1259–1268
Zamparo P, Baldassarre R, Bonifazi M, Piacentini MF (2019) Open water swimming. In: Datta S, Bagchi D (eds) Extreme and rare sports. CRC Press, Taylor & Francis Group, Boca Raton, pp 11–33
Zingg MA, Rüst CA, Rosemann T, Lepers R, Knechtle B (2013) Analysis of swimming performance in FINA World Cup long distance open water races. Extrem Physiol Med 3(1):2. https://doi.org/10.1186/2046-7648-3-2
Zocca R, Toubekis A, Freitas L, Silva AF, Azevedo R, Vilas-Boas JP, Pyne DB, De Sousa-Castro F, Fernandes RJ (2017) Effects of detraining in age group swimmers, performance, energetics and kinematics. J Sport Sci 37(13):1490–1498
Author information
Authors and Affiliations
Contributions
PZ conceptualized and wrote the manuscript. MC and GG performed the literature search and critically revised the work. All authors read and approved the final manuscript.
Corresponding author
Ethics declarations
Conflict of interest
The authors report no conflict of interest.
Additional information
Communicated by Michael Lindinger.
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
About this article

Cite this article
Zamparo, P., Cortesi, M. & Gatta, G. The energy cost of swimming and its determinants. Eur J Appl Physiol 120, 41–66 (2020). https://doi.org/10.1007/s00421-019-04270-y
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00421-019-04270-y