
Abstract
Purpose
To compare maximal and rapid force characteristics, as well as fatigability, between traditional (TRT) and explosive (ERT) resistance-trained men.
Methods
Fourteen TRT (mean age = 25 years) and twelve ERT (mean age = 22 years) men performed rapid maximal contractions followed by an isokinetic fatigue protocol consisting of 50 maximal knee extension (KE) and flexions (KF) at a moderate speed (180° s−¹). Baseline measures included: isokinetic peak torque (PT), isometric rate of torque development (RTD0–50), peak acceleration (ACCmax), and peak velocity (Vmax). Changes in torque with fatigue were used to calculate a fatigue index (FI%).
Results
The ERT group (M ± SD; 1199.05 ± 404.12) displayed a significantly higher isometric RTD0–50 (p = 0.049) during KE than the TRT group (931.73 ± 244.75). No other significant differences in the dependent variables (PT, FI%, ACCmax, Vmax; all p ≥ 0.05) were observed between groups (TRT vs. ERT) for either of the muscle groups (KE and KF).
Conclusions
The results of the present study indicated that only knee extension RTD was able to discriminate between the two groups. These findings suggest that rapid force production may be more sensitive at distinguishing training-specific muscular adaptations than peak acceleration or velocity.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Resistance training has been shown to elicit several essential benefits related to human performance, including, but not limited to, muscular hypertrophy, increased strength, power, and speed (Hooper et al. 2013). Manipulation of resistance-training programming characteristics (i.e., number of sets, repetitions, movement velocity, length of rest periods, and external load) has been shown to produce differential adaptive responses in skeletal muscle specific to the various acute and chronic stressors (Campos et al. 2002; Jones and Rutherford 1987; Mangine et al. 2008; Schuenke et al. 2012; Winchester et al. 2008). These responses have been shown to be influenced by the structure and implementation of resistance activity, as well as the training status (i.e., beginner, intermediate, or advanced) of the individuals involved (Abernethy et al. 1994).
Although resistance training can serve many purposes, it is suggested that the specificity principle may allow for optimal performance. This principle recommends that the exercise prescription should be designed to be similar to the athletic movements that would be performed by the individual (Hansen 1961, 1963; Murray et al. 2007; Pereira and Gomes 2003; Rasch and Morehouse 1957). Recent examinations showed that the greatest gains in force and power production are seen at movement velocities comparable to that of training (Murray et al. 2007; Pereira and Gomes 2003). This is further demonstrated by Behm (1995), who revealed, to maintain high-velocity-specific adaptations in a power training program, the speed of contraction must be high/fast (≥ 240° s− 1) (Murray et al. 2007; Pereira and Gomes 2003). While exercise prescription can have direct effects on performance, studies with younger adults have shown differential responses with the development of muscle power and rapid force generation when comparing explosive (ERT) versus heavy-resistance training (Caserotti et al. 2008; Newton et al. 1999). For example, Newton et al. (1999) found significant increases in vertical jump ability when performing explosive based movements compared to traditional resistance training (TRT). Although power has been shown to increase with TRT (Channell and Barfield 2008; Jozsi et al. 1999), the specificity of ERT may have a greater influence on the ability to apply force rapidly. For example, TRT tends to focus on higher intensities at lower speeds (Channell and Barfield 2008), whereas ERT can be defined as movements (e.g., snatch, hang clean, power clean, and push jerk) in which maximum or near maximum rates of force development are attained (Stone and O’Bryant 1987).
Previous authors have attempted to understand specific training adaptations related to increases in strength and power production (Fielding et al. 2002; Judge et al. 2003). For example, Fielding et al. (2002) found that a ERT program significantly improved peak power, and was equally efficient at increasing muscle strength compared to TRT. In addition to prior training studies, previous authors have assessed neuromuscular differences between individuals with different training backgrounds (Häkkinen and Keskinen 1989; Lattier et al. 2003). Recently, a trend in ERT has incorporated workouts consisting of power movements in combination with other varied functional movements performed at high intensity or to failure (Bergeron et al. 2011; Hak et al. 2013; Smith et al. 2013). Smith et al. (2013) labeled this type of training as high-intensity power training which focused on sustained power output and use of multiple joint movements without a prescribed rest period. While previous literature suggests specific muscular adaptations occur following resistance-training interventions such as increased maximal force (Newton et al. 2002), power (Murray et al. 2007; Newton et al. 2002), hypertrophy (McCaulley et al. 2009; Schuenke et al. 2012), increases in Type II fibers (Schuenke et al. 2012), and initial training statuses (sedentary vs. trained) (Alway et al. 1988; Newton et al. 2008), it may be informative to assess specific rapid force characteristics and fatigue-induced responses of maximal strength between different resistance-trained groups to further elucidate these muscular adaptations related to specificity of training.
Due to the training specificity and the aforementioned observed training adaptations of TRT and ERT, it may be of interest to examine the potential differences in fatigue-induced responses to performance measures, (e.g., maximal strength), between two resistance-trained groups. Specifically, fatigue has been defined as any reduction in the force-producing capacity of a muscle during a maximal voluntary contraction (Gandevia 1992), as well as an inability to maintain a given maximal strength level when performing repeated maximal contractions (Mathiassen 1989). This reduction in force-producing capacity may lead to deficits in subsequent muscular performance, thus hindering physical ability post-exercise.
Although a majority of muscle fatigue literature is based upon the use of intermittent and/or sustained isometric contractions (Bilodeau et al. 2001; Conchola et al. 2015, 2013; Corcos et al. 2002; Pääsuke et al. 1999), the nature of most motor tasks in athletic and voluntary physical activities may lend itself, rather, to the assessment of dynamic muscle contractions (Izquierdo et al. 2009; Wadden et al. 2012). Thus, the ability to identify significant differences in strength and rapid force characteristics between two dissimilarly trained groups following a dynamic fatigue protocol, may lead to further understanding of specific acquired adaptations to training. Therefore, the purpose of this study was to compare the rapid force characteristics and dynamic fatigability between ERT and TRT resistance-trained men. We hypothesized that the ERT group would display greater absolute rapid force characteristics and fatigue resistance, but lower strength when compared to the TRT group.
Methods
Participants
Fourteen traditional-resistance-trained (TRT) (mean ± SD: age = 24.79 ± 3.12 years, height = 179.86 ± 6.25 cm, mass = 94.29 ± 14.15 kg) and twelve explosive resistance-trained (ERT) (mean ± SD: age = 21.83 ± 1.90 years, height = 176.74 ± 5.88 cm, mass = 85.47 ± 11.57 kg) males volunteered to participate in this study. All participants reported being consistently engaged in a structured TRT (mean ± SD: 6.26 ± 3.31 years) or ERT (mean ± SD: 4.92 ± 2.80 years) resistance-training program for a minimum of 6 months, ≥ 3 times per week, prior to the study. Participants who reported their workouts as a combination of strength (≤ 6 repetitions at ≥ 85% 1RM) and hypertrophy (6–12 repetitions at 67–85% 1RM) movements (i.e., back squat, bench press, deadlift, bent over row, and shoulder press) were identified as TRT. Participants who reported workouts consisting of power movements (i.e., snatch, hang clean, power clean, and push jerk) with a goal of 1–2 repetitions for a single-effort (80–90% 1RM) or 3–5 repetitions for multiple effort (75–85% 1RM) events in combination with other varied functional movements performed at high intensity or to failure were identified as ERT. No participants reported current or ongoing musculoskeletal injuries of the lower extremity within the previous 12 months prior to testing. The University Institutional Review Board for human subject’s research approved this study, and each participant signed an informed consent document and pre-exercise health history questionnaire before testing began.
Procedures
This was a descriptive study to investigate group differences in strength, rate of torque development (RTD), velocity (Vmax), acceleration (ACCmax), and fatigability of the knee flexors (KF) and extensors (KE). Each participant visited the laboratory on 2 occasions separated by ~ 48–72 h. During the first visit, participants were familiarized with isometric and isokinetic maximal voluntary contractions (MVCs), as well as the experimental fatigue protocol. During the second visit, participants completed isokinetic and isometric MVCs of the KF and KE prior to 50 isokinetic muscle actions of both muscle groups at 180° s−¹ (Thorstensson and Karlsson 1976).
Isometric RTD assessments
Maximal isometric rapid strength testing was performed on the right leg using a Biodex System 3 isokinetic dynamometer (Biodex Medical Systems, Inc., Shirley, NY, USA). Participants were seated with restraining straps over the trunk, pelvis, and thigh, with the input axis of the dynamometer aligned with the axis of rotation of the knee. All isometric RTD assessments for the KE and KF were performed at knee angles of 120° and 150°, respectively (full extension = 180°). Prior to maximal isometric rapid strength testing, participants performed a 5-min warm-up on a cycle ergometer (Monark Exercise 828E, Vansbro, Sweden) at a self-selected low-intensity. In addition, 3 submaximal isokinetic muscle actions were performed at 60° s−¹ at approximately 75% of their perceived maximal effort for both the KE and KF. Following the submaximal contractions and prior to experimental testing, each participant performed 3 isometric MVCs of the KE and KF with 1 min of recovery between each contraction. The participants were verbally instructed to ‘push’ or ‘pull,’ “as hard and fast as possible” for a total of 3–4 s for all MVCs (Thompson et al. 2013).
Isokinetic velocity assessments
Similar to the aforementioned maximal isometric testing procedures, maximal isokinetic velocity (Vmax) testing was performed on the right leg using the isokinetic dynamometer, in isokinetic mode at 500° s−¹. All isokinetic assessments started at knee angles of 90° and ~ 180° (full extension) for the KE and KF, respectively. Each participant performed 3 isokinetic MVCs through ~ 90° of range of motion (ROM) for the KE and KF with 1 min of recovery between each contraction. Vmax was used to assess the maximal shortening velocity of the muscle–limb unit in which no resistance (with the exception of the lever arm) was provided throughout the duration of the contraction (i.e., velocity of the dynamometer was set above all subjects’ maximum velocity capacities), in accordance with the procedures of Thompson et al. (2014). The participants were verbally instructed to ‘push’ or ‘pull,’ “as fast as possible”.
Experimental fatigue protocol
Five minutes following all Pre MVCs, participants performed the fatigue protocol consisting of 50 continuous repetitions of dynamic isokinetic contractions of the KE and KF at 180° s−¹ (medium velocity) through ~ 90° ROM (Thorstensson and Karlsson 1976). Participants were seated with restraining straps across the trunk, pelvis, and thigh identical to all MVC testing procedures. Additionally, participants started the protocol with their right limb at a resting knee angle of 90°, and their lower limb secured to the dynamometer lever arm, just above the ankle. During the experimental protocol, participants were asked to provide maximal effort for each muscle action and to ‘push’ or ‘pull’, as hard as they can throughout the entire protocol. Experimental testing was terminated at the completion of all 50 maximal repetitions. Verbal encouragement was provided to the participants during the entire protocol.
Signal processing
Torque (N·m), rate of torque development (RTD; N m s− 1), and angular velocity (° s− 1) signals were sampled simultaneously at 2 kHz with a Biopac data acquisition system (MP100WSW, Biopac Systems, Inc.; Santa Barbara, CA, USA), stored on a personal computer (Dell Inspiron 8200, Dell Inc., Round Rock, TX, USA), and processed off-line with custom-written software (LabVIEW version 16.0, National Instruments, Austin, TX, USA). The torque signal was smoothed using a 25 ms zero-shift moving average. All subsequent analyses were performed on the scaled and filtered torque signal. Isometric MVC PT was determined as the highest 25 ms epoch during the entire 3–4 s MVC plateau (Conchola et al. 2013; Thompson et al. 2013). Absolute RTD was calculated from the linear slope of the torque–time curve (Δvelocity/Δtime) over the interval of 0–50 ms (RTD0–50). Isokinetic PT was attained from the first and last few repetitions of the fatigue protocol. Vmax (° s− 1) was calculated as the highest velocity attained during the unloaded MVC. Acceleration (ACCmax; deg·s− 2) was determined as the 10 ms that demonstrated the highest linear slope of the velocity–time curve (Δvelocity/Δtime). These procedures were used to obtain the linear portion of the rate of rise in velocity, while simultaneously excluding the deceleration or “rounding off” of the signal observed at the edge of the velocity plateau. The onset of velocity was determined as the point when the velocity signal reached a threshold 2° s− 1 above baseline. The MVC with the highest PT, RTD0–50, Vmax, and ACCmax prior to the experimental protocol were used for all analyses (Thompson et al. 2014). Isokinetic 180° s− 1 FI% was calculated using initial and final PT which consisted of the average of the three muscle actions with the highest and lowest PT values during the fatigue protocol. All fatigue indices were calculated as “(Final–Initial) / Initial × 100” (Thorstensson and Karlsson 1976).
Statistical analysis
Independent samples t tests were run for the KE and KF between groups (traditional vs. explosive) for each dependent variable (isokinetic PT, isokinetic FI%, isometric RTD0–50, Vmax, and ACCmax). Cohen’s d effect sizes and 95% confidence intervals were run for the KF and KE for each dependent variable assessed for the TRT and ERT groups. PASW software version 23.0 (SPSS Inc, Chicago, IL, USA) was used for all statistical analyses. An alpha level of p ≤ 0.05 was considered significant for all comparisons.
Results
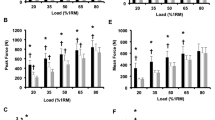
Means and SD values for all dependent variables are presented in Table 1, and the effect sizes are shown in Fig. 1. The ERT group (M ± SD; 1199.05 ± 404.12) displayed a significantly higher baseline (absolute) isometric RTD0–50 (p = 0.049) for the KE compared to the TRT group (931.73 ± 244.75) (Fig. 2). No other significant differences were observed between groups (TRT vs. ERT) for either of the muscle groups (KE and KF) and the dependent variables (isokinetic PT, isokinetic FI%, Vmax, ACCmax; p ≥ 0.05).

Cohen’s d effect sizes and 95% confidence intervals for the knee flexors and extensors for each dependent variable measured between the traditional (TRT) and explosive (ERT) resistance-trained groups. Effect sizes were calculated such that a negative value signified that TRT had a greater mean, while a positive value showed that ERT had a greater mean. *Significant difference between groups (p ≤ 0.05)

Early phase rate of torque development (RTD0–50) scores for each individual assessed combined with the group means and standard deviations. *Significant difference between groups (p = 0.049)
Discussion
The primary finding of the present study was the lack of differences between groups despite using variables that should be sensitive to training specificity. However, a significant difference was observed for knee extension RTD0–50, in which the ERT group had greater early RTD values compared to the TRT group. Besides RTD0–50, no differences were observed for any other variables between training statuses. The lack of differences observed in our study may have been attributed to the training status of the groups. Although there were obvious differences in workout regiments, on average each group had been resistance training for 4–6 years, which could have led to an overlap in types of resistance-training exercises and consequently similar responses to training adaptations. Although our investigation was not a training study, a recent study by MacDonald et al. (2012) supports our findings. The authors compared the effects of 6 weeks of TRT, plyometric training, and complex training on measures of strength (back squat, dead lift, and calf raise) and observed no differences between groups. Additionally, the assessments utilized in this study may not have been specific enough to distinguish the differences between the two groups. The addition of more functional and/or invasive measures may reveal further dissimilarities between the groups training adaptations.
While early rate of force development (0–100 ms) is important for a variety of movements (e.g., sprinting, cutting, kicking, etc.), a potential reason for the present study’s findings with knee extension RTD0–50 could be related to the speed and quickness that ERT lifters perform movements compared to the TRT group. For example, rapid force production determines the magnitude of acceleration in the initial phase of a movement and ultimately affects the velocity of the movement (Aagaard et al. 2002; Kraemer and Newton 2000). Additionally, previous studies that have reported increases in early RTD involved exercises that were executed with maximal acceleration effort (i.e., RFD/RTD) (Behm and Sale 1993; Caserotti et al. 2008; Suetta et al. 2004; Van Cutsem et al. 1998; Young and Bilby 1993). Specifically, Young and Bilby (1993) demonstrated that performing resistance-training exercises with maximal acceleration efforts increased RFD more than when exercises were performed in a slow manner, despite similar loads lifted by the two training groups. As previously mentioned, the ERT group involves movements such as the snatch, hang clean, and push jerk which may contribute directly to explosive performance or strength (Caserotti et al. 2008), since these exercises involve attaining maximum or near maximum rate of force development to accomplish (Stone and O’Bryant 1987). Collectively, this emphasizes the importance of rapid force characteristics in relation to ERT.
Rate of force or torque development is a variable that has been previously assessed for TRT groups with inconsistent results. For example, Aagaard et al. (2002) utilized a heavy-resistance strength training intervention over 14 weeks and reported a significant 15–20% increase in early phase RTD (0–30, 0–50, 0–100 ms). Additionally, Thompson et al. (2015) revealed increases for the KE and KF for early and late phase RTD (0–50, 0–200 ms) following 10 weeks of barbell deadlift training. In contrast, Andersen et al. (2010) implemented training program over 14 weeks and found that early phase RFD remained unchanged even though a significant increase in maximal strength was observed. In contrast, Marshall et al. (2011) reported a decrease for early phase RFD (0–30, 0–50 ms) and RFDpeak in the KE following squat training. While TRT and early RTD has shown dissimilar responses post intervention, studies assessing RFD/RTD following ERT protocols appear to show a reliable increase (Gruber et al. 2007; Oliveira et al. 2013a, b; Young and Bilby 1993). For example, Gruber et al. (2007) reported a 48% increase in RFDpeak following just 4 weeks of explosive resistance training despite the MVC remaining unchanged. Also, Young and Bilby (1993) revealed a 68.7% increase in RFDpeak in the ERT (fast) group compared to just 23.5% in the TRT (slow) group following 7 weeks of squat training. While the present study assessed two different training groups (ERT and TRT), the existing findings suggest that baseline RTD0–50 of the KE was greater for the ERT compared to TRT groups. These findings may indicate that early phase RTD may be a more sensitive measure when distinguishing between two different resistance-trained groups and highlights the importance of rapid torque characteristics.
As aforementioned, this investigation revealed a significant difference for knee extension RTD0–50, in which the ERT group had greater early RTD values compared to the TRT group. While it is plausible that this finding could be attributed to one’s training background (explosive based contractions vs. a traditional controlled movement pattern), previous literature suggests that cross-bridge cycling rates, muscle fiber type, and calcium kinetics (Aagaard et al. 2002; Andersen et al. 2010; Bottinelli et al. 1996; Brody 1976; Larsson and Moss 1993) are directly related with early RTD. Therefore, the present findings could be directly related to these aforementioned characteristics. Additionally, knowing that TRT normally incorporates higher intensities at lower speeds (Channell and Barfield 2008), whereas ERT performs movements at maximum or near maximum rates of force development (Stone and O’Bryant 1987), the baseline early RTD results suggest that ERT exercise prescriptions may provide significant differences in early RTD performance. Nevertheless, another unique finding from the present study was the lack of difference post fatigue between the two training groups; thus, while there was a significant difference between groups at baseline, similar fatigue related responses occurred immediately post-exercise. As previously stated, this may be due to the training status of the groups. Specifically, resistance training causes a transition in myosin heavy chain (MHC) isoforms from MHCIIb to MHCIIa in as early as 4–12 weeks (Kraemer et al. 1995; Rapp and Weicker 1982; Staron et al. 1994). This transition of MHC isoforms corresponds to a shift from type IIB to type IIA fibers and facilitates fiber hypertrophy, which is associated with strength gains and increased oxidative capacity to the muscle, thus improving fatigue resistance of the trained muscles (Abernethy et al. 1994; Kraemer et al. 1996). In addition, we had expected that any adaptation that improves RTD would be accompanied by higher ACC and Vmax. Surprisingly, our investigation did not reveal these variables to be greater as well. While the present study did not assess any recovery time besides immediately post-exercise, future studies may wish to incorporate a variety of training backgrounds (TRT, ERT, sedentary, endurance trained, etc.) assessment of muscle fiber type, EMG, as well as track an acute post-exercise recovery period to see if there are similar or dissimilar recovery patterns between training status, further, incorporating assessments of rapid and maximal strength could add to the literature.
Although the present investigation used an isokinetic dynamometer to assess PT, RTD, Vmax, and ACCmax, we acknowledge that not everyone has the ability to utilize this mode assessment. Future studies should consider comparing these performance measures from different, more practical dynamic exercises. In addition, the present study included only young resistance-trained individuals, thus, the measures may greatly differ compared to older populations that utilize the same training techniques. Lastly, it should be noted that our only significant difference between groups showed a p value of 0.049, if we were to use a Bonferroni correction due to the multiple comparisons this would change the alpha level required to 0.005.
Conclusion
This study was designed to determine if maximal and rapid force characteristics as well as fatigue responses of the knee extensors and flexors could discriminate between TRT and ERT men. The evidence from the present study seems to support the idea that resistance-trained individuals may react similarly when put through a fatiguing bout of dynamic isokinetic exercise. However, it has been shown that the adaptations to training may be specific to the modality of the training. Therefore, it is possible that training specificity of the two groups (i.e., TRT and ERT) led to adaptations that may not completely transfer to isokinetic and isometric performance assessments, since they are different modalities. In addition, the present study indicated a significant difference for the extensors, where RTD0–50 was higher for the ERT group compared to the TRT group. However, the greater RTD0–50 in ERT men was not accompanied by a significantly greater acceleration or maximal velocity. These findings demonstrate that measures of rapid force development may be more sensitive to training-specific adaptations than kinematic variables such as peak acceleration and velocity.
Abbreviations
- ACCmax :
-
Peak acceleration
- ERT:
-
Explosive resistance-trained
- FI%:
-
Fatigue index
- KE:
-
Knee extensors
- KF:
-
Knee flexors
- MVC:
-
Maximal voluntary contraction
- PT:
-
Peak torque
- ROM:
-
Range of motion
- RTD:
-
Rate of torque development
- TRT:
-
Traditional resistance-trained
- V max :
-
Maximal unloaded velocity
References
Aagaard P, Simonsen EB, Andersen JL, Magnusson P, Dyhre-Poulsen P (2002) Increased rate of force development and neural drive of human skeletal muscle following resistance training. J Appl Physiol 93:1318–1326
Abernethy PJ, Jurimae J, Logan PA, Taylor AW, Thayer RE (1994) Acute and chronic response of skeletal muscle to resistance exercise. Sports Med 17:22–38
Alway S, MacDougall J, Sale D, Sutton J, McComas A (1988) Functional and structural adaptations in skeletal muscle of trained athletes. J Appl Physiol 64:1114–1120
Andersen L, Andersen J, Zebis M, Aagaard P (2010) Early and late rate of force development: differential adaptive responses to resistance training? Scand J Med Sci Sports 20
Behm D (1995) Neuromuscular implications and applications of resistance training. J Strength Cond Res 9:264–274
Behm D, Sale D (1993) Intended rather than actual movement velocity determines velocity-specific training response. J Appl Physiol 74:359–368
Bergeron MF et al (2011) Consortium for Health and Military Performance and American College of Sports Medicine consensus paper on extreme conditioning programs in military personnel. Curr Sports Med Rep 10:383–389. https://doi.org/10.1249/JSR.0b013e318237bf8a
Bilodeau M, Erb MD, Nichols JM, Joiner KL, Weeks JB (2001) Fatigue of elbow flexor muscles in younger and older adults. Muscle Nerve 24:98–106
Campos GE et al (2002) Muscular adaptations in response to three different resistance-training regimens: specificity of repetition maximum training zones. Euro J Appl Physiol 88:50–60
Caserotti P, Aagaard P, Buttrup Larsen J, Puggaard L (2008) Explosive heavy-resistance training in old and very old adults: changes in rapid muscle force, strength and power Scand. J Med Sci Sports 18:773–782
Channell BT, Barfield JP (2008) Effect of Olympic and traditional resistance training on vertical jump improvement in high school boys. J Strength Cond Res 22:1522–1527
Conchola EC, Thompson BJ, Smith DB (2013) Effects of neuromuscular fatigue on the electromechanical delay of the leg extensors and flexors in young and old men. Eur J Appl Physiol 113:2391–2399
Conchola EC, Thiele RM, Palmer TB, Smith DB, Thompson BJ (2015) Effects of neuromuscular fatigue on the electromechanical delay of the leg extensors and flexors in young men and women. Muscle Nerve 52:844–51
Corcos DM, Jiang H-Y, Wilding J, Gottlieb GL (2002) Fatigue induced changes in phasic muscle activation patterns for fast elbow flexion movements. Exp Brain Res 142:1–12
Fielding RA, LeBrasseur NK, Cuoco A, Bean J, Mizer K, Singh MAF (2002) High-velocity resistance training increases skeletal muscle peak power in older women. J Am Ger Soc 50:655–662
Gandevia S (1992) Some central and peripheral factors affecting human motoneuronal output in neuromuscular fatigue. Sports Med 13:93–98
Gruber M, Gruber SB, Taube W, Schubert M (2007) Differential effects of ballistic versus sensorimotor training on rate of force development and neural activation in humans. J Strength Cond Res 21:274
Hak PT, Hodzovic E, Hickey B (2013) The nature and prevalence of injury during CrossFit training. J Strength Cond Res. https://doi.org/10.1519/JSC.0000000000000318
Häkkinen K, Keskinen K (1989) Muscle cross-sectional area and voluntary force production characteristics in elite strength-and endurance-trained athletes and sprinters Euro. J Appl Physiol Occup Physiol 59:215–220
Hansen JW (1961) The training effect of repeated isometric muscle contractions. Int Z Angew Physiol 18:474–477
Hansen JW (1963) The training effect of dynamic maximal resistence exercises. Int Z Angew Physiol 19:420–424
Hooper D et al (2013) Effects of resistance training fatigue on joint biomechanics. J Strength Cond Res 27:146–153
Izquierdo M et al (2009) Neuromuscular fatigue after resistance training Inter. J Sports Med 30:614
Jones D, Rutherford O (1987) Human muscle strength training: the effects of three different regimens and the nature of the resultant changes. J Physiol 391:1–11
Jozsi A, Campbell W, Joseph L, Davey S, Evans W (1999) Changes in power with resistance training in older and younger men and women. J Gerontol Ser A Biol Sci Med Sci 54:M591–M596
Judge L, Moreau C, Burke J (2003) Neural adaptations with sport-specific resistance training in highly skilled athletes. J Sports Sci 21:419–427
Kraemer W, Newton R (2000) Training for muscular power. Phys Med Rehabil Clin N Am 11:341–368
Kraemer WJ et al (1995) Compatibility of high-intensity strength and endurance training on hormonal and skeletal muscle adaptations. J Appl Physiol (1985) 78:976–989. https://doi.org/10.1152/jappl.1995.78.3.976
Kraemer WJ, Fleck SJ, Evans WJ (1996) Strength and power training: physiological mechanisms of adaptation. Exerc Sport Sci Rev 24:363–397
Lattier G, Millet GY, Maffiuletti NA, Babault N, Lepers R (2003) Neuromuscular differences between endurance-trained, power-trained, and sedentary subjects. J Strength Cond Res 17:514–521
MacDonald CJ, Lamont HS, Garner JC (2012) A comparison of the effects of 6 weeks of traditional resistance training, plyometric training, and complex training on measures of strength and anthropometrics. J Strength Cond Res 26:422–431. https://doi.org/10.1519/JSC.0b013e318220df79
Mangine G, Ratamess N, Hoffman J, Faigenbaum A, Kang J, Chilakos A (2008) The effects of combined ballistic and heavy resistance training on maximal lower-and upper-body strength in recreationally trained men. J Strength Cond Res 22:132–139
Marshall PW, McEwen M, Robbins DW (2011) Strength and neuromuscular adaptation following one, four, and eight sets of high intensity resistance exercise in trained males Euro. J Appl Physiol 111:3007–3016
Mathiassen SE (1989) Influence of angular velocity and movement frequency on development of fatigue in repeated isokinetic knee extensions. Euro J Appl Physiol Occup Physiol 59:80–88
McCaulley GO, McBride JM, Cormie P, Hudson MB, Nuzzo JL, Quindry JC, Triplett NT (2009) Acute hormonal and neuromuscular responses to hypertrophy, strength and power type resistance exercise. Eur J Appl Physiol 105:695–704
Murray D, Brown L, Zinder S, Noffal G, Bera S, Garrett N (2007) Effects of velocity-specific training on rate of velocity development, peak torque, and performance. J Strength Cond Res 21:870–874
Newton RU, Kraemer WJ, Haekkinen K (1999) Effects of ballistic training on preseason preparation of elite volleyball players. Med Sci Sports Exerc 31:323–330
Newton R, Hakkinen K, Hakkinen A, McCormick M, Volek J, Kraemer W (2002) Mixed-methods resistance training increases power and strength of young and older men. Med Sci Sports Exerc 34:1367–1375
Newton M, Morgan G, Sacco P, Chapman D, Nosaka K (2008) Comparison of responses to strenuous eccentric exercise of the elbow flexors between resistance-trained and untrained men. J Strength Cond Res 22:597–607
Oliveira F, Oliveira A, Rizatto G, Denadai B (2013a) Resistance training for explosive and maximal strength: effects on early and late rate of force development. J Sports Sci Med 12:402
Oliveira F, Rizatto G, Denadai B (2013b) Are early and late rate of force development differently influenced by fast-velocity resistance training? Clin Physiol Funct Imaging 33:282–287
Pääsuke M, Ereline J, Gapeyeva H (1999) Neuromuscular fatigue during repeated exhaustive submaximal static contractions of knee extensor muscles in endurance-trained, power-trained and untrained men. Acta Physiol Scand 166:319–326
Pereira M, Gomes P (2003) Movement velocity in resistance training. Sports Med 33:427–438
Rapp G, Weicker H (1982) Comparative studies on fast muscle myosin light chains after different training programs. Int J Sports Med 3:58–60. https://doi.org/10.1055/s-2008-1026065
Rasch PJ, Morehouse LE (1957) Effect of static and dynamic exercises on muscular strength and hypertrophy. J Appl Physiol 11:29–34. https://doi.org/10.1152/jappl.1957.11.1.29
Schuenke M et al (2012) Early-phase muscular adaptations in response to slow-speed versus traditional resistance-training regimens. Euro J Appl Physiol 112:3585–3595
Smith MM, Sommer AJ, Starkoff BE, Devor ST (2013) Crossfit-based high-intensity power training improves maximal aerobic fitness and body composition. J Strength Cond Res 27:3159–3172. https://doi.org/10.1519/JSC.0b013e318289e59f
Staron RS et al (1994) Skeletal muscle adaptations during early phase of heavy-resistance training in men and women. J Appl Physiol (1985) 76:1247–1255. https://doi.org/10.1152/jappl.1994.76.3.1247
Stone MH, O’Bryant HS (1987) Weight training: a scientific approach. Burgess International Group, New York
Suetta C, Aagaard P, Rosted A, Jakobsen A, Duus B, Kjaer M, Magnusson S (2004) Training-induced changes in muscle CSA, muscle strength, EMG, and rate of force development in elderly subjects after long-term unilateral disuse. J Appl Physiol 97:1954–1961
Thompson BJ, Ryan ED, Sobolewski EJ, Conchola EC, Cramer JT (2013) Age related differences in maximal and rapid torque characteristics of the leg extensors and flexors in young, middle-aged and old men. Exp Gerontol 48:277–282
Thompson BJ, Conchola EC, Palmer TB, Stock MS (2014) Effects of aging on maximal and rapid velocity capacities of the leg extensors. Exp Gerontol 58:128–131
Thompson BJ et al (2015) Barbell deadlift training increases the rate of torque development and vertical jump performance in novices. J Strength Cond Res 29:1–10
Thorstensson A, Karlsson J (1976) Fatiguability and fibre composition of human skeletal muscle. Acta Physiol Scand 98:318–322
Van Cutsem M, Duchateau J, Hainaut K (1998) Changes in single motor unit behaviour contribute to the increase in contraction speed after dynamic training in humans. J Physiol 513:295–305
Wadden KP, Button DC, Kibele A, Behm DG (2012) Neuromuscular fatigue recovery following rapid and slow stretch-shortening cycle movements. Appl Physiol Nutr Metab 37:437–447. https://doi.org/10.1139/h2012-020
Winchester JB, McBride JM, Maher MA, Mikat RP, Allen BK, Kline DE, McGuigan MR (2008) Eight weeks of ballistic exercise improves power independently of changes in strength and muscle fiber type expression. J Strength Cond Res 22:1728–1734
Young WB, Bilby GE (1993) The effect of voluntary effort to influence speed. J Strength Cond Res 7:172–178
Author information
Authors and Affiliations
Contributions
Authors whose names appear on the submission have contributed sufficiently to the scientific work and, therefore, share collective responsibility and accountability for the results.
Corresponding author
Ethics declarations
Conflict of interest
None of the authors has any conflict of interest to disclose.
Additional information
Communicated by William J. Kraemer.
Rights and permissions
About this article

Cite this article
Mackey, C.S., Thiele, R.M., Conchola, E.C. et al. Comparison of fatigue responses and rapid force characteristics between explosive- and traditional-resistance-trained males. Eur J Appl Physiol 118, 1539–1546 (2018). https://doi.org/10.1007/s00421-018-3883-2
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00421-018-3883-2