
Abstract
Background
Children have been shown to have higher lactate (LaTh) and ventilatory (VeTh) thresholds than adults, which might be explained by lower levels of type-II motor-unit (MU) recruitment. However, the electromyographic threshold (EMGTh), regarded as indicating the onset of accelerated type-II MU recruitment, has been investigated only in adults.
Purpose
To compare the relative exercise intensity at which the EMGTh occurs in boys versus men.
Methods
Participants were 21 men (23.4 ± 4.1 years) and 23 boys (11.1 ± 1.1 years), with similar habitual physical activity and peak oxygen consumption (VO2pk) (49.7 ± 5.5 vs. 50.1 ± 7.4 ml kg−1 min−1, respectively). Ramped cycle ergometry was conducted to volitional exhaustion with surface EMG recorded from the right and left vastus lateralis muscles throughout the test (~10 min). The composite right–left EMG root mean square (EMGRMS) was then calculated per pedal revolution. The EMGTh was then determined as the exercise intensity at the point of least residual sum of squares for any two regression line divisions of the EMGRMS plot.
Results
EMGTh was detected in 20/21 of the men (95.2 %) and only in 18/23 of the boys (78.3 %). The boys’ EMGTh was significantly higher than the men’s (86.4 ± 9.6 vs. 79.7 ± 10.0 % of peak power output at exhaustion; p < 0.05). The pattern was similar when EMGTh was expressed as percentage of VO2pk.
Conclusions
The boys’ higher EMGTh suggests delayed and hence lesser utilization of type-II MUs in progressive exercise, compared with men. The boys–men EMGTh differences were of similar magnitude as those shown for LaTh and VeTh, further suggesting a common underlying factor.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Children’s response to exercise is physiologically and functionally different from adults’. For example, compared with adults, children demonstrate lower body-size-normalized maximal isometric strength (Falk et al. 2009) and short-term power (Beneke et al. 2007; Van Praagh and Dore 2002), higher ventilatory threshold (VeTh) (Klentrou et al. 2006) and lactate threshold (LaTh) (Simon et al. 1981; Tanaka and Shindo 1985). In terms of neuromuscular function, children have been shown to have lower motor-unit (MU) activation of the knee extensor muscles (Blimkie 1989; O’Brien et al. 2009, 2010), and a lower mean power frequency during sustained maximal voluntary contraction (MVC) (Halin et al. 2003). According to the size principle (Henneman et al. 1965), the lower level of voluntary recruitment implies that children activate their higher threshold, fast-twitch, type-II MUs to a lesser extent than do adults. The lesser activation of type-II MUs and greater reliance on slow-twitch, type-I MUs may, in turn, explain many performance, metabolic, and neuro-motor child–adult differences beyond just isometric muscle strength (Dotan et al. 2012). Direct supportive evidence for this differential MU activation hypothesis is lacking due to technical or ethical constraints associated with invasive techniques. Hence, new investigative approaches ought to be explored in attempting to further support or refute the hypothesis.
Currently, no technique is available for directly monitoring the proportion of active type-I versus type-II MUs. Furthermore, invasive procedures that might be available for adults (e.g., needle electrodes) are not ethically acceptable for children. A non-invasive technique that is widely accepted as being capable of discerning the onset of type-II MU activation in adults, is the electromyographic threshold (EMGTh) (Candotti et al. 2008; Edwards and Lippold 1956; Hug et al. 2006b, 2003; Lucia et al. 1999; Maestu et al. 2006; Moritani and deVries 1978; Moritani et al. 1984, 1993; Nagata et al. 1981; Petrofsky 1979; Tikkanen et al. 2012). The EMGTh is defined as a non-linear increase, or upward inflection, in the EMG signal during progressive exercise of increasing intensity (Miyashita and Kanehisa 1980; Moritani and deVries 1978). The EMGTh concept is supported by evidence of orderly depletion of glycogen first in type-I and IIA, followed by IIAX (IIAB), and finally IIX (IIB) muscle fibers at increasing workloads (Vollestad and Blom 1985), and by increasing muscle fiber conduction velocity at progressively higher power outputs, suggesting progressive recruitment of large, high-conduction-velocity MUs with increasing muscle force (Farina et al. 2004).
In adults, the EMGTh has been investigated in untrained as well as trained individuals, mostly in cycling and running, but also in upper-extremity exercise modes (Bearden and Moffatt 2001; Candotti et al. 2008; Chwalbinska-Moneta et al. 1994, 1998; Hug et al. 2003, 2004, 2006b, Lucia et al. 1999; Moritani et al. 1993; Nagata et al. 1981; Takaishi et al. 1992; Taylor and Bronks 1994; Tikkanen et al. 2012; Viitasalo et al. 1985), and in a wide range of muscles (Bearden and Moffatt 2001; Chwalbinska-Moneta et al. 1994; 1998; Hug et al. 2003, 2006b; Lucia et al. 1999; Moritani et al. 1984; Nagata et al. 1981; Takaishi et al. 1992). Alongside the EMGTh, a number of studies also determined the VeTh (Nagata et al. 1981; Tikkanen et al. 2012), the LaTh (Candotti et al. 2008; Chwalbinska-Moneta et al. 1998; Moritani et al. 1993; Moritani et al. 1984; Nagata et al. 1981), and the onset of blood lactate accumulation (OBLA) (Tikkanen et al. 2012). These thresholds highly correlated with the EMGTh, although they did not necessarily coincide. This inter-threshold correlation could be expected as increased activation of the more glycolytic type-II MUs, presumed to occur at the EMGTh, implies higher lactate production and accumulation, increased acidosis, and elevated ventilatory drive.
The EMGTh has not been studied in children. Thus, the purpose of this study was to compare the relative exercise intensity at which EMGTh occurs in children versus adults. Given the close relationships between EMGTh and the LaTh and VeTh seen in adults, and the observed higher exercise intensity at which the latter thresholds occur in children, it was hypothesized that the EMGTh would also occur at higher relative exercise intensities in children than in adults. Such findings would suggest a different muscle activation regimen, in which children recruit type-II MUs later and to a lesser extent than do adults.
Methods
Participants
All tests and procedures complied with the Helsinki declaration and were cleared by Brock University’s Research Ethics Board. Twenty-three boys, aged 8–13 years, and 21 men, aged 18–32 years, with similar weekly physical activity history and peak oxygen consumption (VO2pk), volunteered for the study. A summary of participants’ characteristics is provided in Table 1. Written informed consent was provided by all adult participants and by the boys’ parents or guardians prior to enrolment in the study. All boys provided their informed assent to participate. Participants completed questionnaires regarding medical history, physical activity (Godin and Shephard 1985) and sport training history. All participants were physically active and all child participants were involved in some form of organized, competitive sports. Boys were significantly smaller than men but no differences were observed in body composition or maximal oxygen consumption. Sexual maturity in the boys, as determined by self-assessment of secondary sex characteristics [pubic hair (Tanner 1962)], ranged from stages 1 to 4, with eight boys at stage 1, eight at stage 2, five at stage 3, and one at stage 4. One participant’s sexual maturity value was excluded from analysis due to misreport concerns. Years to age of peak height velocity (PHV) for the boys, as estimated using the Mirwald equation (Mirwald et al. 2002), averaged −2.30 ± 0.63 years (n = 18; range −0.88 to −3.59).
Experimental protocol
Participants made two laboratory visits. During the first visit, participants were informed of all tests and procedures to take place over both visits. Anthropometric variables such as height, body mass, and percent body fat (%Fat) were measured. Skinfold thickness was measured in triplicate using Harpenden calipers (British Indicators, Herts, England) and the median value at each site was used. Skinfold thickness (triceps and subscapula) was used to estimate percent body fat using age- and maturity-specific equations (Slaughter et al. 1988). Pubertal stage and physical activity and training history were also determined. Participants were then familiarized with the crank-length-adjustable cycle-ergometer (Excalibur Sport, Lode, Groningen, The Netherlands). Seat and handlebar positioning were determined and recorded for re-use in the subsequent visit. Proper crank length was individually determined at 5-mm increments as a function of inseam length. Participants then proceeded to perform a submaximal and maximal progressive cycling test to determine aerobic capacity (see below).
The second visit was scheduled 3–7 days following the first visit and included a maximal progressive cycling test for the determination of the EMGTh (see below).
Exercise testing
Submaximal VO2 and VO2pk tests (Visit 1): Following a 5-min warm-up period, participants commenced an incremental exercise protocol consisting of 3–5 submaximal stages to determine the VO2-versus-power relationship. Stages were 3.5- and 4-min long for the boys and the men, respectively. Boys started out at 40–60 W and progressed in 15–20 W increments at each subsequent stage. Men started at 80–100 W with 30–40 W increments. These submaximal stages were used to determine the steady-state VO2 in several progressive power output levels.
Following the completion of the submaximal stages, participants recovered for 10–20 min. Participants then performed an incremental exercise protocol to volitional exhaustion for the determination of peak aerobic power (PVO2pk; the mechanical power output corresponding to VO2pk). Workload was increased every minute (10 and 20 W min−1, for the boys and men, respectively) until volitional exhaustion. Pedaling rate was maintained at a minimum of 80 revolutions per minute (rpm).
HR was determined using a HR monitor (Timex Personal Heart Rate Monitor, Timex Group Inc., Toronto, ON, Canada) throughout the test. Expired gas was collected and analyzed using the Moxus metabolic cart (AEI technologies, PA, USA), calibrated prior to each test. VO2pk was determined as the average of the highest VO2 values attained over three consecutive 15-s periods.
EMG Th test (Visit 2): Ryan and Gregor (1992) showed that the mono-articular muscles gluteus maximus, vastus lateralis (VL), vastus medialis, tibialis anterior, and soleus play a relatively invariant role as primary power producers in cycling. Hug et al. (2006b) showed VL to be the most consistent of eight cycling-involved muscles in demonstrating the EMGTh. We, therefore, chose VL as our tested muscle.
Following a 5-min warm-up period, participants performed a ramped cycling test to exhaustion. Starting power output was set at ~35–40 % of PVO2pk and, based on the previously determined PVO2pk, the ramp protocol was set so as to reach volitional exhaustion after ~10 min of exercise. Thus, the mean starting power output for the boys was 51 ± 12 W and power output increased on average 1 W every 6 s. The corresponding values for men were 103 ± 21 W and 1 W every 3 s. Surface EMG, using 10 mm2, bipolar, Ag/Ag surface electrodes (Delsys 2.1, Delsys Inc., Boston, MA, USA), was used to continuously monitor the VL EMG of each leg throughout the test. An area of each thigh at two-thirds the distance from the anterior superior spina iliaca to the superior border of the patella was shaved (if necessary), abraded with skin preparation gel (Nuprep, Weaver and Company, Aurora, CO, USA), and cleaned with rubbing alcohol. Electrodes were placed parallel to the direction of muscle fibers at approximately mid-width of the VL and affixed with proprietary double-sided tape. A reference electrode was placed over the spinous process of the seventh cervical vertebra. Participants were instructed and supervised to maintain a pedaling rate as close to 80 rpm as possible throughout the test and verbal encouragement was provided throughout the test and particularly as the participant approached exhaustion.
EMG data reduction
EMG signals were band-pass filtered (20–450 Hz) using the Bagnoli-4 (Delsys Inc., Boston, MA, USA) bioamplifier and sampled at a rate of 1,000 Hz using a Computer-Based Oscillograph and Data Acquisition System (EMGworks Acquisition, Delsys Inc., Boston, MA, USA).
EMG data were analyzed using a dedicated computer algorithm created in MATLAB (2013 version; MathWorks Inc., Natick, MA, USA). For each pedal stroke, a succinct EMG burst was defined and recorded for each leg and the record was pruned at the beginning and end of each stroke to remove any partial or incomplete bursts, if any. The pruned waveform was then de-trended to offset any baseline deviation (i.e., set baseline value to zero). The root mean square (RMSEMG) was then calculated for each stroke throughout the entire EMG trace. The onset and offset of each RMSEMG burst were identified as the points where RMSEMG rose or fell, respectively, above or below 10 % of the mean RMSEMG value of the entire test record. The mean RMSEMG of each burst (i.e., between the onset and offset) was then extracted for EMGTh determination.
EMGTh Determination
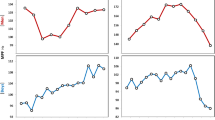
A composite plot, averaging the RMSEMG bursts from both legs, was constructed for each participant. This plot consisted of RMSEMG values (one for each pedal stroke) plotted against the test duration. To reduce the internal fluctuation, a trimmed moving average (30-point window in which the lowest 10 and highest 10 values were trimmed off and the mean of the median 10 points was calculated) was applied to the plot (Fig. 1). In cases where a drop in the RMSEMG at the end of the test was observed in conjunction with a sustained fall below 80 rpm in pedaling cadence, the plot was truncated at the point where cadence began to fall. The EMGTh was then determined by a computer algorithm as the point of least residual sum of squares for any two linear regression line divisions of the data, similar to the approach of Hug et al. (2006a).

A representative RMSEMG trace of an adult man participant. Note the clear rise of the trace above the +3SD confidence interval
Since the algorithm would always determine a point of least residual sum of squares, even when no actual threshold exists, an additional criterion was used to qualify a threshold. Based on previous literature (Hug et al. 2006b), EMGTh was expected to occur at relative power outputs higher than ~80 % P max in adults, and we hypothesized this to be even higher in children. Based on this assumption, a linear regression line was determined for the initial 70 % of the test duration, corresponding to ~80 % of P max (since the EMGTh test commenced at a power output equal to ~35–40 % of PVO2pk). That line was extrapolated to the entire test duration. A 3-standard deviation (SD) confidence interval was applied above the trend line and extended to the end of the trace (Fig. 1). An EMGTh was then confirmed only if the RMSEMG plot rose and stayed above the confidence limit (e.g., Fig. 1), without descending back to within the confidence interval until the end of the test (e.g., Fig. 2). For those participants showing a threshold, the power output at the EMGTh time point was determined. This power output was expressed as a percentage of the peak power output reached during the EMGTh test (%P max) and as a percentage of VO2pk (%VO2pk), based on the data obtained from the first session’s VO2 testing.

A representative RMSEMG trace of a boy participant. No EMGTh is identifiable in this trace
Statistical analysis
All statistical analysis was performed using SPSS v.20 (SPSS Inc., Chicago, IL, USA). The data for all groups are presented as means ± 1 SD. All data were normally distributed. Group differences in physical characteristics and %P max and %VO2pk at EMGTh were assessed using a two-tailed, homoscedastic Student’s t test. Additionally, group differences between ‘Responders’ and ‘Non-Responders’ (see “Results” section) were examined using a two-tailed Student’s t test. Despite the small sample size, assumptions for normal distribution and homogeneity of variance were met. Pearson’s correlations were computed for the boys, men, and the combined groups, between the EMGTh (both as %VO2pk and %Pmax) and VO2pk kg BW−1, PVO2pk kg BW−1, Pmax kg BW−1, leisure time physical activity, and training hours per week, respectively. The acceptable level of significance for all tests was set at p < 0.05.
Results
Physical activity scores and training histories were similar in the two groups (Table 1). Importantly, peak heart rate was, on average, 4 bpm higher in the boys whose VO2pk was also marginally higher. While none of these differences reached statistical significance they do strongly suggest that the boys had attained a comparable level of exertion to that of the men.
An EMGTh was identified in 20 out of the 21 men (95.2 %) and in 18 of 23 boys (78.3 %) (\(\chi^{2}_{(1, \, n = 44)}\) = 2.69, p = 0.10). In the participants for whom a threshold was identified, group differences showed 6.6 % higher (later) EMGTh %P max in the boys compared with the men (t (36) = −2.08, p = 0.045) (86.4 ± 9.6 % vs. 79.7 ± 10.0 %, respectively; Fig. 3). There was also a trend for a higher EMGTh %VO2pk in the boys versus men (t (35) = −1.62, p = 0.12) (101.2 ± 11.4 vs. 94.8 ± 12.2 %, respectively; Fig. 3) (only 17 boys included. VO2 data could not be obtained for one boy).

Group differences in relative exercise intensity at the EMGTh between boys and men in whom EMGTh was identified (‘Responders’). Values are Mean ± 1SD. * boys significantly higher than men, p < 0.05
Correlational analyses revealed that for all participants, EMGTh (%VO2pk) was not correlated with body-mass-normalized VO2pk (VO2pk kg−1) (r = −0.18, p > 0.05). However, when examined separately for each group, a significant negative correlation was observed in the boys (r = −0.51, p = 0.015), but not in the men (r = 0.18, p > 0.05). Also, EMGTh (%VO2pk) similarly correlated with the power output at VO2pk kg−1 (PVO2pk kg−1) in the boys (r = −0.52, p = 0.03) (Fig. 4), but not in the men. No significant correlations were found between EMGTh %P max or EMGTh %VO2pk and training history, or leisure time physical activity.

Correlation between peak aerobic power (PVO2pk) and EMGTh as %VO2pk for the boys who showed EMGTh (n = 17; VO2 data unavailable for one boy)
For further analysis, the boys who demonstrated EMGTh (‘Responders’, n = 18) were compared to those who did not (‘Non-responders’, n = 5) (Table 2). The ‘Responders’ had significantly longer mean test duration than the ‘Non-responders’ (p = 0.016) as well as significantly greater Pmax/PVO2pk ratio (p = 0.027). There were no statistically significant differences in any other variables. In the men’s group, only one participant did not show an EMGTh. He had exceptionally high VO2pk kg−1 and PVO2pk kg−1 values that were ~2.5 SDs above the group’s mean.
Discussion
The main findings of this study are that (a) EMGTh was identified in only 78.3 % (18/23) of the boys, while the corresponding value in the men was 95.2 % (20/21); and (b) Among those in whom EMGTh was detected, the boys’ EMGTh occurred at significantly higher relative workloads compared with the men. As the EMGTh is widely believed to reflect the onset of accelerated increase in higher-threshold, type-II MU recruitment during progressive exercise (Edwards and Lippold 1956; Hug et al. 2003, 2006b; Lucia et al. 1999; Maestu et al. 2006; Moritani and deVries 1978; Moritani et al. 1993, 1984; Nagata et al. 1981; Petrofsky 1979; Tikkanen et al. 2012), these findings suggest that boys activate their type-II MUs later and for a shorter duration than do men.
The results for our men’s group are in close agreement with previous findings (Hug et al. 2006b; Lucia et al. 1999; Takaishi et al. 1992). However, to our knowledge, this is the first study to investigate the EMGTh in children. Compared with the men, our boys’ higher relative EMGTh is in line with our hypothesis, based on known boys–men differences in LaTh and VeTh. Our observed differences of 6.6 %P max and 6.4 %VO2pk are similar to previously reported boys–men %VO2pk differences in VeTh [7.2 %, (Klentrou et al. 2006); 8.9 %, (Anderson and Mahon 2007)] and LaTh [9.1 %, (Anderson and Mahon 2007)] during cycling exercise.
A question arising from our findings is why EMGTh could not be detected in the five ‘Non-responder’ boys? At exhaustion, the force applied by the participating muscles is considerably lower than MVC for the given pedaling cadence (Greig et al. 1985; Sargeant et al. 1981). In view of the fact that 50 % of ‘Responders’ EMGTh occurred very close to exhaustion (>92 % Pmax), it is conceivable that the five ‘Non-Responders’ terminated their tests before attaining the force level at which EMGTh would have taken place. That is, had they been able to continue exercising longer, their EMGTh would have occurred at their observed power at exhaustion or above it. This suggestion is supported by our finding that, in the ‘Responders’, body-mass-normalized peak aerobic power (PVO2pk/kg) negatively correlated with EMGTh %VO2pk (r = −0.52, p = 0.03; Fig. 4). That is, in boys with lower maximal power output, EMGTh was attained closer to maximal aerobic power, compared with more powerful boys. This, in turn, suggests that ‘Non-responders’ may not have been able to produce the power and thus the contractile force required to manifest the EMGTh. Further support for the suggestion is provided by Vollestad and Blom (1985), Vollestad et al. (1984) who found that activation of type IIAX (IIAB) and IIX (IIB) MUs in adult men, during progressive cycling, took place only at 91 % VO2pk and beyond. As shown by Greig et al. (Greig et al. 1985), this power output level would require only ~50 % of the MVC pedal force at that cadence.
Accepting the notion that EMGTh could indeed occur at or above the peak force level attained in progressive exercise to exhaustion, means that EMGTh would only be detected in those individuals able to exceed the critical force by the end of the progressive exercise test. It is conceivable then that ‘Non-Responders’ ended their tests prior to attaining the sufficiently high power output necessary to attain that critical force and elicit EMGTh. Men attain higher relative force and power outputs due to their higher anaerobic capacity compared with the boys (Beneke et al. 2007; Van Praagh and Dore 2002). Thus, most men produce the muscle force necessary for EMGTh manifestation. For this reason, there was little or no correlation between EMGTh and VO2pk or PVO2pk in the men, unlike in the boys. Support for this notion comes from the fact that compared with ‘Non-Responders’, ‘Responders’ had significantly longer test durations and higher Pmax/PVO2pk ratio (Table 2). That is, ‘Responders’ were able to employ more anaerobic power after reaching their respective PVO2pk and thus extend their test durations. Presumably, this was facilitated by the ‘Responders’ greater relative capacity to recruit type-II MUs.
The boys–men difference of 6.6 % in EMGTh %P max (boys = 86.4 %, men = 79.7 %; p = 0.045) is likely an underestimate of the true group difference, since it reflects only the ‘Responders’. Assigning just 100 % as the EMGTh %P max values for the six ‘Non-Responders’ (5 boys, 1 man), the overall boys–men EMGTh difference increased by 30 %, to 8.6 % (boys = 89.3, men = 80.7 %; p < 0.01). Accepting the possibility that EMGTh could occur at >100 % Pmax intensity, the true boys–men difference could well be even larger.
To illuminate possible underlying distinguishing factors, the ‘Responder’ and ‘Non-responder’ boys were compared (Table 2). On average, ‘Responders’ lasted 18 % longer in their respective EMGTh tests than did ‘Non-responders’. Accordingly, their final power output (P max) was 15.5 % higher, although this difference did not reach statistical significance. Since initial loading and the ramping protocol of the EMGTh test were based on peak aerobic power (PVO2pk), >95 % of participants attained a P max that was greater than PVO2pk. However, while the ‘Responders’ P max was 17 % higher, ‘Non-responders’ P max was only 5 % higher than PVO2pk (p < 0.03). Again, this suggests that ‘Non-responders’ were less capable of recruiting the higher-threshold type-II MUs and consequently generating the extra anaerobic power necessary to exceed PVO2pk. The ‘Non-responders’ tended to be younger, lighter, and shorter, have lower lean body mass, be less physically active, and somatically less mature (longer time before PHV). Although none of these differences was statistically significant, the general picture is one of lower maturity level, which may be regarded as consistent with them possessing higher or no detectable EMGTh.
The only man in whom EMGTh could not be detected, had the highest VO2pk (63.6 ml kg−1 min−1) and PVO2pk (5.14 W kg−1), both of which were ~2.5 SD above the mean for the group. Since in comparable studies (e.g., Hug et al. 2003, 2006a, b) the EMGTh was detected in 100 % of participants, we suggest that the absence of EMGTh in this individual was not due to error and that his high aerobic capacity could have been due to particularly high type-I muscle fiber composition. Thus, like the boy ‘Non-Responders’, he too might not have been able to sufficiently engage type-II MUs by the time he reached exhaustion.
Given the previously demonstrated relationships between EMGTh and both LaTh and VeTh in men (Candotti et al. 2008; Chwalbinska-Moneta et al. 1998; Moritani et al. 1993; Moritani et al. 1984; Nagata et al. 1981; Tikkanen et al. 2012), similar relationships presumably exist in boys, as well. In both men and boys, however, there is a persistent difference between exercise intensity at LaTh or VeTh versus that of EMGTh. For example, in men, LaTh and VeTh have been typically found between ~50 and 60 %VO2pk (Anderson and Mahon 2007; Klentrou et al. 2006; Simon et al. 1986), while EMGTh has been identified at ~90 %VO2pk (Lucia et al. 1999; Takaishi et al. 1992) and at 94.8 % VO2pk in the present study. The corresponding values for boys are ~60–70 % VO2pk for LaTh and VeTh (Anderson and Mahon 2007; Klentrou et al. 2006) and 101.2 % VO2pk for EMGTh (in the present study). While boys’ values are generally ~10 % higher than the men’s, the differences between the EMGTh and the LaTh or VeTh thresholds are rather similar: ~45 % in both groups. A notable exception to this pattern is Candotti et al.’s finding of nearly identical EMGTh and LaTh values in adult recreational cyclists (Candotti et al. 2008). A partial explanation for this discrepancy is the LaTh protocol used by Candotti et al. (continuous, 3-min stages), which overestimates LaTh due to the delay in the lactate response to the fast changing workloads. Furthermore, the testing protocol was exhaustion limited at ~200 W, likely due to lactate accumulation and well before VO2pk or PVO2pk could be reached. Thus, the mean reported EMGTh (134 W) occurred at no more than ~67 % of the end-of-protocol power and in reality likely constituted VO2pk percentage considerably lower than that. Since Hug et al. detected two rather than a single EMGTh in professional cyclists (at 52 and 86 % VO2pk) (Hug et al. 2003), it is also conceivable that the EMGTh reported by Candotti et al. (Candotti et al. 2008) is the first (lower) rather than the second of the two thresholds.
The typical magnitude of the EMGTh − LaTh difference appears to suggest that the EMGTh phenomenon is independent of the factors governing LaTh and VeTh, but the apparent consistency of this difference in men and boys seems to suggest otherwise. Several factors should be considered in explaining the apparent EMGTh − LaTh/VeTh discrepancy: (a) While LaTh and VeTh are systemic, whole-body phenomena, the EMGTh is confined to a single muscle. (b) The metabolic acidosis and increased lactate production that underlie both LaTh and VeTh take place as a function of increasing exercise intensity in the active MUs and could occur regardless of whether or not higher-threshold MUs are involved. The EMGTh, on the other hand, is presumably dependent on type-II MU involvement and could thus occur considerably after LaTh or VeTh have already taken place. (c) Based on evidence such as Vollestad and Blom’s (1985) glycogen depletion findings, LaTh and VeTh could be related to increased type-IIA MU recruitment, which occurs at low or moderate intensities. The EMGTh, on the other hand, might be associated with recruitment of the fast-twitch MU types IIAX (IIAB) and IIx (IIB), which typically occur in the later part of exhaustive exercise.
It may be of interest to draw an analogy between children and elite adult endurance athletes. Compared with untrained men, boys are characterized by markedly higher thresholds of all three types (LaTh, VeTh, EMGTh). The same has been shown to be true for endurance athletes, in whom the higher thresholds are attained by extensive training at moderate intensities that, according to the size principle (Henneman et al. 1965) and glycogen depletion data (Vollestad and Blom 1985; Vollestad et al. 1984), predominantly recruit type-I and possibly the lower portion of the type-IIA MU pool. Also, such athletes often possess higher type-I MU composition to begin with (Costill et al. 1976). Consequently, adult endurance athletes utilize type-I MUs to a greater extent than do non-athletes. The present data indicate that children (boys) are characterized by lower utilization of type-II MUs and, thus, greater reliance on type-I MUs utilization as is the case in endurance athletes. The difference is that in children this is not due to training or talent, but rather to what we suggest is their lower utilization of type-II MUs.
Conclusions
Our findings suggest that the fundamental nature of MU recruitment in progressive exercise [Henneman’s size principle; (Henneman et al. 1965)] is qualitatively similar in boys and men. There is, however, a significant quantitative difference in that the boys’ EMGTh occurred at higher exercise intensities compared with the men’s. This age-related difference suggests later recruitment of higher threshold MUs in boys and is in line with previous findings of higher relative LaTh and VeTh in boys compared with men. These findings support the child–adult differential MU activation hypothesis. We suggest that this support is particularly insightful since the EMGTh is currently the best non-invasive ‘window’ into the type of differential MU activation at the core of the hypothesis.
Abbreviations
- EMG:
-
Electromyography
- EMGTh :
-
Electromyographic threshold
- HR:
-
Heart rate
- LaTh :
-
Lactate threshold
- MU/MUs:
-
Motor unit/motor units
- MVC:
-
Maximal voluntary contraction
- OBLA:
-
Onset of blood lactate accumulation
- PHV:
-
Peak height velocity
- Pmax:
-
Maximal power attained at end of the EMGTh test
- PVO2pk:
-
Peak aerobic power (mechanical power output corresponding to VO2pk)
- RER:
-
Respiratory exchange ratio
- RMS:
-
Root mean square
- SD:
-
Standard deviation
- VeTh :
-
Ventilatory/gas-exchange threshold
- VO2 :
-
Oxygen consumption
- VO2pk:
-
Peak oxygen consumption
References
Anderson CS, Mahon AD (2007) The relationship between ventilatory and lactate thresholds in boys and men. Res Sports Med 15:189–200
Bearden SE, Moffatt RJ (2001) Leg electromyography and the VO2-power relationship during bicycle ergometry. Med Sci Sports Exerc 33:1241–1245
Beneke R, Hutler M, Leithauser RM (2007) Anaerobic performance and metabolism in boys and male adolescents. Eur J Appl Physiol 101:671–677
Blimkie CJ (1989) Age- and sex-associated variation in strength during childhood: anthropometric, morphologic, neurologic, biomechanical, endocrinologic, genetic, and physical activity correlates. In: Gisolfi CV (ed) Perspectives in exercise science and sports medicine, vol 2., Youth, exercise and sportsBenchmark Press, Indianapolis, pp 99–163
Candotti CT, Loss JF, Melo Mde O, La Torre M, Pasini M, Dutra LA, de Oliveira JL, de Oliveira LP (2008) Comparing the lactate and EMG thresholds of recreational cyclists during incremental pedaling exercise. Can J Physiol Pharmacol 86:272–278
Chwalbinska-Moneta J, Hanninen O, Penttila I (1994) Relationships between EMG and blood lactate accumulation during incremental exercise in endurance- and speed-trained athletes. Clin J Sports Med 4:31–38
Chwalbinska-Moneta J, Kaciuba-Uscilko H, Krysztofiak H, Ziemba A, Krzeminski K, Kruk B, Nazar K (1998) Relationship between EMG blood lactate, and plasma catecholamine thresholds during graded exercise in men. J Physiol Pharmacol 49:433–441
Costill DL, Fink WJ, Pollock ML (1976) Muscle fiber composition and enzyme activities of elite distance runners. Med Sci Sports 8:96–100
Dotan R, Mitchell C, Cohen R, Klentrou P, Gabriel D, Falk B (2012) Child-adult differences in muscle activation—a review. Pediatr Exerc Sci 24:2–21
Edwards RG, Lippold OC (1956) The relation between force and integrated electrical activity in fatigued muscle. J Physiol 132:677–681
Falk B, Usselman C, Dotan R, Brunton L, Klentrou P, Shaw J, Gabriel D (2009) Child-adult differences in muscle strength and activation pattern during isometric elbow flexion and extension. Appl Physiol Nutr Metab 34:609–615
Farina D, Macaluso A, Ferguson RA, De Vito G (2004) Effect of power, pedal rate, and force on average muscle fiber conduction velocity during cycling. J Appl Physiol 97:2035–2041
Godin G, Shephard RJ (1985) A simple method to assess exercise behavior in the community. Can J Appl Sport Sci 10:141–146
Greig C, Sargeant AJ, Vollestad NK (1985) Muscle force and fibre recruitment during dynamic exercise in man. J Physiol Lond 371:176P
Halin R, Germain P, Bercier S, Kapitaniak B, Buttelli O (2003) Neuromuscular response of young boys versus men during sustained maximal contraction. Med Sci Sports Exerc 35:1042–1048
Henneman E, Somjen G, Carpenter DO (1965) Functional significance of cell size in spinal motoneurons. J Neurophysiol 28:560–580
Hug F, Laplaud D, Savin B, Grelot L (2003) Occurrence of electromyographic and ventilatory thresholds in professional road cyclists. Eur J Appl Physiol 90:643–646
Hug F, Decherchi P, Marqueste T, Jammes Y (2004) EMG versus oxygen uptake during cycling exercise in trained and untrained subjects. J Electromyogr Kinesiol 14:187–195
Hug F, Laplaud D, Lucia A, Grelot L (2006a) A comparison of visual and mathematical detection of the electromyographic threshold during incremental pedaling exercise: a pilot study. J Strength Condition Res/Natl Strength Condition Assoc 20:704–708
Hug F, Laplaud D, Lucia A, Grelot L (2006b) EMG threshold determination in eight lower limb muscles during cycling exercise: a pilot study. Int J Sports Med 27:456–462
Klentrou N, Nishio M-L, Plyley M (2006) Ventilatory breakpoints in boys and men. Pediatr Exerc Sci 18:216–225
Lucia A, Sanchez O, Carvajal A, Chicharro JL (1999) Analysis of the aerobic-anaerobic transition in elite cyclists during incremental exercise with the use of electromyography. Br J Sports Med 33:178–185
Maestu J, Cicchella A, Purge P, Ruosi S, Jurimae J, Jurimae T (2006) Electromyographic and neuromuscular fatigue thresholds as concepts of fatigue. J Strength Condition Res Natl Strength Condition Assoc 20:824–828
Mirwald RL, Baxter-Jones AD, Bailey DA, Beunen GP (2002) An assessment of maturity from anthropometric measurements. Med Sci Sports Exerc 34:689–694
Miyashita M, Kanehisa H (1980) Correlation between efficiency in cycling and maximal power of human extensor muscles. J Sports Med Phys Fitness 20:365–370
Moritani T, deVries HA (1978) Reexamination of the relationship between the surface integrated electromyogram (IEMG) and force of isometric contraction. Am J Phys Med 57:263–277
Moritani T, Tanaka H, Yoshida T, Ishii C, Yoshida T, Shindo M (1984) Relationship between myoelectric signals and blood lactate during incremental forearm exercise. Am J Phys Med 63:122–132
Moritani T, Takaishi T, Matsumoto T (1993) Determination of maximal power output at neuromuscular fatigue threshold. J Appl Physiol 74:1729–1734
Nagata A, Muro M, Moritani T, Yoshida T (1981) Anaerobic threshold determination by blood lactate and myoelectric signals. Jpn J Physiol 31:585–597
O’Brien TD, Reeves ND, Baltzopoulos V, Jones DA, Maganaris CN (2009) The effects of agonist and antagonist muscle activation on the knee extension moment-angle relationship in adults and children. Eur J Appl Physiol 106:849–856
O’Brien TD, Reeves ND, Baltzopoulos V, Jones DA, Maganaris CN (2010) In vivo measurements of muscle specific tension in adults and children. Exp Physiol 95:202–210
Petrofsky JS (1979) Frequency and amplitude analysis of the EMG during exercise on the bicycle ergometer. Eur J Appl Physiol Occup Physiol 41:1–15
Ryan MM, Gregor RJ (1992) EMG profiles of lower extremity muscles during cycling at constant workload and cadence. J Electromyogr Kinesiol 2:69–80
Sargeant AJ, Hoinville E, Young A (1981) Maximum leg force and power output during short-term dynamic exercise. J Appl Physiol Respir Environ Exerc Physiol 51:1175–1182
Simon J, Young JL, Blood DK, Segal KR, Case RB, Gutin B (1986) 1986 Plasma lactate and ventilation thresholds in trained and untrained cyclists. J Appl Physiol 60:777–781
Simon G, Berg A, Simon-Alt A, Keul J (1981) Determination of the anaerobic threshold depending on age and performance potential. Dtsch Z Sportsmed 32:7–14
Slaughter MH, Lohman TG, Boileau BA (1988) Skinfold equations for estimation of body fatness in children and youth. Hum Biol 60:709–723
Takaishi T, Ono T, Yasuda Y (1992) Relationship between muscle fatigue and oxygen uptake during cycle ergometer exercise with different ramp slope increments. Eur J Appl Physiol Occup Physiol 65:335–339
Tanaka H, Shindo M (1985) Running velocity at blood lactate threshold of boys aged 6-15 years compared with untrained and trained young males. Int J Sports Med 6:90–94
Tanner JM (1962) Growth at adolescence. Blackwell Scientific Publications, Oxford
Taylor AD, Bronks R (1994) Electromyographic correlates of the transition from aerobic to anaerobic metabolism in treadmill running. Eur J Appl Physiol Occup Physiol 69:508–515
Tikkanen O, Hu M, Vilavuo T, Tolvanen P, Cheng S, Finni T (2012) Ventilatory threshold during incremental running can be estimated using EMG shorts. Physiol Meas 33:603–614
Van Praagh E, Dore E (2002) Short-term muscle power during growth and maturation. Sports Med 32:701–728
Viitasalo JT, Luhtanen P, Rahkila P, Rusko H (1985) Electromyographic activity related to aerobic and anaerobic threshold in ergometer bicycling. Acta Physiol Scand 124:287–293
Vollestad NK, Blom PC (1985) Effect of varying exercise intensity on glycogen depletion in human muscle fibres. Acta Physiol Scand 125:395–405
Vollestad NK, Vaage O, Hermansen L (1984) Muscle glycogen depletion patterns in type I and subgroups of type II fibres during prolonged severe exercise in man. Acta Physiol Scand 122:433–441
Acknowledgments
The authors wish to thank all participants for the hard work and dedication they invested in this study. We are also indebted to the boys’ parents or guardians for consenting, bringing the boys, and making it all possible. Special gratitude and appreciation is reserved for Mr. James Desjardins for developing the necessary software in a most proficient manner and with the utmost patience for the whims and wishes of the researchers. Funding: The study was funded by the Canadian Institutes of Health Research, Grant No199944.
Conflict of interest
The authors have no competing interests to declare.
Author information
Authors and Affiliations
Corresponding author
Additional information
Communicated by Toshio Moritani.
Rights and permissions
About this article

Cite this article
Pitt, B., Dotan, R., Millar, J. et al. The electromyographic threshold in boys and men. Eur J Appl Physiol 115, 1273–1281 (2015). https://doi.org/10.1007/s00421-015-3100-5
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00421-015-3100-5